
翘嘴鳜NADPH氧化酶三个调节亚基cDNA的克隆及表达特征
2010-12-25 06:40:14胡宝庆文春根
Zoological Research 2010年3期
胡宝庆,刘 毅,文春根,*
(1. 南昌大学 生命科学学院,江西 南昌 330031;2. 江西师范大学 生命科学学院,江西 南昌 330022)
翘嘴鳜NADPH氧化酶三个调节亚基cDNA的克隆及表达特征
胡宝庆1,刘 毅2,文春根1,*
(1.南昌大学 生命科学学院,江西 南昌330031;2.江西师范大学 生命科学学院,江西 南昌330022)
吞噬细胞NADPH氧化酶能生成用于清除病原微生物的活性氧簇 (reactive oxygen species, ROS),在机体的防御体系中起着非常重要的作用。该文利用RT-PCR结合RACE-PCR的方法,对翘嘴鳜 (Siniperca chuatsi) NADPH氧化酶的3个调节亚基p40phox、p47phox和p67phox的cDNA进行了克隆。结果显示p40phox基因cDNA序列全长为1 406 nt,开放阅读框长度为1 050 nt,翻译成349个氨基酸;p47phox 基因cDNA序列全长为1 686 nt,开放阅读框为1 209 nt,翻译成402个氨基酸;p67phox基因cDNA序列全长为2 185 nt,开放阅读框长度为1 488 nt,翻译成495个氨基酸。半定量PCR分析显示在翘嘴鳜血液、脑、鳃、性腺、心脏、头肾、肠、肾、肝、脾、胸腺组织中都能检测到这3个亚基的mRNA表达,然而,它们在不同组织中的表达强度具有差异。经柱状黄杆菌灭活苗FKG4免疫后,p40phox亚基mRNA在翘嘴鳜血液和头肾中的表达量显著上升,p47phox在头肾和脾脏中的表达量显著上升,而p67phox在血液、头肾和脾脏中的表达量均显著上升。由此推断NADPH氧化酶参与了翘嘴鳜机体的抗菌免疫应答。
翘嘴鳜;NADPH氧化酶;分子克隆;柱状黄杆菌
NADPH氧化酶主要存在于中性粒细胞及其它吞噬细胞的细胞膜上,能够产生活性氧(reactive oxygen species, ROS),具有杀菌作用,并参与宿主细胞的免疫应答(Babior, 1999)。它由位于细胞膜上和细胞质内的多种蛋白成分组成。细胞膜上的NADPH氧化酶成分是异源二聚体跨膜蛋白,称为细胞色素b558,由gp91phox和p22phox两个催化亚基组成(Nauseef, 2004)。细胞质中的成分包括p40phox,p47phox,p67phox亚基及一个小分子量GTP结合蛋白Rac,这些亚基对于酶的活化起着调节作用,吞噬细胞受到细胞因子、激素或病原微生物刺激时,它们会装配到细胞色素b558上,形成具有完整生物活性的多酶复合体(Nauseef, 2004)。人(Homo sapiens) (Rooyer-Pokora et al, 1986; Volpp et al, 1989; Leto et al, 1990; Wientjes et al, 1993)、鼠(Mus musculus) (Bjorgvinsdottir et al, 1996; Jackson et al, 1994; Mizuki et al, 1998)、野牛(Bison bison) (Gauss et al, 2002)和海豚(Tursiops truncatus) (Inoue et al, 2000, 2001)细胞膜上和细胞质中NADPH氧化酶的5个亚基均已被克隆和描述。
除Rac外,鱼类NADPH氧化酶的5个亚基的cDNA也已从红鳍东方鲀(Takifugu rubripes) (Inoue et al, 2004)、鲤鱼(Cyprinus carpio)(Mayumi et al, 2008)中克隆。此外,在NCBI数据库中搜索到虹鳟(Oncorhynchus mykiss)和香鱼(Plecoglossus altivelis)的这5个亚基的cDNA全序列以及大西洋鲑(Salmo salar)的gp91phox、p47phox、p67phox亚基的cDNA全序列。
翘嘴鳜(Siniperca chuatsi)在我国分布广泛,具有较高经济价值,享有“淡水石斑”之誉。柱状黄杆菌(Flavobacterium columnare)是引起淡水鱼类烂鳃病发生的病原体。本文克隆了翘嘴鳜细胞质中NADPH氧化酶的3个调节亚基基因cDNA全长,并利用灭活柱状黄杆菌菌苗(FKG4)对鳜鱼进行免疫,检测这些亚基mRNA在鳜鱼不同组织中的表达情况,以期为了解鱼类呼吸爆发的机制及柱状黄杆菌疫苗的免疫途径奠定基础。
1 材料与方法
1.1 动物处理、FKG4的制备及免疫注射
翘嘴鳜购自武汉市江夏区牛山湖渔场,为人工网箱养殖,实验前,将购得的鱼(450±50)g在实验室自动充气水循环系统中人工饲养14 d,水温保持(25±1) °C,每日投喂小杂鱼作为饲料,选取健康的鱼作实验用。
取25 ℃振荡培养36 h的柱状黄杆菌培养液100 mL,10 000g离心10 min,沉淀用无菌PBS (pH7.2) 清洗3次,用含1.0%福尔马林的PBS重悬,室温下灭活24 h,10 000g离心10 min,PBS清洗两次,离心,用无菌PBS稀释到约1.0×108cell s/mL,置于4 °C冰箱备用。
免疫注射前,将FKG4菌苗取出升温至室温,实验鱼用MS222溶液(1 mg/L)麻醉,分为2个组,每组6条,一组每尾鱼经胸鳍下注射0.1 mL的FKG4,另一组则每尾注射同等剂量的灭菌PBS作为对照组,然后继续饲养7 d取样。
1.2 主要试剂及菌株
Trizol试剂购自Invitrogen公司,Ex-Taq聚合酶,DL-2000分子质量标准,PMD-18T载体、pTA2载体均购自TaKaRa公司;胶回收试剂盒购自Omego公司,其它试剂均为国产分析纯。菌株F. columnare G4由中国科学院水生生物研究所提供,大肠杆菌(Escherichia coli)DH5α由本实验室保存。
1.3 引物的设计与合成
根据已报道的红鳍东方豚和魟鳟的NADPH氧化酶p40phox、p47phox和p67phox亚基cDNA序列设计简并引物,特异性引物根据扩增出来的中间片断设计,所用引物均由上海华诺公司合成(表1)。
1.4 总RNA的提取和SMART cDNA的合成
分别取翘嘴鳜头肾和脾脏混合,按Trizol试剂盒说明书介绍的方法提取组织的总RNA。 琼脂糖凝胶电泳分析RNA的质量。按照Clontech公司的SMART RACE cDNA Amplification Kit方法进行第一链cDNA合成。具体操作按说明书进行。
1.5 各亚基中间片段及全长cDNA的扩增
根据表1所列引物,以第一链cDNA做为模板,先用简并引物扩增出基因的cDNA片段,再根据各个基因对应的特异性引物,用5′-RACE引物和SMART 5′锚定引物(5′-AAGCAGTGGTAACAACGCAGAGTACGCGGG-3′)扩增基因的5′端,用3′-RACE引物和SMART 3′锚定引物(5′-AAGCAGTGGTAACAACGCAGAGTACT30VN-3′)扩增基因的3′端。扩增产物经克隆、测序后,拼接基因的5′端和3′端序列,获得各基因的全长cDNA序列。
1.6 基因的氨基酸推测和序列比较分析
将测序所得到的cDNA序列用NCBI网站(http: //www.ncbi.nlm.nih.gov/blast)的BLASTN、BLASTX和TBLASTX软件进行同源基因的搜索,查找与其相关的同源序列;氨基酸序列的推断和疏水性分析通过ExPASy网站上(http://www.expasy.org/)的软件完成;氨基酸序列同源性比较由ClustalW1.8程序完成。 结构域分析用SMART软(http://smart.emblheidelberg.de)预测,按照MEGA 3.1中的邻接法(neighbor-jointing method)构建基于推导氨基酸的无根系统发生树,树的可靠性以1 000次bootstrap重复来评判。
1.7 组织表达与实时荧光PCR
利用反转录PCR方法检测翘嘴鳜NADPH氧化酶各调节亚基在鱼体组织中的表达。提取健康鱼血液、脑、鳃、性腺、心脏、头肾、肠、肾、肝、脾、胸腺组织的总RNA反转录成cDNA,然后将它们稀释成相同的浓度,并以其为模板进行PCR反应。反应产物的电泳条带的亮度强弱指示各亚基基因表达的高低。
用实时荧光PCR方法定量分析翘嘴鳜NADPH氧化酶各调节亚基在经FKG4免疫注射后的鱼体组织中表达变化。免疫注射FKG4后7 d提取免疫组和对照组翘嘴鳜血液、鳃、头肾、肠及胸腺和脾脏中的总RNA,逆转录成cDNA。用Real-time PCR引物扩增各基因序列(表1),PCR产物用DNA胶纯化试剂盒纯化后克隆进入载体pTA2。质粒DNA经纯化后用于制作标准浓度曲线,曲线的斜率应在−3.6与−3.2之间,相关系数应大于0.96。cDNA质粒溶液的浓度以OD260光密度值下进行测定,相应的基因拷贝数按1 µg的1 000 bp DNA约等于9.1×109分子的公式进行换算(Overbergh et al, 2003)。定量实时荧光PCR在Chromo4 Real-Time (MJ Research)上进行;分别将翘嘴鳜NADPH氧化酶3个调节亚基在各组织中的基因拷贝数除以β-actin基因的拷贝数以得到均一化,所得比值用来进行统计分析,免疫组和对照组之间特异基因表达的差异用t-检验分析,P<0.05为显著性差异。
2 结 果
2.1 翘嘴鳜NADPH氧化酶各调节亚基的cDNA全长及特征
翘嘴鳜NADPH氧化酶3个调节亚基cDNA的全长序列登录号为DQ341372 (p40phox)、DQ341374 (p47phox)和DQ341373 (p67phox)。p40phox亚基cDNA序列全长为1 406 bp,开放阅读框长度为1 050 bp,5′UTR长度为138 bp,3′UTR长度为218 bp包含mRNA不稳定信号、加尾信号和poly(A)尾各1个。由cDNA序列推断的蛋白包含349个氨基酸,预测相对分子质量为40.4 k,理论pI为6.50。p47phox亚基cDNA序列全长为1 686 bp,开放阅读框长度为1 209 bp,5′UTR长度为296 bp包含1个微卫星序列(AC),3′UTR长度为181 bp包括mRNA不稳定信号、加尾信号、 poly(A)尾各1个。该cDNA序列编码402个氨基酸,预测其相对分子质量为46.2 k,理论pI为9.27。 p67phox亚基cDNA序列全长为2 185 bp,开放阅读框长度为1 488 bp,5′UTR长度为59 bp,3′UTR长度为638 bp包括1个mRNA不稳定信号、3个加尾信号和1个poly(A)尾。该cDNA序列翻译成495个氨基酸,预测其分相对子质量为55.4 k,理论pI为5.81。3个cDNA序列推断的蛋白质序列均无信号肽和跨膜区。
2.2 翘嘴鳜与其他动物的对应调节亚基的氨基酸序列比对及进化树分析
用BLAST程序对翘嘴鳜NADPH氧化酶3个调节亚基基因编码的蛋白质进行同源性搜索(表2)。翘嘴鳜NADPH氧化酶的3个亚基与红鳍东方豚和虹鳟的同源亚基的氨基酸序列同源性在65.9%~82.5%之间,而与人、鼠、野牛、海豚等哺乳动物的对应亚基同源性在44.2%~55.8%之间。
运用ExPASy网站的软件分析各亚基的结构与功能域发现, 翘嘴鳜的p40phox蛋白含有PX 结构域(His18-Thr138)、SH3结构域(Pro172-Lys227)、PB1(Leu246-Ala339)结构域各1 个。P47phox蛋白序列包括PX结构域(Asn4-Arg121)和SH3结构域(Glu200-Gly255) 各1个。p67phox蛋白包括3个TPR基序(motif) (Ser37-Leu70;Ala71-Asn104;Cys121-Ala154)、1个PB1结构域(Gln347-Lys425)和2个SH3结构域(Glu243-Glu298;Ile436-Pro491)。另外,通过CLUSTAL比对显示这些亚基的结构域与人、鼠、野牛、海豚等哺乳动物对应的亚基蛋白质序列的结构域和功能位点基本上匹配(图1—3),说明这些亚基的功能与哺乳动物对应的亚基相似。
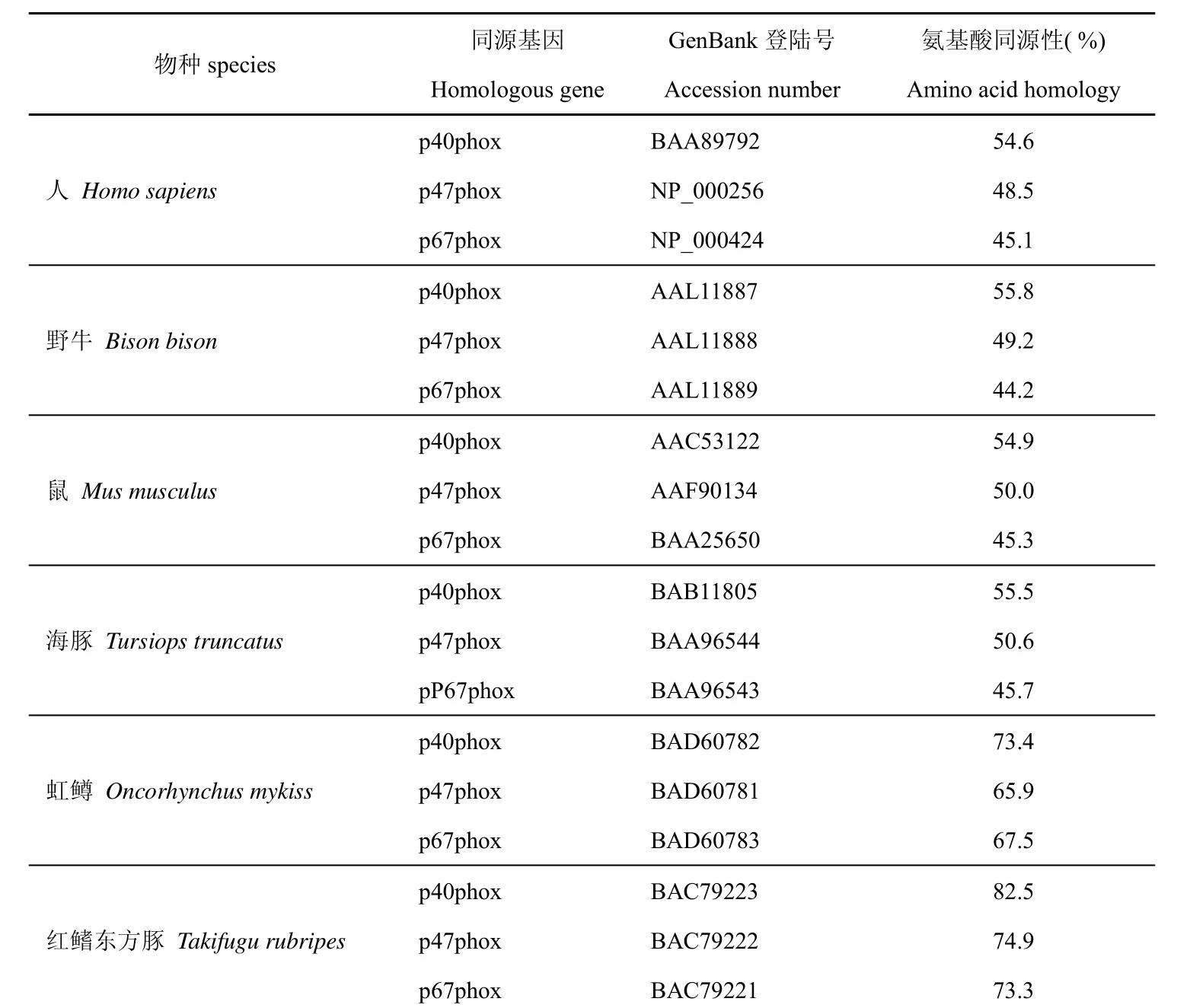
表 2 由翘嘴鳜p40phox、p47phox和p67phox亚基的cDNA推断的氨基酸序列与其他动物对应亚基氨基酸比较Tab. 2 Comparison of the amino acid homology among decuced amino acid sequences of p40phox, p47phox and p67phox subunit cDNA from Siniperca chuatsi and homologous genes from other species

图 1 翘嘴鳜p40phox与其他脊椎动物p40phox之间的氨基酸序列比对Fig. 1 Alignment of p40phox amino acid sequences of Siniperca chuatsi with that of other vertebrates

图 2 翘嘴鳜p47phox 与其他脊椎动物p47phox 之间的氨基酸序列比对Fig. 2 Alignment of p47phox amino acid sequences of Siniperca chuatsi with that of other vertebrates
对7种动物NADPH氧化酶3个调节亚基建立NJ系统树(图4a, b, c),结果显示鱼类独立于哺乳类动物之外,聚在同一枝上。翘嘴鳜NADPH氧化酶3个调节亚基在进化上与红鳍东方鲀的亲缘关系最近。
2.3 翘嘴鳜p40phox、p47phox和p67phox亚基组织表达及免疫刺激后的基因表达
翘嘴鳜NADPH氧化酶3个调节亚基在鱼体血液、脑、鳃、性腺、心脏、头肾、肠、肾、肝、脾、胸腺组织中的基因表达强度不同(图5)。在11种组织中都能检测到各亚基的基因表达。p40phox在肝脏组织中表达最弱,在其他10种组织中表达都较强。p47phox在头肾、鳃、心脏、脾组织中表达较强,在其余7种组织中的表达较弱;p67phox在头肾、鳃、脾组织中表达较强,在血液、脑、性腺、心脏、肠、肾、胸腺组织中表达较弱,然而,在肝脏组织中几乎检测不到。
经FKG4免疫后,翘嘴鳜3个调节亚基在血液、鳃、头肾、肠、脾及胸腺组织中的表达也有差异(图6)。在对照组鳜鱼的头肾中,3个亚基的表达量都很高。经过免疫注射7d后,p40phox在鳜鱼血液和头肾组织中的表达显著上升;p47phox在头肾和脾组织中的表达显著上升;p67phox在血液、头肾和脾组织中的表达显著上升;然而,3个调节亚基在鳃、肠及胸腺中的表达均无显著地变化。
3 讨 论
翘嘴鳜p40phox氨基酸序列长度比红鳍东方鲀p40phox长1个氨基酸,比虹鳟p40phox短9个氨基酸,但比人、野牛、鼠、海豚等哺乳动物p40phox长10个氨基酸;p47phox的氨基酸序列长度比红鳍东方鲀p47phox短21个氨基酸,比虹鳟p47phox短9个氨基酸,比人、野牛、鼠和海豚等哺乳动物p47phox分别长12、10、12和11个氨基酸;p67phox的氨基酸序列长度与红鳍东方鲀p67phox相同,比虹鳟p67phox短9个氨基酸,比人、野牛、鼠、海豚等哺乳动物p67phox分别短21、22、20、21个氨基酸。虽然翘嘴鳜NADPH氧化酶的3个调节亚基p40phox、p47phox和p67phox的氨基酸序列与人、鼠、野牛及海豚的相应亚基长度不一,且相似性较低,但是它们都有类似的结构和功能域以及相互作用位点。人的p40phox和p47phox的PX结构能有选择地识别PtdIns(3)P和PtdInsP2,这有助于它们在活化后被锚定到细胞膜上,进而装配成有活性的多酶复合体,Arg57、Arg58、Tyr59、Arg105残基对于维持这一功能是必要的(Nauseef, 2004; Karathanassis et al, 2002)。这一结构在翘嘴鳜以及其它动物内都存在,并且这些位点均保守。人的p40phox的SH3结构域能够与p47phox的C端的PRR结构相互作用,PC基序是与p67phox发生作用的部位 (Nauseef, 2004)。翘嘴鳜也像其他动物一样均具有类似的结构。人的p67phox的TPR结构与Rac亚基相互作用(Nauseef, 2004),这一结构在其他动物中均存在。由此推断翘嘴鳜与人的NADPH氧化酶亚基都拥有相似的活化机制。
哺乳动物p67phox亚基的脯氨酸富裕区(PRR)是与亚基p47phox相互作用的重要区域(Wientjes et al, 2003),而鱼类的该区域都不保守。由此推测在鱼类的p67phox亚基与p47phox亚基的相互作用可能比哺乳动物弱。人的p47phox的PX结构有两个可区分的碱性区域,在空间结构上呈两个口袋状,能识别磷脂酰肌醇,有助于p47phox在质膜上的锚定;Arg43、Phe44和Arg90是第一个碱性袋必需的;Arg70、Lys55和 His51对第二个碱性袋的存在是必需的,两个疏水性残基Ile65与Trp80在结合时插入到质膜或溶酶体膜上(Karathanassis et al, 2002; Stahelinet al, 2003)。虽然预测的翘嘴鳜p47phox蛋白的Ile65、Trp80残基和Arg70、Lys55、His51很保守,但是Arg43、Phe44残基分别被Thr 和Tyr残基替代。因此,推测翘嘴鳜p47phox的PX结构的第一个碱性袋可能不存在,表明p47phox结合到质膜或溶酶体膜上的能力比哺乳动物要弱。过去认为p47phox N端PX结构域中的PXXP结构域起着自动抑制的作用,它能与p47phox C端的SH3区域相互作用,掩盖了磷脂酰肌醇结合位点,因而阻止了细胞质亚基复合物与膜的结合(Hiroaki et al, 2001)。最近资料认为p47phox的两个SH3结构能形成一个小沟,p47phox的碱性区域被掩盖在这个小沟内,在细胞静息期,小沟中保守的Trp193和Trp263残基与碱性区域的核心结构(296~304残基)相互作用,从而起到自动抑制作用,这2个SH3对于维持这种作用都是必需的(Groemping & Rittinger, 2005)。红鳍东方鲀、虹鳟的p47phox像其他动物一样,都包含2个SH3结构域 (Inoue et al, 2004),但是翘嘴鳜p47phox仅有1个SH3结构,而且Trp193残基缺失,因此推测翘嘴鳜碱性区域与SH3结构域之间的相互作用可能不存在或很弱,它的分子间自动抑制作用可能是由PXXP与SH3区域相互作用造成。

图 3 翘嘴鳜p67phox与其它脊椎动物p67phox之间的氨基酸序列比对Fig. 3 Alignment of p67phox amino acid sequences of Siniperca chuatsi with that of other vertebrates

图 4 用7种动物p40phox(a)、p47phox(b)、p67phox(c)亚基氨基酸序列构建的NJ系统树Fig. 4 Phylogenetic tree inferred from amino acid sequences of subunits of p40phox(a),p47phox(b),p67phox(c) from seven animals

图 5 翘嘴鳜p40phox, p47phox, p67phox亚基在不同组织中的基因表达Fig. 5 Expression of Siniperca chuatsi p40phox, p47phox, p67phox transcripts in a variety of tissues

图 6 翘嘴鳜p40phox (a)、p47phox (b)及p67phox (c)在免疫鱼(■)与对照鱼(□ ) 6种织中的表达Fig. 6 Expression of Siniperca chuatsi p40phox (a), p47phox (b) and p67phox (c) in 6 tissues of vaccinated (■) and control fish (□)
磷酸化作用被认为是NADPH氧化酶活化过程中的一个关键的事件。大多数的亚基(除Rac和gp91phox)在活化过程中都显示不同程度的磷酸化,人的p47phox有11处磷酸化位点,分别位于丝氨酸303、304、310、315、320、328、345、348、359、370和379(Groemping & Rittinger, 2005)。p40phox有2处磷酸化位点,位于Thr154 和Ser315(Groemping & Rittinger, 2005),p67phox仅确定Thr233发生磷酸化(Groemping & Rittinger, 2005)。翘嘴鳜的这些亚基绝大多数磷酸化位点都很保守,然而,p40phox的Gly321代替了相应位点的Ser315;p47phox的Ser345、Ser359分别被Ala、Lys代替;p67phox中Thr233被Glu233取代,由此表明翘嘴鳜NADPH氧化酶3个亚基的磷酸化作用比哺乳动物弱。
鱼类脾脏的椭圆体、头肾中的造血组织、心脏的围心腔及血液等组织中含有较为丰富的吞噬细胞(Ellis, et al, 1976)。鳃、肠作为鱼类主要的黏膜相关淋巴组织(mucosa-associated lymphoid tissue),也含有较为丰富的吞噬细胞 (McMillan et al, 1997; Lin et al, 1998)。这些吞噬细胞都能够产生呼吸爆发(respiratory burst) (Lin et al, 1999; Clerton et al, 1998)。因此,翘嘴鳜的3个亚基的mRNA在头肾、脾脏、血液中表达量较高,这与鲫鱼和红鳍东方鲀的表达模式是一致的(Inoue et al, 2004; Mayumi et al, 2008)。即使在同一组织中,三个亚基的mRNA表达量也有些差异,可能是由于各亚基基因的转录时间差异造成的。利用LPS体外刺激人或红鳍东方鲀的粒细胞6~24 h后,NADPH氧化酶的各亚基中,均只有p47phox的mRNA表达显著上升,其他亚基的mRNA表达无明显变化(Inoue et al, 2004; Cassatella et al, 1991)。然而,用FKG4对翘嘴鳜进行免疫刺激后发现,不仅p47phox在头肾、脾中的表达显著上升,而且p40phox和p67phox在头肾、脾和血液中的表达也显著上升。这可能是由于活体免疫刺激了鱼体的这些主要免疫器官,从而导致组织中粒细胞含量增多;在鳃、肠、胸腺中的表达变化不显著,可能是由于这些组织在鱼体免疫系统中处于次要地位。
致谢:本研究得到了中国科学院水生生物研究所聂品实验室的帮助,在此表示感谢!
Babior BM. 1999. NADPH oxidase: An update [J]. Blood,93(5): 1464-1476.
Bjorgvinsdottir H, Zhen L, Dinauer MC. 1996. Cloning of murine gp91phox cDNA and functional expression in a human Xlinked chronic granulomatous disease cell line [J]. Blood,87(5): 2005-2010.
Cassatella MA, Bazzoni F, Amezaga MA, Rossi F. 1991. Studies on thegene expression of several NADPH oxidase components[J]. Biochem Soc Trans,19(1): 63-67.
Clerton P, Troutaud D, Deschaux P. 1998. The chemiluminescence response of leucocytes isolated from the gut of rainbow trout(Oncorhynchus mykiss) [J]. Fish Shellfish Immunol,8: 73-76.
Ellis AE, Munroe ALS. 1976. Defence mechanisms in fish. 1. A study of the phagocytic system and the fate of intraperitoneally injectedparticulate material in the plaice (Pleuronectes platessa L.) [J]. J Fish Biol,8: 67-78.
Gauss KA, Bunger PL, Siemsen DW, Young CJ, Nelson-Overton L, Prigge JR, Swain SD, Quinn MT. 2002. Molecular analysis of the bison phagocyte NADPH oxidase: Cloning and sequencing of five NADPH oxidase cDNAs[J]. Comp Biochem Physiol B: Biochem Mol Biol,133(1): 1-12.
Groemping Y, Rittinger K. 2005. Activation and assembly of the NADPH oxidase:a structural perspective [J]. Biochem J,386(Pt3): 401-416.
Hiroaki H, Ago T, Ito T, Sumimoto H, Kohda D. 2001. Solution structure of the PX domain, a target of the SH3 domain [J]. Nat Struct Biol,8(6): 526-530.
Inoue Y, Itou T, Jimbo T, Sakai T, Ueda K, Imajoh-Ohmi S, Iida T. 2000. Molecular cloning and identification of bottle-nosed dolphin flavocytochrome b gp91phox and p22phox subunits[J]. Vet Immunol Immunopathol,76(1-2): 137-150.
Inoue Y, Itou T, Jimbo T, Sakai T, Ueda K, Imajoh-Ohmi S. 2001. Molecular cloning and identification of bottle-nosed dolphin p40(phox), p47(phox) and p67(phox) [J]. Vet Immunol Immunopathol,78(1): 21-33.
Inoue Y, Suenaga Y, Yoshiura Y, Moritomo T, Ototake M, Nakanishi T. 2004. Molecular cloning and sequencing of Japanese pufferfish (Takifugu rubripes) NADPH oxidase cDNAs[J]. Dev Comp Immunol,28(9): 911-925.
Jackson SH, Malech HL, Kozak CA, Lomax KJ, Gallin JI, Holland SM. 1994. Cloning and functional expression of the mouse homologue of p47phox [J]. Immunogenetics,39(4): 272-275.
Karathanassis D, Stahelin RV, Bravo J, Perisic O, Pacold CM, Cho W, Williams RL. 2002. Binding of the PX domain of p47(phox) to phosphatidylinositol 3,4-bisphosphate and phosphatidic acid is masked by an intramolecular interaction [J]. EMBO J,21(19): 5057-5068.
Leto TL, Lomax KJ, Volpp BD, Nunoi H, Sechler JM, Nauseef WM, Clark RA, Gallin JI, Malech HL. 1990. Cloning of a 67-kD neutrophil oxidase factor with similarity to a noncatalytic region of p60c-src [J]. Science,248(4956): 727-730.
Lin SH, Davidson GA, secombes CJ. 1998. Morphological study of cells isolated from the perfused gill of dab and Atlantic salmon [J]. J Fish Biol,53:560-568.
Lin SH, Ellis AE, Davidson GA, secombes CJ. 1999. Migratory,respiratory burst and mitogenic responses of leucocytes isolated from the gills of rainbow trout(Oncorhynchus mykiss) [J]. Fish Shellfish Immunol,9: 211-226.
Mayumi M, Takeda Y, Hoshiko M, Serada K, Murata M, Moritomo T, Takizawa F, Kobayashi I, Araki K, Nakanishi T, Sumimoto H. 2008. Characterization of teleost phagocyte NADPH oxidase: molecular cloning and expression analysis of carp (Cyprinus carpio) phagocyte NADPH oxidase[J]. Mol Immunol,45(6): 1720-1731.
McMillan DN, Secombes CJ. 1997. Iso1ation of rainbow trout (Oncorhynchus mykiss) intestinal intraepithelial lymphocytes(IEL) and measurement of their cytotoxic activity [J]. Fish Shellfish Immunol,7: 527-541.
Mizuki K, Kadomatsu K, Hata K, Ito T, Fan QW, Kage Y, Fukumaki Y, Sakaki Y, Takeshige K, Sumimoto H. 1998. Functional modules and expression of mouse p40phox and p67phox, SH3-domain-containing proteins involved in the phagocyte NADPH oxidase complex [J]. Eur J Biochem,251(3): 573-582.
Nauseef WM. 2004. Assembly of the phagocyte NADPH oxidase [J]. Histochem Cell Biol,122(4): 277-291.
Overbergh L, Giulietti A, Valckx D, Decallonne R, Bouillon R, Mathieu C. 2003. The use of real-time reverse transcriptase PCR for the quantification of cytokine gene expression[J]. J Biomol Tech,14(1): 33-43.
Rooyer-Pokora B, Kunkel LM, Monaco AP, Goff SC, Newburger PE, Baehner RL, Cole FS, Curnutte JT, Orkin SH. 1986. Cloning the gene for an inherited human disorder-chronic granulomatous disease-on the basis of its chromosomal location[J]. Nature,322(6074): 32-38.
Stahelin RV, Burian A, Bruzik KS, Murray D, Cho W. 2003. Membrane binding mechanisms of the PX domains of NADPH oxidase p40phox and p47phox [J]. J Biol Chem,278(16): 14469-14479.
Volpp BD, Nauseef WM, Donelson JE, Moser DR, Clark RA. 1989. Cloning of the cDNA and functional expression of the 47-kilodalton cytosolic component of human neutrophil respiratory burst oxidase [J]. Proc Natl Acad Sci USA,86(18): 7195-7199.
Wientjes FB, Hsuan JJ, Totty NF, Segal AW. 1993. p40phox, a third cytosolic component of the activation complex of the NADPH oxidase to contain src homology 3 domains [J]. Biochem J,296(Pt3): 557-561.
Wientjes FB, Segal AW. 2003. PX domain takes shape [J]. Curr Opin Hematol,10(1): 2-7.
cDNA Cloning and Expression Characterization of Three Regulatory Subunits of NADPH Oxidase Within Cytoplasm from Mandarin Fish,Siniperca chuatsi
HU Bao-Qing1, LIU Yi2, WEN Chun-Gen1,*
(1. College of Life Sciences, Nanchang University, Nanchang 330031, China;
2. College of Life Sciences, Jiangxi Normal University, Nanchang 330022, China)
The phagocyte NADPH oxidase plays a crucial role in host defense against invading microorganisms by catalyzing the formation of reactive oxygen species, which is the precursor of a variety of microbicidal oxidants such as hydrogen peroxide (H2O2). In the present study, full-length cDNAs of three regulatory subunits of NADPH oxidase, including p40phox, p47phox, p67phox were cloned from head kidney of mandarin fish utilizing the reverse transcription polymerase chain reaction and rapid amplification of cDNA ends. Sequence analysis showed that the full length cDNA of p40phox is 1 406 nt, containing a 1 050 nt open reading frames that encodes a 349 amino acid protein, the full length cDNA of p47phox is 1 686 nt, containing a 1 209 nt open reading frames that encodes a 402 amino acid protein, the full length cDNA of p40phox is 2 185 nt, containing a 1 488 nt open reading frames that encodes a 495 amino acid protein. Semi-quantitative RT-PCR analyses from various tissues indicated that mRNAs of the three subunits can be detected in the blood, brain, heart, spleen, kidney and thymus, but their expression intensity are different in tissues. Stimulating the mandarin fish with formalin killedFlavobacterium columnareG4 significantly up-regulated the expression of p40phox in blood and head kidney; and p47phox in head kidney and spleen; and p67phox in blood, head kidney and spleen. The results suggested that mandarin NADPH oxidase was involved in the immune responses against bacteria.
Siniperca chuatsi; NADPH oxidase; Molecular cloning;Flavobacterium columnare
胡宝庆(1972-),男,江西波阳人,讲师,主要从事水产动物疾病研究。E-mail:baoqinghu@sina.com
Q78; Q959.483; Q959.483.06
A
0254-5853-(2010)03-0250-11
10.3724/SP.J.1141.2010.03250
2009-09-14;接受日期:2009-12-21
国家自然科学基金资助项目(30960296);江西省科技攻关项目(2004)
*通讯作者(Corresponding author),文春根 (1963−),男,汉族,江西南昌人,教授,主要从事水产动物疾病方面的研究。E-mail: cgwen@ncu.edu.cn
猜你喜欢
垂钓(2023年10期)2024-01-02 08:47:05
昆明医科大学学报(2020年12期)2021-01-26 00:44:12
世界最新医学信息文摘(2020年68期)2020-12-25 11:55:27
科学养鱼(2020年4期)2020-06-05 07:33:46
动物营养学报(2016年9期)2016-10-14 07:47:23
中国粮油学报(2016年5期)2016-01-23 02:44:40
吉林大学学报(医学版)(2015年4期)2015-12-17 07:48:13
海洋科学(2014年2期)2014-03-13 15:00:17
茶叶通讯(2014年2期)2014-02-27 07:55:39
中国医学科学院学报(2013年6期)2013-03-11 20:26:01
