
整合酶及细胞辅助因子LEDGF与慢病毒整合
2010-09-10 04:24:30郝飞飞田晋红周建华
中国预防兽医学报 2010年3期
郝飞飞,田晋红,周建华
(1.西南大学药学院,重庆 400716;2.中国农业科学院哈尔滨兽医研究所,黑龙江哈尔滨 150001)
整合酶及细胞辅助因子LEDGF与慢病毒整合
郝飞飞1,2,田晋红1,周建华2*
(1.西南大学药学院,重庆 400716;2.中国农业科学院哈尔滨兽医研究所,黑龙江哈尔滨 150001)
慢病毒属逆转录病毒科,包括人免疫缺陷病毒(HIV-1和HIV-2)、猴免疫缺陷病毒(SIV)、马传染性贫血病毒(EIAV)、牛免疫缺陷病毒(BIV)、猫免疫缺陷病毒(FIV)、绵羊梅迪-维斯纳病毒(MVV)和山羊关节炎-脑炎病毒(CAEV)。其中EIAV是人类最早发现的慢病毒[1]。这些慢病毒成员在形态、基因组结构、细胞嗜性、病毒的生活周期、病毒复制的分子机制,尤其是病毒的整合机制等方面相似。近些年来,由于HIV-1对人类健康的危害,慢病毒复制的各个环节倍受关注,期望找出能够控制HIV-1复制的靶点,抑制其在体内的复制。慢病毒基因组整合至宿主染色体和基因的高度易变性使得慢病毒的治疗和疫苗研发异常困难。因此,整合酶是慢病毒研究热点之一。
1 整合酶的结构与功能
慢病毒的整合酶具有3个相同的保守结构域。以的HIV-1的整合酶为例,由病毒pol基因末端编码的288个氨基酸组成,包含3个不同结构和功能的结构域,即N端结构域(N terminal domain,NTD)、催化核心区结构域(Catalytic core domain,CCD)和 C端结构域(C terminal domain,CTD)。每个结构域直接或间接地促进整合酶和DNA的整合[2]。
1.1 整合酶的结构 以HIV-1为例,该酶的NTD包括1 aa~46 aa,通过一个接头片段(47 aa~55 aa)与CCD连接。NTD具有一个在所有慢病毒的整合酶和逆转录过程中都保守的HTH基序,包括两对保守氨基酸残基-组氨酸残基12和16以及半胱氨酸残基40和43[3-4],该基序可以和Zn2+结合形成锌指结构并且促进整合酶的功能性寡聚化[5-6]。与Zn2+结合的残基突变会促使整合酶单体的产生并且抑制3'端加工和链转移反应[7]。此外,最近生物化学和结构生物学的研究表明,NTD和CCD的相互作用会发生在整合酶的功能性四聚化中。若将NTD和CCD相互作用表面的14位丙氨酸替换成赖氨酸,则使整合酶四聚体变得不稳定,并降低整合酶的催化活性[8-9]。
CCD是整合酶的催化活性中心,是多聚核苷酸转移酶超级家族的成员之一,与细菌的核糖核酸酶H有共同的骨架和相似的催化机制[10]。CCD的催化位点是由保守的3个氨基酸残基-天冬氨酸残基64和116,谷氨酸残基152组成,主要作用是与二价金属离子结合,对整合酶的催化起到辅助作用[11-12]。体外突变和细胞内突变这些氨基酸残基会严重降低内整合酶的活性[7-13]。实验证明,Mg2+和Mn2+对酶与DNA底物的反应是必要的,由于Mg2+在细胞内的相对浓度较高,被认为是生理学辅助因子。整合酶利用相同的催化位点催化3'端加工和链转移反应[14]。因此,CCD很可能是结合病毒和靶DNA的结合位点。对HIV整合酶的晶体结构解析发现,CCD由6个α螺旋和5个β片层组成,包括3个保守的氨基酸残基形成一个D,DX35E基序[14],可以与金属离子直接结合。突变其中的氨基酸残基,会严重地降低酶的功能[32]。
HIV-1的CTD结构域富含碱性氨基酸残基,由5个β片层组成,属于类SH3结构域[15],而通常具有相似折叠的蛋白会以非特异性方式结合到DNA的小沟处[16-17]。CTD是整合酶3个结构域中最为不保守的区域,被认为是与DNA底物结合的区域。实验证明,C末端241位和242位的亮氨酸残基突变为丙氨酸残基会影响整合酶二聚体的形成,从而减弱整合酶的催化活性[18]。
1.2 整合酶的功能 慢病毒侵染宿主并融合入细胞(图1A),经脱壳后,病毒RNA在逆转录酶的作用下逆转录成cDNA,并与整合酶一起完成3'端加工(3'processing)和链转移(Strand transfer)反应这2个过程,将病毒的DNA整合入宿主基因组中(图1B)。
1.2.1 3'端加工作用首先,整合酶以二聚体的方式与病毒的长末端重复序列(LTR)结合[2]。LTR包含了一个保守的CAGT序列,整合酶通过水解磷酸二酯键分别切下碱基GT,从而露出碱基CA,形成了可与染色体DNA相匹配的交错切口[19]。此后,连接在cDNA两端的2个整合酶二聚体形成四聚体,在胞浆中与宿主因子形成整合前复合物(Pre-integration complex,PIC)。这个过程称为3'端加工。
1.2.2 链转移反应在3'端加工之后,PIC进入细胞核,整合酶交错切割宿主细胞染色体DNA,产生间隔5个碱基的交错切口,然后病毒DNA的3'端带自由羟基的碱基与宿主DNA的5'端共价连接,并由细胞的DNA修复酶将病毒DNA 5'末端与宿主DNA 3'末端补齐,完成整合过程[19]。
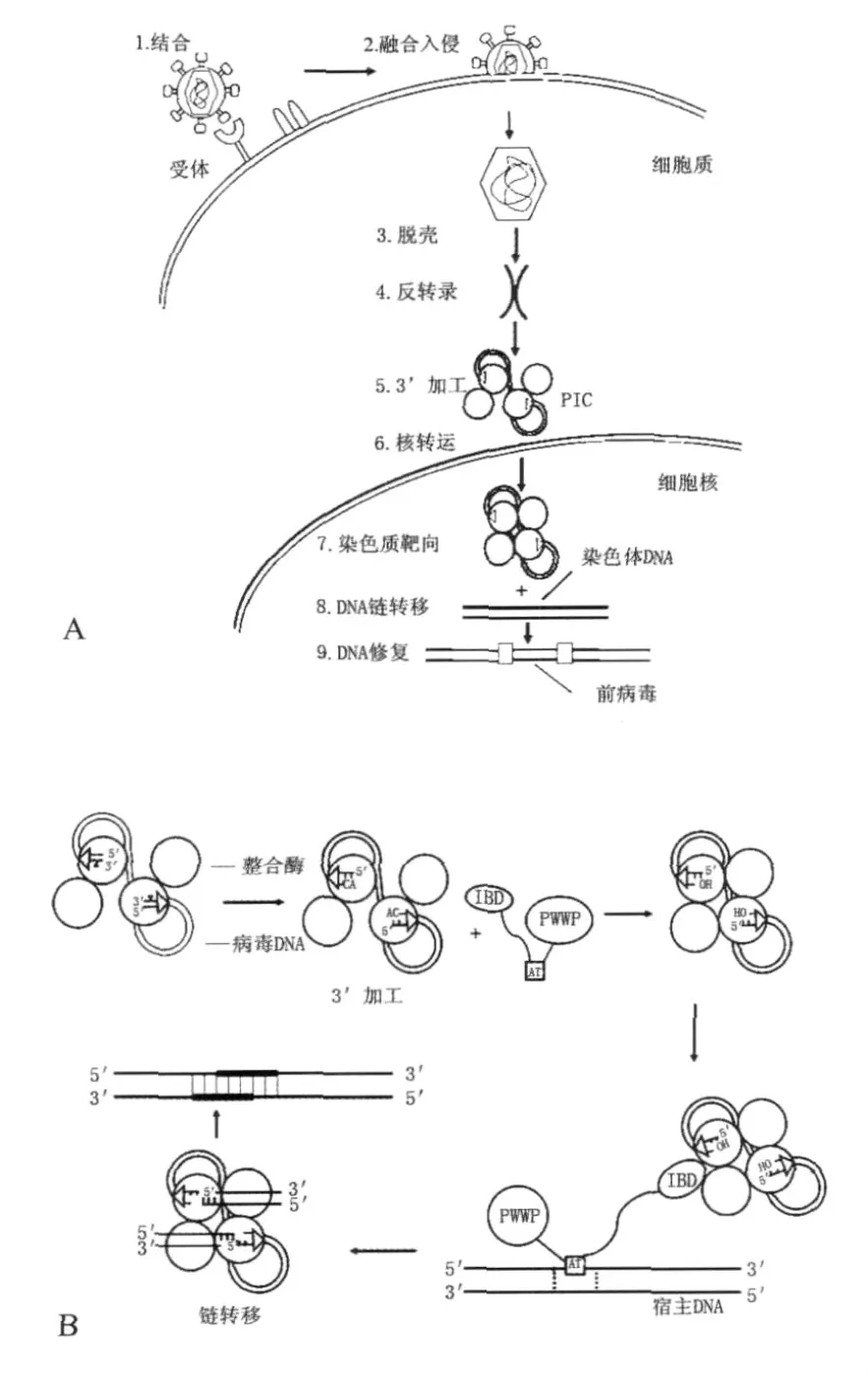
图1 慢病毒侵染细胞过程和逆转录后DNA的整合机制
2 LEDGF/p75的主要结构
LEDGF与已发现的转录协同激活因子p75为同一种蛋白,因此也称为LEDGF/p75。LEDGF是普遍存在于细胞核内的蛋白,在细胞周期过程中与染色体关系紧密[20-21]。LEDGF主要通过一个位于C末端的小的α螺旋结构域-整合酶结合结构域(Integrase-binding domain,IBD)与慢病毒的整合酶相互作用。除了IBD结构域以外,全长LEDGF蛋白还包含其它结构和功能性部分:N端的PWWP结构域,可以介导蛋白质-DNA和蛋白质-蛋白质相互作用;核定位信号区(Nuclear-localization signal,NLS),主要决定LEDGF的核定向转运,参与LEDGF与整合酶的链合作用;AT-钩基序,易于和富含AT碱基的DNA小沟状结构结合,辅助PWWP将LEDGF链合在宿主DNA特定序列;极性氨基酸区域(Charged region,CR1-3)[22-23]。
3 LEDGF调节整合酶-DNA的相互作用
LEDGF是一个重要的慢病毒整合酶配体,可以调节慢病毒在被感染细胞中的整合反应[24]。这种细胞内源性蛋白在结构上具有双重功能:其C末端包含的整合酶结合结构域IBD,其作用是与整合酶结合,而N端部分则是将PIC链接至宿主染色体。LEDGF可以与整合酶相互作用形成复合体,从而使整合酶免于体内蛋白酶的降解[8-9,25]。LEDGF对整合酶与染色体的作用也有一定的辅助作用,可以促进病毒整合的进行。体外纯化LEDGF蛋白并以DNA为底物模拟其相互作用,可检测到LEDGF具有较强的调节链转移反应活性。实验是以一个短链DNA为底物,以一个环状质粒为靶目标,LEDGF既可有效地提高半位点(Half site)整合反应,即只能激发单一短链DNA一端的整合,又可以调节全位点(Full site)整合反应,即2个短链DNA一端整合到目标质粒上。然而,当用长链DNA替换短链DNA,LEDGF只能有效地提高半位点的整合反应。但目前还不了解为什么对于不同长度的DNA片段会有不同的整合结果。另外一个实验结果表明,当将适量的LEDGF加入经14℃,15 min预孵育的整合酶-DNA复合物中,可以有效的激发整合反应。然而,若先将LEDGF与整合酶预先孵育,再加入病毒DNA,结果半位点的整合反应显著增加[25]。
4 整合酶的四聚化以及LEDGF在其中的作用
LEDGF可以通过稳定整合酶亚单位与亚单位之间的相互作用调节整合酶的结构,并且促进整合酶的四聚化[8]。实验证明,整合酶的四聚化对于LEDGF的高亲和性结合非常重要。一个稳定的整合酶四聚体可以有效地保证催化3'端加工和链转移反应中酶的活性。当溶液中存在整合酶的单体、二聚体、四聚体以及其他的聚合形式时,IBD会强烈的促使整合酶形成四聚体,并且激发整合酶的催化功能。位点突变实验证明,赖氨酸残基14、186和188对整合酶四聚体的形成和与LEDGF的高亲和性结合上起到非常重要的作用。此外,若将赖氨酸残基14、186和精氨酸残基187突变为丙氨酸,则会明显降低整合酶在3'加工和链转移反应中的活性[25]。这些实验结果说明,四聚体的形成对于整合酶与DNA底物的相互作用,以及与LEDGF的结合都非常重要。用蛋白质印记实验分析整合酶与DNA的复合物时发现,赖氨酸残基14在与DNA结合上起到一定作用。整合酶赖氨酸残基14对于其二聚体-二聚体的作用以及整合酶与宿主DNA和LEDGF的结合都非常重要[26]。另一些实验提示,赖氨酸残基14,186和精氨酸残基187确实在病毒感染过程中起着重要作用。赖氨酸残基188突变为丙氨酸则会导致病毒的细胞感染水平下降,但并不会影响病毒的感染与传播[27]。根据这些实验结果推测,功能性四聚体的形成可以使整合酶保持四个与LEDGF高亲和性位点。这样,一个LEDGF分子就可以有效地将PIC链合到宿主染色体上。所以,整合酶四聚体是一种重要的生物学形式,在发挥其催化活性和与LEDGF的高亲和性上都起到不可或缺的作用。
5 LEDGF对整合酶活性的影响
IBD被认为是LEDGF直接与整合酶相互作用的区域。虽然IBD也可以稳定整合酶的四聚体,并且激发3'端加工反应[25],但却没有充分的证据证明该区域能够促进链转移反应的进行[28]。IBD基因敲除后的LEDGF,能竞争性抑制LEDGF依赖的HIV-1整合酶链转移反应[28],若过量表达GFP的IBD也会抑制HIV-1在人体内的整合[29]。Turlure等人[22]用裸露的DNA为靶目标进行的研究表明,LEDGF的226 aa~530 aa氨基酸残基所具有的刺激链转移反应活性约为全长LEDGF的50%。另外,激发EIAV整合酶的链转移反应需要LEDGF结构中AT-钩基序的存在。相似的实验采用重构的多聚核小体作为靶目标,结果显示在HIV-1整合到染色体DNA过程中,需要N端的PWWP结构域的参与[30]。Meehan等人[31]的实验结果证明,将IBD与异源性染色体结合会产生功能性辅助因子,能够使HIV-1在缺少LEDGF的细胞内整合反应正常进行。因此,LEDGF对慢病毒整合酶的影响包括提高生物学相关的多聚化,从而特异性的促进链反应和链合PIC到靶DNA。此外,目前还不能排除LEDGF对整合酶活性位点有变构调节功能。
6 目前研究存在的问题和展望
多项生物学实验证明,LEDGF蛋白通过其IBD结构域对整合酶催化核心区进行特异性识别,将整合酶链合到宿主染色体的特定区域,从而介导慢病毒整合酶对宿主染色体特异位点的结合。因此,LEDGF是保持慢病毒复制及感染性所必需的辅助因子。虽然LEDGF蛋白的IBD结构域与整合酶催化核心区复合物的晶体结构已被解析,但是对于LEDGF在细胞中天然特性方面的了解还是空白,甚至LEDGF在细胞染色体内在分配还未见报道。新近发现的一些其他与LEDGF结合的配体会在某些情况下利用其链合作用整合到染色体上,这将有助于进一步了解其天然的功能。
HIV的整合反应为抗逆转录病毒药物的开发提供了重要的靶点,抑制整合酶-LEDGF的相互作用被广泛认为可以用来开发新型药物。在结构生物学方面,LEDGF已经被认为是研究逆转录病毒DNA整合反应非常有用的工具。我们期望通过进一步对LEDGF的结构以及LEDGF与慢病毒整合酶共同形成的复合体结构进行解析,为整合酶抑制剂的开发提供支持。
[1]殷震,刘景华主编.动物病毒学2版.北京科学出版社,1997.
[2]Delelis O,Carayon K.Integrase and integration:Biochemical activities of HIV-1 integrase[J].Retrovirology,2008,5,114.
[3]Cai M,Zheng R,Caffrey M,et al.Solution structure of the N-terminal zinc binding domain of HIV-1 integrase[J].Nat Struct Biol,1997,4,567-577.
[4]Bushman F D,Engelman A,Palmer I,et al.Domains of the integrase protein of human immunodeficiency virus type 1 responsible for polynucleotidyl transfer and zinc binding[J].PNAS,USA 1993,90:3428-3432.
[5]Zheng R,Jenkins T M,Craigie R.Zinc folds the N-terminal domain of HIV-1 integrase,promotes multimerization,and enhances catalytic activity[J].PNAS,USA 1996,93:13659-13664.
[6]Lee S P,Xiao J,Knutson J R,et al.Zn2+promotes the self-association of human immunodeficiency virus type-1 integrasein vitro[J].Biochemistry 1997,36,173-180.
[7]Engelman A,Craigie R.Identification of conserved amino acid residues critical for human immunodeficiency virus type 1 integrase functionin vitro[J].J Virol.1992,66:6361-6369.
[8]Hare S,Di Nunzio F,Labeja A,et al.Structural basis for functional tetramerization of lentiviral integrase[J].PLoS Pathog,2009,5:e1000515.
[9]McKee C J,Kessl J J,Shkriabai N,et al.Dynamic modulation of HIV-1 integrase structure and function by cellular lens epithelium-derived growth factor(LEDGF)protein[J].J Biol.Chem,2008,283:31802-31812.
[10]Dyda F,Hickman A B,Jenkins T M,et al.Crystal structure of the catalytic domain of HIV-1 integrase:Similarity to other polynucleotidyl transferases[J].Science 1994,266:1981-1986.
[11]Fayet O,Ramond P,Polard P,et al.Functional similarities between retroviruses and the IS3 family of bacterial insertion sequences[J].Mol Microbiol,1990,4:1771-1777.
[12]Kulkosky J,Jones K S.Residues critical for retroviral integrative recombination in a region that is highly conserved among retroviral/retrotransposon integrases and bacterial insertion sequence transposases[J].Mol Cell Biol,1992,12:2331-2338.
[13]Hazuda D J,Wolfe A L.Viral long terminal repeat substrate binding characteristics of the human immunodeficiency virus type 1 integrase[J].J Biol Chem,1994,269:3999-4004.
[14]Goldgur Y,Dyda F,Hickman A B,et al.Three new structures of the core domain of HIV-1 integrase:An active site that binds magnesium[J].PNAS,USA 1998,95:9150-9154.
[15]Eijkelenboom A P,Lutzke R A,Boelens R,et al.The DNA binding domain of HIV-1 integrase has an SH3-like fold[J].Nat Struct Biol,1995,2:807-810.
[16]Robinson H,Gao Y G,McCrary B S,et al.The hyperthermophile chromosomal protein Sac7d sharply kinks DNA[J].Nature,1998,392:202-205.
[17]Krueger J K,McCrary B S,Wang A H,et al.The solution structure of the Sac7d/DNA complex:A small-angle X-ray scattering study[J].Biochemistry 1999,38:10247-10255.
[18]Lutzke R A,Plasterk R H.Structure-based mutational analysis of the C-terminal DNA-binding domain of human immunodeficiency virus type 1 integrase:Critical residues for protein oligomerization and DNA binding[J].J Virol,1998,72:4841-4848.
[19]Richman D D.HIV chemotherapy[J].Nature,2001,410(6831):995-1001.
[20]Cherepanov P,Maertens G,Proost P,et al.HIV-1 integrase forms stable tetramers and associates with LEDGF/p75 protein in human cells[J].J Biol Chem,2003,78:372-381.
[21]Vanegas M,Llano M.Identification of the LEDGF/p75 HIV-1 integrase-interaction domain and NLS reveals NLS-independent chromatin tethering[J].J Cell Sci,2005,118:1733-1743.
[22]Turlure F,Maertens G,Rahman S,et al.A tripartite DNA binding element,comprised of the nuclear localization signal and two AT-hook motifs,mediates the association of LEDGF/p75 with chromatinin vivo[J].Nucleic Acids Res,2006,34:1663-1675.
[23]Llano M,Vanegas M,Hutchins N,et al.Identification and characterization of the chromatin-binding domains of the HIV-1 integrase interactor LEDGF/p75[J].J Mol Biol,2006,360:760-773.
[24]Engelman A,Cherepanov P.The lentiviral integrase binding protein LEDGF/p75 and HIV-1 replication[J].PLoS Pathog,2008,4:e1000046.
[25]Pandey K K,Sinha S.Transcriptional coactivator LEDGF/p75 modulates human immunodeficiency virus type 1 integrase-mediated concerted integration[J].J Virol,2007,81:3969-3979.
[26]Zhao Z,McKee C J,Kessl J J,et al.Subunit-specific protein footprinting reveals significant structural rearrangements and a role for N-terminal Lys-14 of HIV-1 integrase during viral DNA binding[J].J Biol.Chem,2008,283:5632-5641.
[27]Lu R,Limon A,Devroe E,et al.classⅡintegrase mutants with changes in putative nuclear localization signals are primarily blocked at a postnuclear entry step of human immunodeficiency virus type 1 replication[J].J Virol,2004,78:12735-12746.
[28]Cherepanov P,Devroe E.Identification of an evolutionarily conserved domain in human lens epithelium-derived growth factor/transcriptionalco-activatorp75 (LEDGF/p75)thatbinds HIV-1 integrase[J].J Biol Chem.2004,279:48883-48892.
[29]Llano M,Saenz D T,Meehan A,et al.An essential role for LEDGF/p75 in HIV integration[J].Science,2006,314:461-464.
[30]Botbol Y,Raghavendra N K,Rahman S,et al.Chromatinized templates reveal the requirement for the LEDGF/p75 PWWP domain during HIV-1 integrationin vitro[J].Nucleic Acids Res.2008,36:1237-1246.
[31]Meehan A M,Saenz D T,Morrison J H,et al.LEDGF/p75 proteins with alternative chromatin tethers are functional HIV-1 cofactors[J].PLoS Pathog,2009,5:e1000522.
[32]Gao K,Wong S,Bushman F.Metal binding by the D,DX35E motif of human immunodeficiency virus type 1 integrase:selective rescue of Cys substitutions by Mn2+in vitro[J].J Virol,2004,78:6715-6722.
S852.65
B
1008-0589(2010)03-0241-04
(本文编辑:赵晓岩)
猜你喜欢
生物化学与生物物理进展(2022年6期)2022-07-21 11:52:06
煤炭与化工(2021年6期)2021-08-06 10:04:10
中学生数理化(高中版.高考理化)(2021年2期)2021-03-19 08:52:38
科学(2020年3期)2020-11-26 08:18:22
当代水产(2020年3期)2020-06-15 12:03:02
安徽大学学报(自然科学版)(2016年2期)2016-09-20 12:09:27
池州学院学报(2015年3期)2016-01-05 01:13:04
实用皮肤病学杂志(2015年4期)2015-12-22 11:21:42
天津科技大学学报(2015年2期)2015-08-09 01:40:42
医学研究杂志(2015年12期)2015-06-10 06:57:46
