
高体革鯻5种同工酶的组织特异性研究
2010-11-27 02:49张龙岗董学飒付佩胜
长江大学学报(自科版) 2010年5期
杨 玲,安 丽,张龙岗,董学飒,付佩胜
(山东省淡水水产研究所, 山东 济南 250117)
高体革鯻5种同工酶的组织特异性研究
杨 玲,安 丽,张龙岗,董学飒,付佩胜
(山东省淡水水产研究所, 山东 济南 250117)
采用聚丙烯酰胺垂直梯度凝胶电泳法,分析了高体革鯻(Scortumbarcoo)的心脏、肝脏、脾脏、肾脏、肌肉、眼睛等6种组织中的5种同工酶(乳酸脱氢酶、苹果酸脱氢酶、酯酶、过氧化物酶、过氧化氢酶)的表达情况,发现这5种酶在表型、分布和活性上均表现出明显的组织特异性,且与各组织执行的生理机能相一致。
高体革鯻(Scortumbarcoo);同工酶;组织特异表达
高体革鯻(Scortumbarcoo)为原产于澳大利亚的淡水鱼类,隶属于鲈形目(Perciformes)鯻科(Terapontidae)革鯻属 (Scortum)。鱼体呈纺锤形,体侧有一至数个黑色晶莹的椭圆形斑块及零星分布的小斑块,宛如黑色的宝石,因而得名“宝石鲈”。因其外形美观,可食部分比例大,无肌间骨,具有较强的环境适应力、抗病力强、生长速度快等优点,适于池塘及网箱养殖,故被引进世界各地。目前,国内学者已经对高体革鯻的生物学特性、生长发育、养殖技术、营养成分分析、核型分析等进行过研究[1~5],但未见有关其同工酶方面的报道。为此,本研究采用聚丙烯酰胺垂直板不连续电泳的方法,对高体革鯻的肝脏、脾脏、心脏、肾脏、肌肉、眼睛等6种组织的5种同工酶——乳酸脱氢酶(LDH)、苹果酸脱氢酶(MDH)、酯酶(EST)、过氧化物酶(POD)、过氧化氢酶(CAT)进行了分析,以期为高体革鯻的生化遗传结构研究、种质资源鉴定以及遗传多样性分析提供可借鉴的参考依据。
1 材料与方法
1.1 材料
试验用鱼取自山东省淡水水产研究所,规格为体长25~30 cm,体重525~670 g的高体革鯻成鱼。
1.2 方法
(1)样品制备 共取成鱼30尾,剪断尾动脉和鳃,在流水中放血以减少组织带血。活体解剖,迅速摘取眼睛、心脏、肾脏、脾脏、肝脏、肌肉等组织,用预冷的0.8%生理盐水将各组织冲洗干净,取适量称重,放入预冷的研钵中,用消毒的剪刀适当剪碎,加入预冷的0.01 mol/L的pH 7.4的磷酸缓冲液(组织∶缓冲液=1 g∶3 mL),置冰浴中研磨匀浆,匀浆液置4 ℃冰箱抽提1~2 h后,高速冷冻离心机(4 ℃)15 000 r/m离心30 min,取上清液,加入等体积的40%的蔗糖溶液,分装后置 -20 ℃保存备用。
(2)电泳及染色 采用聚丙烯酰胺垂直梯度凝胶电泳法,分离胶浓度(T)为7%或7.5%,浓缩胶浓度为4%或3%。电泳缓冲液为Tris-Gly ,高离子强度电泳缓冲液(pH 8.3)或低离子强度电泳缓冲液(pH 8.7)。梯度胶的浓度和缓冲系统见表1,点样量为20 μL,以溴酚蓝作指示剂,电泳在4 ℃冰箱进行。同工酶的染色方法参照文献[6~9]稍作改进。同工酶的命名和分析参照文献[9~11],以各酶带的相对迁移率(Rf)从小到大依次命名,即向阳极泳动最快的编为1号,其余依次顺序编为2,3,4,…,同一基因位点的不同等位基因按照从阴极到阳极的顺序依次标记为a、b、c,…。
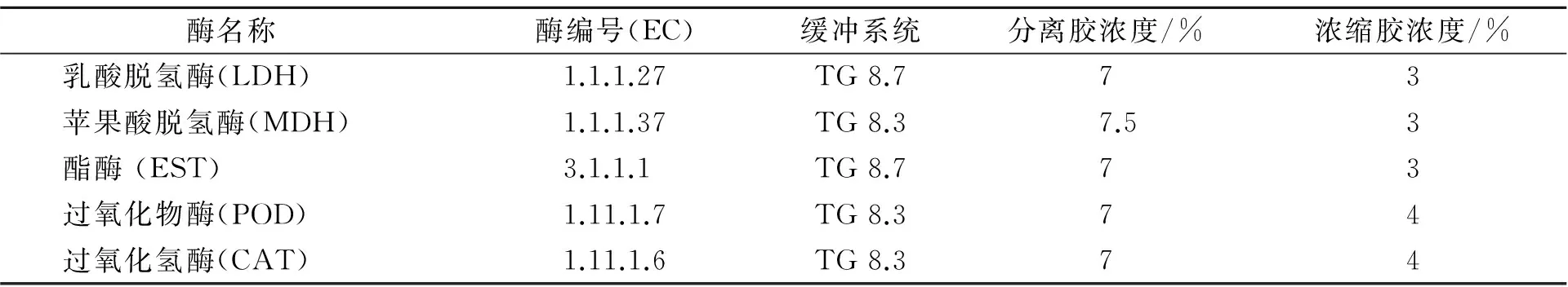
表1 用于分析高体革鯻同工酶的名称、编号、缓冲系统和凝胶浓度Table 1 The name ,serial number ,buffer system and gel concentration for analyzing the isozyme of S. barcoo
注:TG 8.3 指Tris-甘氨酸高离子强度电泳缓冲液(pH 8.3);TG 8.7指Tris-甘氨酸低离子强度电泳缓冲液(pH 8.7)
2 结果与分析
2.1 过氧化物酶(POD,E.C.1.11.1.7)

S:脾脏;K:肾脏;L:肝脏;M:肌肉;H:心脏; E:眼睛图1 高体革鯻不同组织的POD电泳图谱Figure 1 Electrophoretogram of POD in various tissues of S. barcoo

S:脾脏;K:肾脏;L:肝脏;M:肌肉;H:心脏; E:眼睛图2 高体革鯻不同组织的EST电泳图谱Figure 1 Electrophoretogram of EST in various tissues of S. barcoo
高体革鯻不同组织的POD电泳图谱见图1。由图1可见,其 POD由5个基因座位编码(Pod-1、2、3、4、5),其中Pod-1、Pod-2和Pod-5为多态位点,Pod-1在脾脏、肾脏中表达为由同一位点编码的2条同二聚体酶带,在心脏中检测到1条酶带,即Pod-1a。Pod-1在心脏 、脾脏中含量较高,在肾脏中含量稍低,在肝脏、肌肉和眼睛中没有检测到。Pod-2在脾脏、肾脏和肝脏中有表达,在心脏、肌肉和眼睛中没有检测到酶的活性,其中Pod-2a仅在脾脏和肾脏中表达,Pod-2b和Pod-2c在脾脏、肾脏和肝脏中有表达,肝脏中含量较少。除肌肉组织外,其他组织中都有Pod-3和Pod-4的表达,心脏和眼睛的含量较少,脾脏和肾脏中的含量较高,眼睛中Pod-3和Pod-4酶带的迁移速度较其他3种组织中的快。Pod-5仅在脾脏和肾脏中有表达,其他组织中均没有检测到其活性,在脾脏中含量较高。
在肌肉组织中没有检测到POD活性,其他各组织均表达为2~8条酶带。其中脾脏和肾脏中POD的活性最强,染色也最深。酶谱分析表明,高体革鯻6种组织中没有任何2种组织具有完全相同的POD表达情况,说明高体革鯻的POD同工酶有较为显著的组织特异性。
2.2 酯酶(EST,E.C.3.1.1.1)
高体革鯻不同组织的EST电泳图谱见图2。由图2可见,EST在各组织中都有表达,共检测到5条酶带。Est-1在肝脏和肾脏中表达最强,脾脏中的表达次之,在心脏中表达很弱,在眼睛和肌肉中没有表达。Est-2也是在肝脏和肾脏中表达最强,脾脏中的表达次之,在心脏、眼睛和肌肉中没有表达。Est-3和Est-4在各组织中均有表达,但是表达的强度不一,Est-3在肝脏和肾脏中表达最强,脾脏中次之,心脏和眼睛中较弱,肌肉中表达最少。Est-4在肾脏、眼睛和心脏中表达较强,在脾脏和肝脏中较弱,在肌肉中表达最弱。Est-5只在肾脏中有表达,在其他组织中没有检测到其活性。
2.3 苹果酸脱氢酶(MDH,E.C.1.1.1.37)
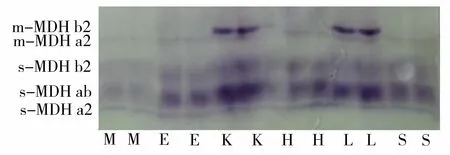
S:脾脏;K:肾脏;L:肝脏;M:肌肉;H:心脏; E:眼睛图3 高体革鯻不同组织的MDH电泳图谱Figure 3 Electrophoretogram of MDH in various tissues of S. barcoo

S:脾脏;K:肾脏;L:肝脏;M:肌肉;H:心脏; E:眼睛图4 高体革鯻不同组织的CAT电泳图谱Figure 4 Electrophoretogram of CAT in various tissues of S. barcoo

S:脾脏;K:肾脏;L:肝脏;M:肌肉;H:心脏; E:眼睛图5 高体革鯻不同组织的LDH电泳图谱Figure 5 Electrophoretogram of LDH in various tissues of S. barcoo
硬骨鱼类的MDH为二聚体,存在互不形成异聚体的上清液型(s-MDH)和线粒体型(m-MDH)2部分,均是由2个基因编码的二聚体酶[12]。高体革鯻不同组织的EST电泳图谱见图3。MDH在高体革鯻中可以检测到这2种类型的酶带。靠近阳极的为s-MDH型,表达为3条酶带,靠近阴极的为m-MDH型,表达为1~2条酶带。MDH在各组织中的表达存在差异,在肝脏和肾脏中的含量最丰富,心脏、脾脏、眼睛等组织的含量次之,在肌肉中的表达最少。m-MDHb2只在肝脏和肾脏中表达,其他组织中没有表达。 s-MDH在6种组织中的表达基本一致,在肝脏和肾脏中表达最强,在肌肉中表达最弱。
2.4过氧化氢酶(CAT,E.C.1.11.1.16)
高体革鯻不同组织的CAT电泳图谱见图4,可见在高体革鯻脾脏中CAT的活性最强,在肝脏和肾脏中CAT的活性较弱,在心脏、肌肉和眼睛中没有检测到其活性。由此可见,高体革鯻的CAT同工酶存在较为明显的组织差异。
2.5乳酸脱氢酶(LDH,E.C.1.1.1.27)
高体革鯻不同组织的LDH电泳图谱见图5,共检测到5条酶带,其中靠近阳极的有3条酶带。LDH-1和LDH-2在肝脏中表达最强,心脏和肌肉中的表达较弱,其他组织中没有表达;LDH-3在各组织中都有表达,在肾脏和心脏中的表达最强,肝脏和眼睛中的表达次之,肌肉和脾脏中表达最弱;LDH-4只在肾脏和肝脏中表达,其他组织中没有表达;LDH-5只在心脏中有表达。可见,高体革鯻的LDH同工酶具有显著的组织特异性。
3 结论与讨论
3.1高体革鯻同工酶的表达具有明显的组织特异性
高体革鯻的6种组织中,肌肉共检测到3种酶,MDH、EST和LDH,其含量比在其他组织中明显要低。眼睛和心脏一样,除CAT以外的其他4种酶都有表达,心脏中的表达量较高,眼睛中的表达量比较低。脾脏、肾脏和肝脏中这5种酶全部表达,相对而言,脾脏表达量较小,肝脏和肾脏中这5种酶的表达量是最多的。
从本研究可以看出,高体革鯻同工酶的组织特异性,表现在以下几方面:(1)位点的表达。某些位点的同工酶仅限于某种组织所特有,在其他组织中,该同工酶不表达或活性太低而检测不出。如m-MDH-b2仅在肝脏和肾脏中表达,其他组织中没有表达。LDH-5只在心脏中表达,LDH-4只在肝脏和肾脏中表达;ESt-5只在肾脏中表达。(2)位点表达的程度即酶活性的强弱不同。①同一位点在不同组织中的相对含量或表达的数量差别很大,如Pod-3和Pod-4在脾脏和肾脏中表达较强,其他4种组织中表达较弱或不表达。Est-2在肝脏和肾脏中表达较强,其他组织中较弱。②不同位点在不同组织中的相对活性不同,如心脏中的Pod-1的表达比其他位点要强,而其他组织的正好相反,Pod-2、Pod-3和Pod-4的表达都比Pod-1强。肾脏和肝脏中的EST-2的表达比其他位点强,而其他组织中的EST-4的表达最强。(3)等位基因表达的差异。由于复等位基因的存在,不同组织中会有不同的等位基因表达,结果同一位点的酶谱表型在不同组织中有明显的差异,或有时等位基因编码的亚基不一定完全表达或不能通过电泳鉴定其产物,即存在“哑等位基因”或“无效基因”,结果导致不同组织中同一位点的酶谱表型出现差异[13]。如Pod-2在脾脏和肾脏中有3条带,而在其他组织中有2条带或者不表达。眼睛中Pod-3和Pod-4的迁移速度比其他组织中的略快,可能是由于基因的突变或酶蛋白的修饰所造成的,是眼睛区别于其他组织的标记。
3.2 高体革鯻同工酶组织特异性与功能的相关性
(1)过氧化物酶(POD)可利用H202氧化供氢体,降解嘌呤、酚和胺等物质并减轻其毒性[14]。本研究也表明,在血流丰富、白细胞及吞噬细胞较多的脾脏、肾脏、肝脏、心脏中POD的表达较强,而在肌肉中没有表达。与青海湖裸鲤和鳜鱼的POD酶谱相似[15,16],而与岱衢族大黄鱼和道氏虹鳟的POD酶谱不同[17,18],岱衢族大黄鱼的心脏中没有检测到POD活性,道氏虹鳟肌肉中的POD表达量较高。
(2)酯酶(EST)是催化酯类化合物水解并进入中间代谢的重要酶类,对于酯类代谢和生物膜的结构与功能具有一定作用,还能催化酯键水解,有解毒作用[15]。本研究中EST同工酶在高体革鯻肝脏中表达最强,是与肝脏作为机体重要的解毒器官密切相关的,这和其他鱼类的酯酶分布情况类似[13,15~20]。EST同工酶在高体革鯻肾脏和脾脏中的表达也较强,与肾脏和脾脏作为主要的代谢和造血器官有关。这与褐点石斑鱼、赤点石斑鱼、岱衢族大黄鱼等类似,而和波纹唇鱼不同,后者在肾脏和脾脏中没有表达。高体革鯻肌肉中EST仅检测到Est-3和Est-4两条酶带,且染色较浅,说明其活性较弱。这和波纹唇鱼、岱衢族大黄鱼、赤点石斑鱼、鳜鱼等相似,和道氏虹鳟不同,后者肌肉中EST活性较强。本研究中Est-3和Est-4具有广泛的组织分布,在细胞内执行最基本的生理功能。EST-5只在肾脏中有表达,是肾脏组织的特征性酶带。
(3)苹果酸脱氢酶(MDH)是三羧酸循环中的一种酶,在细胞中的功能主要是使苹果酸脱氢参与糖酵解后的有氧代谢,因此,MDH在心脏、肝脏和肾脏等有氧代谢旺盛的组织中染色较深[13]。高体革鯻的MDH同工酶也分上清液型(s-MDH)和线粒体型(m-MDH)2种类型,6种组织都有s-MDH酶带,在肝脏和肾脏中活性最强,组织间差异不大;而m-MDH具有明显的组织特异性,在肝脏和肾脏中含量丰富,其他组织中含量较少或无。这与岱衢族大黄鱼的MDH酶谱类似[17],而与石斑鱼和道氏虹鳟不同[18~21],后者各组织中都存在m-MDH,而s-MDH酶带有显著的组织特异性。
(4)过氧化氢酶(CAT)为二聚体酶,能将体内的H2O2分解为H2O和O2,是生物防御体系的关键酶之一,存在于动物的肝和红细胞中,为机体提供了抗氧化防御机理[15]。本研究中高体革鯻脾脏中的CAT活性最强,肝脏和肾脏中也有表达,在其他组织中没有检测到其活性,这与脾脏作为造血和免疫的重要器官,肝脏作为机体重要的解毒器官,肾脏具有内分泌方面的功能,可以分泌肾素、前列腺素、激肽、促红细胞生成素,刺激骨髓造血等功能密切相关的。本研究和虹鳟及青海湖裸鲤的CAT酶谱有差异[15,22],CAT同工酶在青海湖裸鲤肝脏和肾脏中表达最强,心脏中较弱,肌肉中没有表达;虹鳟的眼睛中有CAT活性表达。
(5)乳酸脱氢酶(LDH)参与糖酵解,在无氧条件下能将糖酵解产生的丙酮酸还原为乳酸,产能以维持机体的能量需要。鱼类的LDH一般是由A、B、C 3个基因位点编码的四聚体酶,其中A、B位点存在于各种组织中,而C位点则局限于特定的组织器官中[23]。一般来说,LDH-B4主要位于好氧性的器官组织如心、脑中,LDH-A4主要位于厌氧性器官组织如肌肉、肝脏中[20]。
本研究结果表明,高体革鯻的LDH由3个基因座位编码,其中A和B 2个基因合成的亚基组合成3种不同的四聚体,其中LDH4对应为LDH-A4, LDH5对应为LDH-B4。LDH1、LDH2和LDH3是否为C位点表达的产物或是由LDH-A和LDH-B的复等位基因造成的还有待进一步研究。在鱼类中,A与B亚基的结合受阻,不能形成经典的5条酶带的现象是较多的[17,19~20],王金星等[24]认为,亚基的结合受阻所形成的酶带减少是进化过程中亚基相互结合的位点发生变异的必然结果。从高体革鯻的LDH酶谱可以看出,在肝脏和肾脏中LDH-4的活性最强,与该器官的厌氧代谢有关。LDH5只在心脏中有表达,与心脏的有氧代谢相关。肌肉和脾脏中LDH活性较弱。其他鱼类肌肉中的LDH同工酶活性较强[12,13,15,20],而本研究中高体革鯻肌肉的LDH活性却显得较弱,其原因还有待于进一步研究。
[1]叶奕佐,叶 嵘,孙文学,等.高体革鯻生物学特性初步观察[J].水产科技情报,2004,31(1):20 ~22.
[2]田海军,高 强,龙 勇,等.高体革鯻生物学特性和养殖技术[J].广西水产科技,2006,(4):46~47.
[3]骆豫江,朱新平,潘德博,等. 高体革鯻仔稚鱼的生长和发育[J].水产学报,2008,32(5):697~703.
[4]鲍 丹,陶宁萍,丁卓平,等.高体革鯻、鲈和鳜的营养成分分析比较[J].上海水产大学学报,2006,(1): 123~127.
[5]潘德博,骆豫江,朱新平,等.高体革鯻的形态特征及核型分析[J].大连水产学院学报,2009,24(2):185~188.
[6]周延清,杨清香,张改娜,等. 生物遗传标记与应用[M].北京:化学工业出版社,2008.47~69.
[7]Manchenko G P(华子纯,郑伟娟等译). 酶的凝胶电泳检测手册[M].北京:化学工业出版社,2008.49~290.
[8]郭尧君. 蛋白质电泳实验技术[M].北京:科学出版社,1999.70~78.
[9]胡书能,万贤国. 同工酶技术及其应用[M].长沙:湖南科学技术出版社,1985.70~85.
[10]熊全沫. 鱼类同工酶谱分析(上)[J].遗传,1992,14(2):41~44.
[11]熊全沫. 鱼类同工酶谱分析(下)[J].遗传,1992,14(3):47~48.
[12]姜建国,熊全沫,姚汝华,等.青鱼不同组织中同工酶的表达模式[J].水生生物学报,1997,21(4):353~358.
[13]区又君,齐旭东,李加儿,等.波纹唇鱼不同组织5种同工酶表达的差异[J].南方水产,2009,5(2):51~55.
[14]陈 康. 动物细胞中过氧化物酶体的功能[J].生物学通报,1995,30(1):18~19.
[15]孟 鹏,史建全,祁洪纺,等. 青海湖裸鲤同工酶表达的组织特异性分析[J].海洋水产研究,2008,29(5):112~118.
[16]杨军峰,周 乔,邵雪玲,等. 斑鳜和翘嘴鳜不同组织中的过氧化物酶、酯酶和乳酸脱氢酶的比较研究[J].水利渔业,2007,27(6):7~9.
[17]管丹东,李明云,叶帅东,等.岱衢族大黄鱼不同组织的同工酶谱[J].宁波大学学报(理工版),2008,21(1):34~38.
[18]顾 岩,孙中武,尹洪滨,等. 道氏虹鳟同工酶的组织分布及表达[J].东北林业大学学报,2008,36(2):58~64.
[19]尹绍武,廖经球,黄 海,等.褐点石斑鱼不同组织4种同工酶的研究[J].海洋通报,2007,26(1):472~474.
[20]邓思平,刘楚吾. 赤点石斑鱼4种同工酶的组织分布及基因位点分析[J].浙江海洋学院学报(自然科学版),2004,23(2):103~106.
[21]尹绍武,黄 海,廖经球,等.点带石斑鱼与褐点石斑鱼4种同工酶的比较研究[J].水产科学,2007,26(9):500~503.
[22]庞艳红,孙中武,尹洪滨,等. 5个虹鳟群体的生化遗传分析[J].水产学报,2007,31(4):539~544.
[23]薛国熊,官 平,张燕生,等. 草鱼同工酶比较酶学和免疫化学性质[J].水产学报,1992,16(4):357~363.
[24]王金星,周才武. 鱼类的乳酸脱氢酶同工酶[J].海洋湖沼通报,1989,(1):51~57.
2010-01-05
山东省农业良种工程项目(优质抗病速生鱼类良种选育)
杨 玲(1967-),女,山东济南人,研究员,主要从事遗传育种技术研究.
付佩胜,E-mail :jnsfu@126.com
10.3969/j.issn.1673-1409(S).2010.02.013
Q55;Q959.483
A
1673-1409(2010)02-S036-05
猜你喜欢
中老年保健(2022年3期)2022-08-24
现代临床医学(2021年6期)2021-11-20
宁夏医学杂志(2020年3期)2021-01-21
中国生殖健康(2019年10期)2019-01-07
特别健康(2018年9期)2018-09-26
临床医药文献杂志(电子版)(2017年11期)2017-05-17
当代医药论丛(2017年22期)2017-04-12
现代检验医学杂志(2016年5期)2016-08-20
中国卫生标准管理(2015年15期)2016-01-15
中国当代医药(2015年21期)2015-03-01
