
等离子体对小麦种子萌发过程中某些生理变化的影响
2010-09-19 09:04黄明镜
山西农业科学 2010年11期
黄明镜
(山西省农业科学院旱地农业研究中心,山西太原030006)
采用各种物理因子刺激农作物及其种子,可提高种子活力,促进种子萌发,并能提高种子抗逆能力以及农作物的产量和质量[1]。等离子体作为一种刺激因素,在农业科学领域,对作物的影响及其增产效应正在进行广泛探索[2]。
采用不同类型等离子体种子处理生物激活技术,在多种作物和蔬菜上,不仅可以促进作物萌发与生长,而且能够有效地改善作物的抗逆性和提高产量。黄明镜等[3]研究表明,用等离子体处理小麦、玉米、南瓜和萝卜等种子,发现其活力及抗旱性明显提高。尹美强等[4]研究表明,用等离子体处理番茄种子能增强番茄抗旱性,使根系活力和ATP含量增加,植物抗逆性提高。郝学金等[5]进行了用低温等离子体处理黄瓜、番茄、辣椒、甘蓝和萝卜等蔬菜种子的增产技术研究。王敏等[1]研究了大气压等离子体处理对黄瓜种子萌发及幼苗生长的影响。方向前等[6-7]研究表明,等离子体处理玉米、大豆种子能明显提高化肥利用率,增产效果显著。李学慧等[8]在蔬菜上的研究表明,等离子体处理有一定的增产效应,从而显示出了等离子体农业新技术的广阔前景。
迄今为止,关于等离子体对种子萌发及增产效应的影响研究报道较多,但对等离子体处理对种子萌发过程中的生理变化还不太清楚。
本试验对种子萌发起重要作用的α-淀粉酶活性、可溶性蛋白含量的变化、过氧化物酶活性等的变化情况进行了研究,以期揭示等离子体的作用机理。
1 材料和方法
1.1 材料处理和培养
选用抗旱性差异显著的小麦品种晋麦66号和晋麦47号为试验材料。将经挑选的籽粒饱满、大小均一的干种子用适宜的弧光等离子体处理,置于铺有2层滤纸的培养皿中,加入适量自来水,在20℃培养箱中培养。
1.2 α-淀粉酶的提取和测定
α-淀粉酶活性测定用3,5-二硝基水杨酸比色法[9]。称取1 g萌发的小麦种子,置于研钵中,磨成匀浆倒入量筒,用水稀释至50 mL,混匀后在室温下放置20 min。每隔数分钟振荡1次,然后以4 000 r/min冷冻离心10 min,取上清液备用。在520 nm的波长下比色,用麦芽糖含量表示α-淀粉酶的活性。
1.3 可溶性蛋白质的测定
采用考马斯亮蓝G-250比色法[9]。取发芽的小麦种子1 g,用50倍的0.1 mol/L磷酸缓冲液(pH=7)在冰浴下研磨提取,在4℃条件下以3 000 r/min离心10 min。提取液中加入考马斯亮蓝在595 nm处比色。
1.4 等离子体小麦种子萌发和幼苗生长过程中POD活性的变化
过氧化物酶活性测定用愈创木酚法[9],样品中过氧化物酶用硝酸钙溶液在冰浴中准确提取3 min,用愈创木酚与H2O2在20℃下反应15 min,用分光光度计在波长475 nm下测定其OD值。
2 结果与分析
2.1 等离子体处理对小麦萌发过程中α-淀粉酶活性的影响
种子的萌发是逐渐消耗贮藏物质的过程。小麦种子内淀粉含量多,而淀粉是在淀粉酶的作用下进行分解,因此,通过测定淀粉酶活性,能够反映种子萌发的快慢。小麦种子中α-淀粉酶活性随着萌发时间的变化如图1所示。
从图1可以看出,等离子体处理的小麦种子在萌发过程中,α-淀粉酶的活性显著高于对照,尽管品种特性不同,但是等离子体对其α-淀粉酶的促进效果较为一致,在种子萌发的第2天和第3天,晋麦66号的α-淀粉酶活性分别比对照高143.1%和115.5%,晋麦47号分别比对照高122.3%和60.6%,且达到了极显著水平。不同品种对等离子体的反映有所差别,晋麦66号对等离子体的响应表现出较高的敏感性,等离子体处理萌发24 h的α-淀粉酶活性就高出对照40.9%,随着萌发时间的延长,α-淀粉酶活性提高的幅度始终高于晋麦47号。
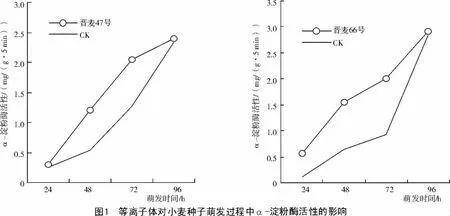
淀粉酶活性可反映种子萌发活力,等离子体处理后淀粉酶活性的提高,从机理上说明了其对种子萌发的促进效应。
2.2 等离子体处理对小麦萌发过程中可溶性蛋白含量的影响
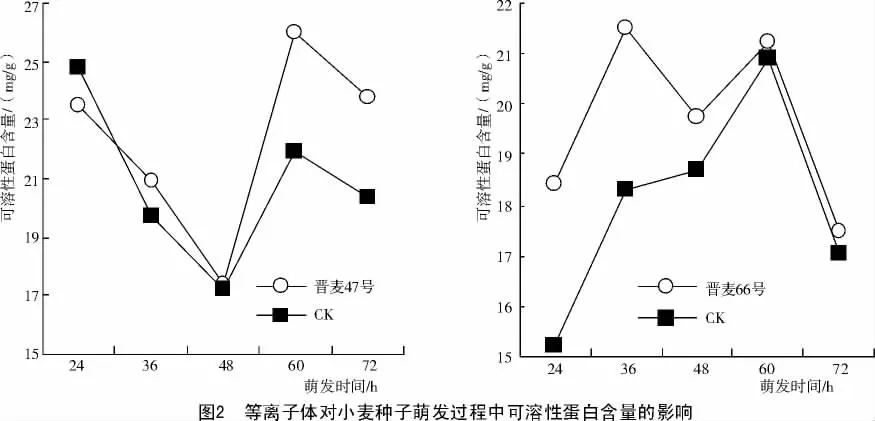
种子的萌发不但是贮藏物质的降解过程,同时也是各种酶即蛋白质逐渐合成的过程,因此,用可溶性蛋白含量的变化,可以反映萌发种子生化过程的快慢。小麦萌发过程中可溶性蛋白含量的变化如图2所示。
由图2可知,品种间可溶性蛋白含量随萌发时间的变化,曲线有很大差异,但是随萌发时间的延长,等离子体处理的可溶性蛋白含量始终比对照高。晋麦47号等离子体处理萌发24,36,48,60,72 h 分别比对照提高 -5.03%,5.78%,1.36%,18.56%和17.24%;晋麦66号等离子体处理萌发 24,36,48,60,72 h 分别 比 对照 提 高20.99%,17.28%,5.69%,1.74%和2.30%。
2.3 等离子体处理对作物种子萌发和幼苗生长过程中过氧化物酶活性的影响
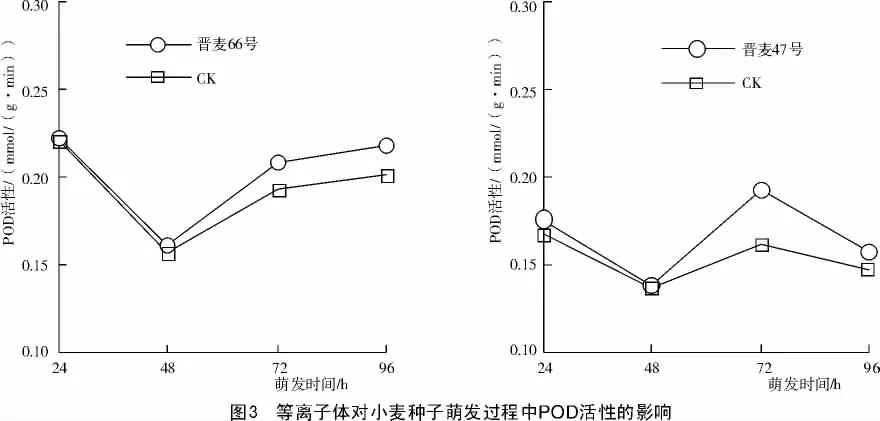
种子萌发过程中过氧化物酶的行为是种子萌发与生长进程的决定因子之一,是生物生长与分化的灵敏指标,等离子体诱导小麦种子在萌发过程中POD活性的提高,表明等离子体对小麦的萌发和生长有一定的促进效应。本研究分别对等离子体处理的小麦种子及幼苗的POD活性进行了测定,结果表明:在种子萌发初期,等离子体对POD活性的促进效应不明显,随萌发过程的推进,等离子体提高POD活性的效应逐渐加强。种子萌发第3天等离子体的促进效应达最高峰,晋麦66号种子的POD活性比对照提高10.5%,晋麦47号种子的POD活性比对照提高18.5%。随着种子萌发时间的延续,贮藏物质消耗殆尽,转为幼苗的生长阶段。品种之间种子萌发72 h之内差异不明显,72~96 h之间等离子体处理比对照的POD活性提高幅度最大,晋麦66号处理比对照提高8.3%~8.8%;晋麦47号处理较对照提高了6.6%~18.9%,此阶段晋麦47号的POD活性随萌发时间的延长有所下降,并且等离子体处理下降幅度较大,说明等离子体对不同作物种子萌发过程中POD活性的影响不一致(图3)。
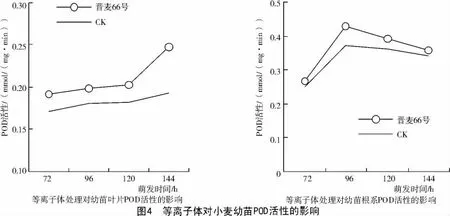
在萌发过程中对晋麦66号幼苗叶片和根系的POD活性进行测定,结果显示,等离子体处理后,小麦叶片的POD活性在其生长过程中始终高于对照,根系POD活性在96 h后虽然有所降低,但是等离子体处理POD活性仍高于对照。随着萌发时间的延长,等离子体处理小麦根系的POD活性比对照提高了4.8%~15.3%,处理后的叶片POD活性比对照提高了9.7%~27.9%(图 4)。
3 结论与讨论
种子是植物遗传信息的载体之一,其质量的好坏直接影响植株的生长发育和产量。等离子体处理对生物具有光激活效应,经等离子体处理后,种子细胞内的电子、离子、分子等带电离子与光的相互作用而使种子细胞吸收辐射能量[10],造成物体变热、电离、发生电性变化、生物学反应等,促使种子活化,提高种子活力和萌发能力,激活种子中各种酶的活性[11]和种子萌发期的代谢水平[12],促进作物的增产。种子萌发过程中,在各种酶的作用下,物质在不断的合成与降解,提供作物生长所需的物质和能量。本试验结果表明,等离子体处理提高了α-淀粉酶活性、过氧化物酶活性和可溶性蛋白含量,加强了种子的代谢水平,促进了作物的生长。但不同作物品种对等离子体的敏感性不同,其生理反应亦不同。
[1] 王敏,杨思泽,陈青云,等.大气压等离子体处理对黄瓜种子萌发及幼苗生长的影响[J].农业工程学报,2007,23(2):195-200.
[2] 马步洲,黄明镜.等离子体在农业上的应用[J].山西农业,2003(1):26.
[3] 黄明镜,马步洲,岳艳翠,等.等离子体对种子活力及抗旱性的影响[J].干旱地区农业研究,2002,20(4):65-68.
[4] 尹美强.弧光等离子体与梯度磁场结合处理番茄种子的生物效应[J].核聚变与等离子体物理,2006,26(1):75-80.
[5] 郝学金,秦建国.低温等离子处理研究初报[J].山西农业科学,1998,26(2):39-41.
[6] 方向前,边少锋,孟祥盟,等.等离子体处理玉米对化肥利用率的影响[J].中国农学通报,2006,22(3):203-205.
[7] 方向前,边少锋,付稀厚,等.等离子体处理大豆对化肥利用率的影响[J].中国农学通报,2007,23(6):392-395.
[8] 李学慧,曹阳,胡铁军,等.等离子体电磁处理对蔬菜产量的影响 [J].哈尔滨师范大学自然科学学报,2002,18(6):51-53.
[9] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[10] 余增亮.离子束生物技术引论[M].合肥:安徽科学技术出版社,1996.
[11] 夏丽华,张春玉.磁场处理对番茄种子活力及苗期长势的影响[J].东北师大学报(自然科学版),1999(3):66.
[12] 曹学成,程炳嵩,邹琦,等.脉冲磁场对小麦生理效应的初步研究 [J].山东农业大学学报(自然科学版),1998,29(3):345-350.
猜你喜欢
空间科学学报(2021年6期)2021-03-09
现代塑料加工应用(2021年5期)2021-02-28
中国粮油学报(2019年4期)2019-07-12
中成药(2018年8期)2018-08-29
新农业(2017年2期)2017-11-06
医学研究杂志(2015年9期)2015-07-01
中国当代医药(2015年9期)2015-03-01
食品工业科技(2014年15期)2014-03-11
食品工业科技(2014年9期)2014-03-11
食品工业科技(2014年7期)2014-03-11
