
紫花苜蓿复合体 (M edicago sativa comp lex)叶片形态特征及数量分类研究
2010-12-31 13:50:12崔大方羊海军赵业彬李庆艳陈考科
植物资源与环境学报 2010年3期
崔大方,羊海军,赵业彬,李庆艳,陈考科
(华南农业大学 a.林学院;b.公共基础课实验教学中心;c.生命科学学院,广东 广州 510642)
紫花苜蓿复合体 (Medicagosativacomp lex)是由紫花苜蓿 (M.sativaL.)和与之容易杂交并能产生可育后代的种类共同构成的复合群体,包括紫花苜蓿、黄花苜蓿 (M.falcataL.)以及分布于高加索、中亚细亚、西伯利亚和中国新疆一带的天然杂交类群,如天山苜蓿 (M.tianschanicaVassilcz.)、西锡金苜蓿 (M. schischkiniiSum nev.)、伊犁苜 蓿(M.subdicyclaVassilcz.)、密序苜蓿 (M.agropyretorumVassilcz.)、大花苜蓿 (M.trautvetteriiSum nev.)和座垫苜蓿 (M. rivularisV assilcz.)等[1-2],大体包括 3类表型:1)开紫色花,荚果螺旋盘卷;2)开黄色花,荚果镰状弯曲或稍直;3)介于前两者之间,花色多样,荚果镰状弯曲至螺旋盘卷。其中第 3类植物在形态上有许多重叠,彼此的界限极为模糊,分类学争议较大,成为植物分类学研究的重点。1982年,Sm all等采用 2个元素特征 (Ca和 S)及 12个形态特征对M.sativa-falcata连续体进行了数量分析[3];1984年,Sm all等根据花色、生长习性和荚果盘卷圈数展示了紫花苜蓿、黄花苜蓿和多变苜蓿 (M.variaM artyn)三者之间的连续性变异特点[4];Quiros等利用蛋白标记,将可能与紫花苜蓿天然杂交的M.falcata、M.cretaceaM.B ieb.、M. varia、M.tunetataM urb.和M.glutinosaM.B ieb.合并称为紫花苜蓿复合体[5];1989年,Sm all等总结了苜蓿属(MedicagoL.)的分类进展,支持将多变苜蓿和黄花苜蓿划分为紫花苜蓿的 2个亚种,即M.sativassp.varia(M artyn)A rcang.及M.sativassp.falcata(L.)A rcang.[6];何咏松等则认为紫花苜蓿和黄花苜蓿同属 1个种的 2个亚种或生物型或生态型[7];崔大方等[8]和耿华珠[9]通过叶、花、果、花粉形态以及染色体核型分析,将黄花苜蓿与紫花苜蓿的天然杂交类群归并为多变苜蓿,并在多变苜蓿内新组合了若干种下分类群;李拥军等采用种子贮藏蛋白分析技术和RAPD分子标记技术,支持将紫花苜蓿复合体划分为3个亚种,即M.sativassp.sativa、M.sativassp.varia和M.sativassp.falcata,并发现前 2个亚种间的遗传差异相对较小,而二者与第 3个亚种间的遗传差异相对较大[10-11];孙毅等根据核糖体 DNA的 ITS序列分析结果将黄花苜蓿与紫花苜蓿并入 1个种[12]。
综上所述,由于长期以来在不同地理条件下杂交和分离,紫花苜蓿与黄花苜蓿的基因重新组合,出现了许多天然杂交类群,子代和亲本类型多样,这些类群的分类归属问题一直未能得到科学合理的解决。为此,在前期工作的基础上,作者对紫花苜蓿复合体的叶部表征形态和内部结构进行了全面的观察,并运用比较形态学、比较解剖学和数量分类学等研究方法对紫花苜蓿复合体进行比较分析,为解决紫花苜蓿复合体中各分类群的分类归属问题提供研究依据。
1 材料和方法
1.1 材料
供试紫花苜蓿复合体分类群的具体信息见表 1,凭证标本保存于新疆师范大学植物标本室(XJNU)。
1.2 方法
1.2.1 叶表皮显微特征的观察方法 选取标本上部完好的成熟叶片,在小叶中部中脉附近剪取 0.5 cm×0.5 cm的小片,浸泡在V(体积分数 30%过氧化氢)∶V(乙酸)=1∶1的混合溶液中;60℃温箱中静置 3~5h后,用蒸馏水冲洗干净;剥取叶表皮,用体积分数10%过氧化氢溶液漂白 5~10m in,再用质量体积分数 1%番红溶液染色 20~30 m in,然后用体积分数1%的甘油封片。在 DMLB型显微镜 (德国莱卡公司生产)下观察并照相。气孔密度为 15个观察视野的平均值。

表 1 供试紫花苜蓿复合体各分类群的凭证标本信息Tab le 1 In form a tion of voucher spec im en s of tested taxa in M ed icago sa tiva com p lex
1.2.2 叶表皮亚显微特征的观察方法 选取标本上部完好的成熟叶片,经体积分数 10%过氧化氢溶液漂白 5~10m in,用纯净水洗净,再用体积分数 30%~100%的乙醇进行梯度脱水,用双面胶将叶片固定在样品台上镀膜喷金后,用 XL-30-ESEM型环境扫描电镜(荷兰菲利普公司生产)观察并照相。
1.2.3 叶片组织结构的观察方法 采用石蜡切片法观察叶片的解剖结构特征。从叶片中部至边缘切取面积 0.5 cm×0.5 cm的小片(包含中脉),用 FAA溶液固定后,用体积分数 50%~100%的乙醇进行梯度脱水,然后用二甲苯透明处理,并用石蜡包埋后切片,切片厚度 8~10μm,番红 -固绿对染,中性树胶封片。在DMLB型显微镜下观察、测量并照相。其中,叶片、栅栏组织和海绵组织厚度各测量 30个数值,计算平均值及标准差以及组织疏松度和紧缩度。疏松度 =(海绵组织厚度/叶片厚度)×100%;紧缩度 = (栅栏组织厚度/叶片厚度)×100%。
1.2.4 分类群的表征分析方法 基于表征形态数据分别计算相关系数和平均欧氏距离,并采用UPGMA法进行聚类分析,然后选择最优者应用结合线法进行类群划分[13];采用主成分分析法探讨紫花苜蓿复合体叶部特征的分类学价值。
用于聚类分析的叶片形态表征性状包括 6个数量性状及 9个定性性状。数量性状为上表皮气孔密度、下表皮气孔密度、叶片厚度、栅栏组织层数、组织疏松度、组织紧缩度。定性性状及其赋值为:上表皮细胞形状(不规则形 0,不规则形或多边形 1);上表皮细胞长短轴分化程度 (多数明显 0,明显或不明显 1,少数明显 2);上表皮细胞垂周壁式样(深波状 0,波状至浅波状 1,平直或弓形 2,其他 3);下表皮细胞形状(不规则形 0,不规则形或多边形 1);下表皮细胞长短轴分化程度(多数明显 0,明显或不明显 1,少数明显2);下表皮细胞垂周壁式样 (深波状 0,波状至浅波状1,平直或弓形 2);表皮细胞切面形状(近长方形 0,近长方形或近圆形 1,近圆形 2);栅栏组织细胞排列状况(紧密 0,较紧密 1,较疏松 2,疏松 3);海绵组织细胞排列状况(疏松 0,较疏松 1,较紧密 2,紧密 3)。
1.3 数据处理
数据处理全部使用生物统计“NTSYS-PC”软件包完成。对所采集的数据进行聚类分析,将原来的性状作为运算单位,计算相似性系数并组成相似性系数矩阵,所得矩阵用NTSYS-PC version 2.1e进行处理。
聚类分析程序为:将原始矩阵用 Standardization子程序进行标准化,然后用 Sim ilarity子程序中的Interval data求出欧氏距离矩阵,再用 C lustering程序中的 Shani子程序以UPGMA方法进行聚类。
主成分分析程序为:将原始矩阵用 Standardization子程序进行标准化,然后用 Sim ilarity子程序中的Interval data求出各变量的相关系数,再用 O rdination中的 Eigen子程序算出特征值和特征向量等。
2 结果和分析
2.1 扫描电镜下紫花苜蓿复合体各分类群的叶表皮亚显微特征
紫花苜蓿复合体 6个分类群叶表皮亚显微特征的观察结果见图版Ⅰ-1~6。紫花苜蓿、西锡金苜蓿、座垫苜蓿和天山苜蓿的下表皮细胞均为不规则形(图版Ⅰ-1,3,4,6);黄花苜蓿和大花苜蓿的下表皮细胞多数为不规则形,少数呈多边形(图版Ⅰ-2,5)。根据波状起伏程度,不规则形细胞的垂周壁式样分为深波状、波状和浅波状;多边形细胞的垂周壁式样有平直和弓形。根据细胞表面隆起程度,可分为 3类: 1)紫花苜蓿、西锡金苜蓿、天山苜蓿的细胞表面向上隆起呈山丘状,后者稍不明显;2)黄花苜蓿和座垫苜蓿的细胞表面平整或近平整;3)大花苜蓿的细胞表面近平整或向下凹陷。各分类群的下表皮都被有稀疏表皮毛和气孔器。表皮毛均为单细胞非腺毛,基部膨大,先端锐尖,毛上有微小乳突。所有分类群的气孔器均由气孔和保卫细胞组成,并被 3~5个表皮细胞所包围,属不规则型气孔器,并具有蜡质气孔盖。
2.2 光学显微镜下紫花苜蓿复合体各分类群的叶片表皮形态特征
在光学显微镜下,紫花苜蓿复合体各分类群叶片上、下表皮的细胞形状、长短轴分化程度、垂周壁式样、气孔器类型及气孔密度的观察和统计结果见图版Ⅰ-7~12和表 2。光学显微镜下这 6个分类群叶表皮的形态特征(图版Ⅰ-7~12)与扫描电镜观察结果基本相似。除黄花苜蓿的上、下表皮细胞有少量呈多边形、多数呈不规则形外,其余分类群叶表皮细胞均为不规则形;大部分分类群的叶表皮细胞长短轴明显;垂周壁呈波状,但有深浅之分,其中黄花苜蓿上、下表皮细胞的垂周壁尽管也为波状至浅波状,但也有垂周壁呈平直或弓形,与其他分类群有所不同;6个分类群叶表皮气孔器均属不规则型,上、下表皮均有气孔,且上表皮气孔密度大于下表皮 (天山苜蓿除外),其中黄花苜蓿叶片上、下表皮的气孔密度均最大,分别为 237和 188mm-2。
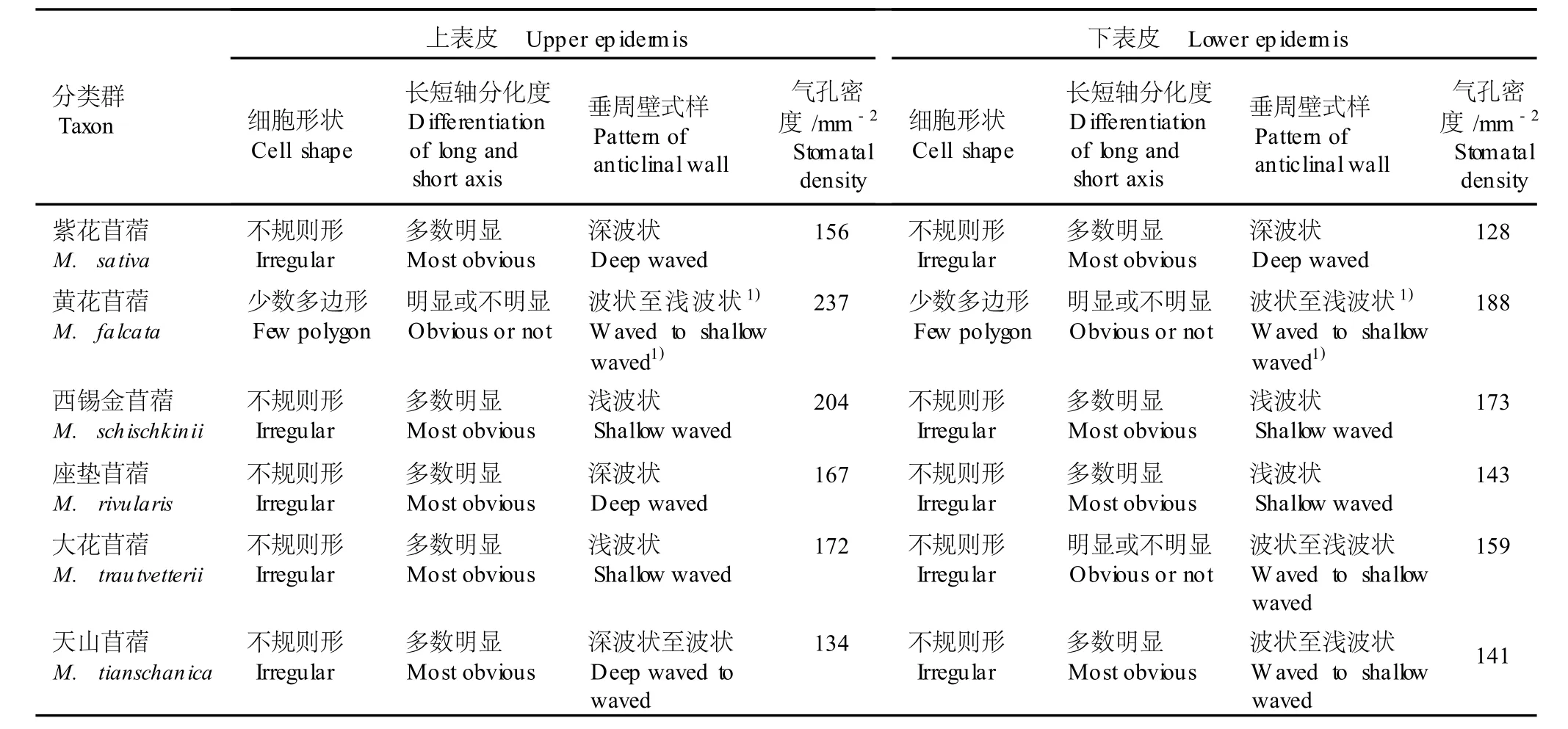
表 2 光学显微镜下紫花苜蓿复合体不同分类群叶表皮形态特征的观察结果Tab le 2 O bserva tion resu lt ofm orpholog ica l characters of lea f ep iderm is of d ifferen t taxa in M ed icago sa tiva com p lex by LM
2.3 紫花苜蓿复合体各分类群的叶片解剖结构特征
供试的紫花苜蓿复合体 6个分类群的叶片解剖结构特征见表 3和图版Ⅱ。6个紫花苜蓿复合体分类群的叶片解剖结构特征相似:叶片均为薄纸质型,厚度相差不大,为 130~170μm;上、下表皮细胞切面观为近圆形或近长方形。栅栏组织具有 1~2层细胞,厚度为 41~68μm;通常靠近表皮的第 1层栅栏组织细胞呈长圆柱状,排列相对紧密;第 2层栅栏组织细胞呈短圆柱状,排列较第 1层相对疏松。海绵组织厚度差别较大,为 32~75μm,细胞排列较疏松。
表 3显示,紫花苜蓿复合体 6个分类群叶片细胞组织结构的差异较明显,其中组织疏松度最大的为大花苜蓿(57.46%),最小的为紫花苜蓿 (24.85%),前者是后者的 2.31倍;组织紧缩度最大的为紫花苜蓿(51.70%),最小的为大花苜蓿 (31.56%),前者是后者的 1.64倍。
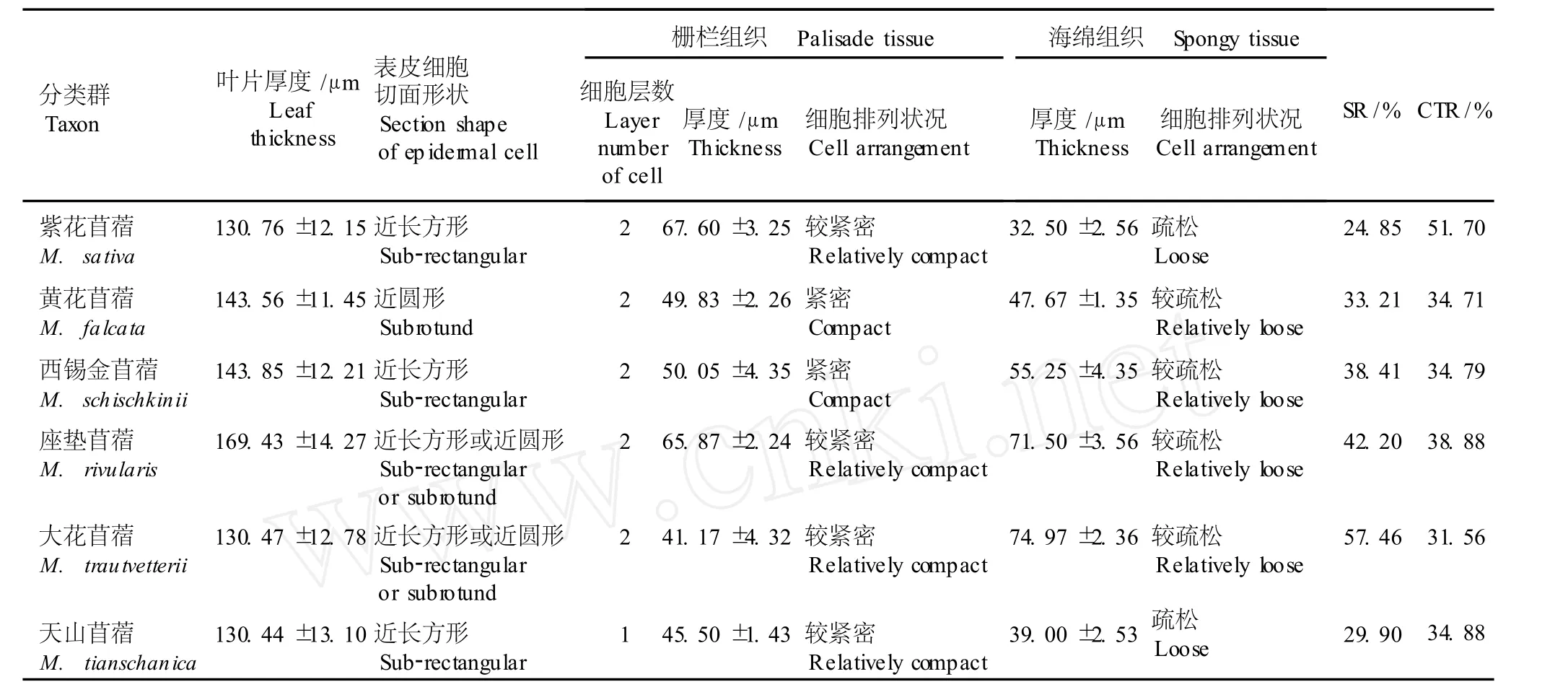
表 3 紫花苜蓿复合体不同分类群叶片解剖结构特征的观察结果1)Tab le 3 O bserva tion resu lt of leaf ana tom ica l structure fea tures of d ifferen t taxa in M ed icago sa tiva com p lex1)
2.4 基于紫花苜蓿复合体各分类群叶片表征形态性状的聚类分析结果
基于 15个叶片表征形态性状的紫花苜蓿复合体各分类群的UPGMA聚类分析结果见图 1。应用结合线法[13]划分类群时,在结合过程中,前 4次结合水平变化平缓,而第 5次结合水平迅速提高,产生了 1次飞跃。如果取飞跃前后 2次结合水平的平均值 1.53 (欧氏距离)作为划分类群的标准,则可得到 2支,其中黄花苜蓿独立为一支,其余 5个分类群聚成另一支。此处的结合水平可以作为种的界线。从划分结果看,中国境内的紫花苜蓿复合体应包含 2个种,即黄花苜蓿和紫花苜蓿。当结合水平取1.18(欧氏距离)时,第 2支又被分成 2个亚支,其中一个亚支包含紫花苜蓿和天山苜蓿,另一个亚支则包括西锡金苜蓿、座垫苜蓿和大花苜蓿,由此可以看出它们的相似性程度极高,且与紫花苜蓿的距离更近。

图1 紫花苜蓿复合体不同分类群的 UPGM A聚类分析结果F ig.1 C luster ana lysis resu lt of UPGM A of d ifferen t taxa in M ed icago sa tiva com p lex
2.5 基于紫花苜蓿复合体各分类群叶片表征形态性状的主成分分析结果
基于 15个叶片表征形态性状的紫花苜蓿复合体各分类群的主成分分析 Q分布图见图 2,结果显示,主成分分析Q分布图的结果与基于欧氏距离进行的聚类分析结果有较高的一致性。

图 2 紫花苜蓿复合体不同分类群主成分分析的 Q分布图F ig.2 Q d istr ibution m ap of pr incipa l com ponen t ana lysis of d ifferen t taxa in M ed icago sa tiva com p lex
主成分分析结果 (表4)表明,前3个主成分的累计贡献率达 85.14%,足以代表全部性状所提供的信息。第 1主成分的贡献率达 53.55%,是认识紫花苜蓿复合体的主要方面,各性状的绝对权重值主要是表皮细胞形状、垂周壁式样和气孔密度的差异性反映,实际情况有 2种不同的倾向:一种表现为表皮细胞形状不规则、垂周壁波状至深波状、气孔密度较小;另一种表现为部分表皮细胞规则、垂周壁浅波状至平直、气孔密度较大。第 2主成分的贡献率为 19.13%,主要反映了叶片内部组织结构的致密性。第 3主成分的贡献率为 12.46%,主要反映了表皮细胞的轴性分化特征。因此,对于紫花苜蓿复合体各分类群而言,叶片的表皮细胞形状、垂周壁式样、轴性分化特征、组织疏松度和气孔密度等性状具有较好的分类价值。

表 4 紫花苜蓿复合体叶部特征的主成分分析结果1)Tab le 4 Resu lt of pr incipa l com ponen tana lysisof leaf character istics of M ed icago sa tiva com p lex1)
3 讨论和结论
中国境内的紫花苜蓿复合体在组成上相对简单,《中国植物志》记载该复合体包括 3种 1变种[14],而文献[9]记载该复合体包括 3种 2变种 6变型,这两种处理结果都将多变苜蓿置于种的层面上,只是后者又从中划分出多个变型。分类学家大都认为多变苜蓿是Medicagosativa和M.falcata的杂交产物,具有较为完整的形态渐变,因而分类困难。就本文涉及的多变苜蓿的次级分类群而言,前苏联的有关植物志[1-2]常将该种处理成许多种,如:将分布于高加索、中亚细亚、西伯利亚和中国新疆一带的相关天然杂交类群 分 成M.schischkinii、M.tianschanica、M. agropyretorum、M.subdicycla、M.trautvetterii和M. rivularis等。欧美学者习惯倾向于生物学种,允许较大的形态变异尺度,故将这些变异类型都归入多变苜蓿,并做为异名处理[15],由他们确认的紫花苜蓿复合体不仅包括M.sativassp.×varia(M artyn)A rcang.,甚至将黄花苜蓿〔M.sativassp.falcata(L.)A rcang.〕也囊括其中。相比之下,中国学者的处理意见比较折中。作者基于紫花苜蓿复合体不同分类群的叶部特征、应用聚类分析和结合线法划分的分类结果与传统的分类结果有所差别。结合水平取 1.53,可得到 2个分支,即黄花苜蓿和以紫花苜蓿为代表的一支,进一步的划分结果表明紫花苜蓿是一个多型种。这 2个分支的性别亲和能力强,基因交流容易,并且杂交后代的生活力丝毫没有削弱的迹象,如果仅从性别亲和力上考虑,理所当然可以认为它们是同一种。但是,高等植物间的性别隔离远没有动物那样严格,并且在形态上 2个分支的差异又极为显著,因此作者认为将它们处理为 2个不同的种可能更加妥当。
因表型变异幅度极大,多变苜蓿的归属问题一直是分类学上的难题。从聚类分析的树系图看,多变苜蓿(本文中供试的为其异名的西锡金苜蓿、座垫苜蓿、天山苜蓿和大花苜蓿)与紫花苜蓿归在一类,说明多变苜蓿与紫花苜蓿的相似性水平较之与黄花苜蓿明显要高,这与李拥军等[10-11]的研究结果一致,但它们在形态特征上仍然存在一定的差异,这一点与《中国植物志》上的分类处理是一致的,所以,作者认为将多变苜蓿处理为独立的种有一定的合理性。
综上所述,作者认为多变苜蓿起源于黄花苜蓿和紫花苜蓿的杂交,叶部形态特征更接近于紫花苜蓿。国产的紫花苜蓿复合体包含 3个分类群,即紫花苜蓿、黄花苜蓿及多变苜蓿。从叶片的形态特征聚类分析结果看,前苏联学者曾作为种级单位命名的一些杂交后代分类群(如西锡金苜蓿、座垫苜蓿、天山苜蓿和大花苜蓿等)的性状分化不稳定,它们都应属于多变苜蓿的同种异名植物。
基于不同相似性系数所得到的聚类分析结果存在差别,而基于欧氏距离的聚类分析结果与主成分分析中的Q分析结果较吻合,因而,根据欧氏距离进行的聚类分析结果更加可靠,可作为划分紫花苜蓿复合体分类群的基础。
[1]Grossheim A A.Flora of the USSR:Vol.11[M].Moscow-Leningrad:USSR,1945:129-176.
[2]Pavlov N V.Flora of Kazakhstan:Vo l.5[M].A lma-A ta:Academ y of Science of Kazakhstan SSR,1961:30-50.
[3]Small E,Lefkovitch L P.Agrochemotaxmetry of alfalfa[J]. Canadian Journalof Plant Science,1982,62:919-928.
[4]Small E,B rookesB S.Taxonom ic circum scrip tion and identification in theMedicagosativa-falcata(alfalfa)continuum[J].Econom ic Botany,1984,38(1):83-96.
[5]QuirosC F,Bauchan GR.The genusMedicagoand theorigin of theMedicagosativacomp lex[M]∥Hanson A A,BarnesD K,H ill R R.Agronom y Monographs:No.9.M adison:American Society of Agronom y,1988:93-124.
[6]Small E,Jomphe M.A synopsis of the genusMedicago(Legum inosae)[J].Canadian Journalof Botany,1989,67(11):3260 -3294.
[7]何咏松,吴仁润.苜蓿自交不亲和性研究[J].中国草业科学, 1987,4(4):6-11.
[8]崔大方,田允温,闵继淳,等.新疆苜蓿属植物的分类研究[M]∥于兆英.西北地区现代植物分类学研究:第一卷.北京:科学技术文献出版社,1992:43-57.
[9]耿华珠.中国苜蓿[M].北京:中国农业出版社,1995:10-24.
[10]李拥军,苏加楷.中国苜蓿地方品种亲缘关系的研究:Ⅰ种子贮藏蛋白标记[J].草业学报,1999,8(1):31-41.
[11]李拥军,苏加楷.中国苜蓿地方品种亲缘关系的研究:ⅡRAPD标记[J].草业学报,1999,8(3):46-53.
[12]孙 毅,梁爱华,王景雪,等.根据核糖体DNA ITS序列分析苜蓿属的系统分类[J].西北植物学报,2003,23(2):242-246.
[13]徐克学.数量分类学[M].北京:科学出版社,1994.
[14]中国科学院中国植物志编辑委员会.中国植物志:第四十二卷第二分册[M].北京:科学出版社,1994:304-328.
[15]Lesins K A,Lesins I.GenusMedicago(Legum inosae),a Taxogenetic Study[M].Hague:D rW Junk Publisers,1979.
猜你喜欢
干旱地区农业研究(2022年1期)2022-01-28 04:51:30
现代畜牧科技(2021年9期)2021-10-13 06:38:52
现代畜牧科技(2021年4期)2021-07-21 06:13:04
装备制造技术(2019年12期)2019-12-25 03:06:52
科技创新与品牌(2019年12期)2019-02-06 04:03:29
中国三峡(2017年4期)2017-06-06 10:44:22
为了孩子(3~7岁)(2016年6期)2016-05-14 03:09:17
中国铸造装备与技术(2015年5期)2015-12-10 10:23:33
原子与分子物理学报(2015年1期)2015-11-24 12:49:26
食品科学(2013年15期)2013-03-11 18:25:51
