
流行性乙型脑炎病毒E蛋白的截短表达与间接ELISA诊断方法的建立
2010-09-10 04:24:28祖立闯王金良沈志强
中国预防兽医学报 2010年3期
祖立闯,王金良,管 宇,沈志强,*,董 林,李 娇
(1.山东绿都生物科技有限公司,山东滨州 256600;2.山东省滨州畜牧兽医研究院,山东滨州 256600)
流行性乙型脑炎病毒E蛋白的截短表达与间接ELISA诊断方法的建立
祖立闯1,王金良2,管 宇2,沈志强1,2*,董 林2,李 娇1
(1.山东绿都生物科技有限公司,山东滨州 256600;2.山东省滨州畜牧兽医研究院,山东滨州 256600)
为建立一种快速检测流行性乙型脑炎病毒抗体的方法,本研究参照已发表的JEV基因组序列,应用RT-PCR扩增了长约1 000 bp的E基因片段,连接pET30a表达载体中,经诱导后获得了以包涵体形式表达的重组E蛋白。重组蛋白纯化后,经免疫印迹检测证明其具有良好的抗原性和特异性。以该蛋白作为诊断抗原,建立了检测流行性乙型脑炎病毒抗体的E-ELISA诊断方法。该诊断方法具有良好的敏感性、特异性和重复性,为JEV的快速诊断、免疫猪群抗体监测和JEV流行病学调查提供了一种快速、简便的血清学诊断方法。
乙型脑炎病毒;E蛋白;截短表达;间接ELISA
流行性乙型脑炎是由日本乙型脑炎病毒(Japanese encephalitis virus,JEV)引起的以中枢神经系统发生病变的急性传染病,也是一种人畜共患的自然疫源性疾病。临床上以高热、意识障碍、惊厥、强直性痉挛和脑膜刺激等神经症状为主要特征[1]。该病以蚊虫为媒介传播,对人类危害巨大,是人类中枢神经系统最常见的虫媒病之一[1]。我国是乙脑发病人数最多的国家,占世界总发病数的80%以上,病死率10%以上,幸存者中约有15.3%留有不同程度的后遗症[2]。在动物中,猪是本病的中间宿主,带毒猪是本病的主要传染源,它与蚊形成了蚊、猪循环传播模式,并对病毒起放大器作用,可引起妊娠母猪流产、死胎;公猪发生睾丸炎;仔猪因脑炎死亡,给养猪业造成了巨大的经济损失[1,3]。
JEV属于黄病毒科黄病毒属,单股正链RNA病毒,基因组长11 kb(10 976 bp),整个基因组只有一个开放阅读框架,编码一个全长3 432个氨基酸的多蛋白前体[4]。该多蛋白前体经蛋白酶切割加工,产生3个结构蛋白:核衣壳蛋白(C)、膜蛋白(PrM/M)、囊膜糖蛋白(E);7个非结构蛋白[4-5]。E基因大小为1 500 bp,编码500个氨基酸,E蛋白具有神经毒性和神经侵袭性的致病位点,作为膜融合蛋白促使病毒进入神经细胞[6]。E蛋白还是JEV病毒粒子表面最主要的结构蛋白,其表面的抗原决定簇具有血凝活性和中和活性,能刺激机体产生血凝抑制抗体、补体结合抗体、中和抗体,引起宿主产生保护性的免疫应答,从而保护机体免受病毒攻击[7-9]。本研究对E基因进行了截短表达,并建立了间接ELISA诊断方法,为流行性乙型脑炎诊断试剂盒的研制提供了有力的技术支持。
1 材料和方法
1.1 病毒株、菌株、细胞与血清 JEV毒株DH5α、 BL21(DE3)、 pET30a(+)、 仓 鼠 肾 细 胞BHK-21均由本实验室保存。由本实验分离并保存。猪圆环病毒2型(PCV-2)、猪伪狂犬病病毒(PRV)、猪繁殖与呼吸综合征病毒(PRRSV)、猪细小病毒(PPV)、猪流行性腹泻病毒(PEDV)、猪传染性胃肠炎病毒(TGEV)、猪瘟病毒(CSFV)阳性血清及JEV参考阴阳性血清均购自深圳康百得生物科技有限公司,其它血清样品由本实验室保存。
1.2 主要试剂 M-MLV反转录酶、Ribonuclease Inhibitor、ExTaq酶、pMD18-T、T4 DNA连接酶及其它限制性内切酶均购自宝生物工程(大连)有限公司;兔抗猪IgG辣根过氧化物酶(HRP)标记抗体、低分子量蛋白Marker均购自北京鼎国生物技术有限公司;TRIzol、质粒DNA提取试剂盒与胶回收试剂盒购自上海生工生物工程技术服务有限公司。
1.3 引物设计与合成 根据GenBank登录的JEV基因组序列(AF315119.1),利用oligo6.2设计扩增E基因的一对引物,P1 5'-GCGAATTCATCCTCCTGCTGTTGGTC-3';P25'-GCGAAGCTTCGCAACGGAAACAATCG-3',引物由上海生物工程公司合成。
1.4 病毒基因组RNA的提取 将JEV接种于BHK-21单层细胞,待细胞出现典型的病变后,收获病毒,按TRIzol试剂说明提取病毒基因组RNA。
1.5 E基因的RT-PCR扩增与克隆 以提取的病毒基因组RNA为模板,反转录合成cDNA。以cDNA为模板进行PCR反应,反应条件为:94℃5 min、94℃l min,56℃l min,72℃ l min,共进行30个循环;72℃10 min,4℃终止反应。PCR产物纯化后与pMD18-T载体于4℃过夜连接,然后转化DH5α大肠杆菌感受态细胞,PCR及EcoRⅠ/HindⅢ双酶切鉴定得到阳性重组质粒pMD18-T-E,由上海生物工程公司进行序列测定。
1.6 E基因表达载体的构建与鉴定 利用EcoRⅠ/HindⅢ双酶切pMD18-E,回收目的片段,将其定向克隆到经EcoRⅠ/HindⅢ双酶切的pET30a表达载体中,转化感受态细菌DH5α,经PCR及酶切鉴定后获得重组表达载体pET-E。鉴定阳性克隆送上海生物工程有限公司测序,以验证其阅读框架。
1.7 pET-E重组蛋白的表达与纯化 将pET-E重组质粒转化BL21(DE3)感受态细胞,采用不同诱导剂浓度、不同时间、不同温度诱导pET-E重组蛋白的表达。经SDS-PAGE电泳分析,确定最佳诱导剂浓度、诱导时间、诱导温度。以最佳条件诱导表达后,超声破碎细胞,12 000 r/min离心15 min,分别收集上清及沉淀进行SDS-PAGE分析,鉴定目的蛋白是以包涵体形式存在于沉淀中,用尿素对其进行变性处理,包涵体溶解后采用镍离子亲和层析法纯化,纯化蛋白透析复性,进行SDS-PAGE电泳分析。
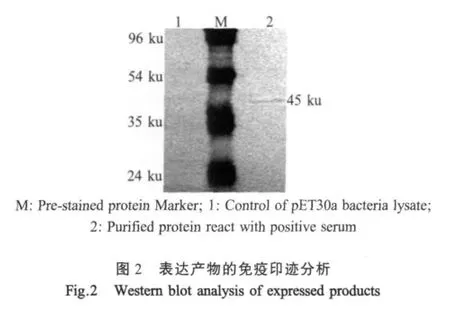
1.8 E重组蛋白的western blot活性检测 纯化的重组蛋白经SDS-PAGE后,转印至硝酸纤维素膜,5%脱脂奶粉4℃封闭过夜,加JEV阳性血清(1∶100稀释)37℃作用1 h,TBST缓冲液洗3次,加入兔抗猪IgG辣根过氧化物酶(HRP)标记抗体(1∶5 000稀释)37℃作用50 min,TBST缓冲液洗3次,在二氨基联苯胺(DAB)缓冲溶液中显色10 min。
1.9 间接ELISA诊断方法的建立
1.9.1 JEV E-ELISA反应条件的优化试验在相同的反应条件下,分别将重组抗原以不同包被浓度与待检样品的不同稀释倍数、不同封闭液与不同封闭时间、二抗不同稀释倍数与不同作用时间组成方阵,经方阵滴定试验分别进行各组份的条件优化。原则是阳性血清OD450nm≈1.0,阴性血清OD450nm<0.2,选取P/N值最大的各个组合。在相同反应条件下,与待检样品分别作用30 min~120 min,底物分别显色5 min~20 min,选择阳性血清OD450nm≈1.0,阴性血清OD450nm<0.2,P/N值最大的一抗、底物作用时间。
1.9.2 JEV E-ELISA判断标准的确定试验判定标准采用Cut-Off值法,血凝抑制试验(HI)检测为阴性的38份血清样本,利用JEV E-ELISA已优化的最佳反应条件检测,将OD450nm值转换成S/P值,计算出38份血清的S/P平均值(χ)和标准差(SD),我们规定 S/P(样本)≥χ+3SD时为阳性,S/P(样本)<χ+2SD时为阴性,当χ+2SD≤S/P(样本)<χ+3SD为可疑,需重新检测样本,重检时S/P值≥χ+2SD时判为阳性,S/P值<χ+2SD时为阴性。
1.9.3 特异性试验以建立的判断标准与PCV-2、PRV、PRRSV、PPV、PEDV、TGEV、CSFV阳性血清进行间接JEV E-ELISA检测,同时设JEV标准阳、阴性血清对照,确定重组抗原与其它常见猪病阳性血清是否发生交叉反应。

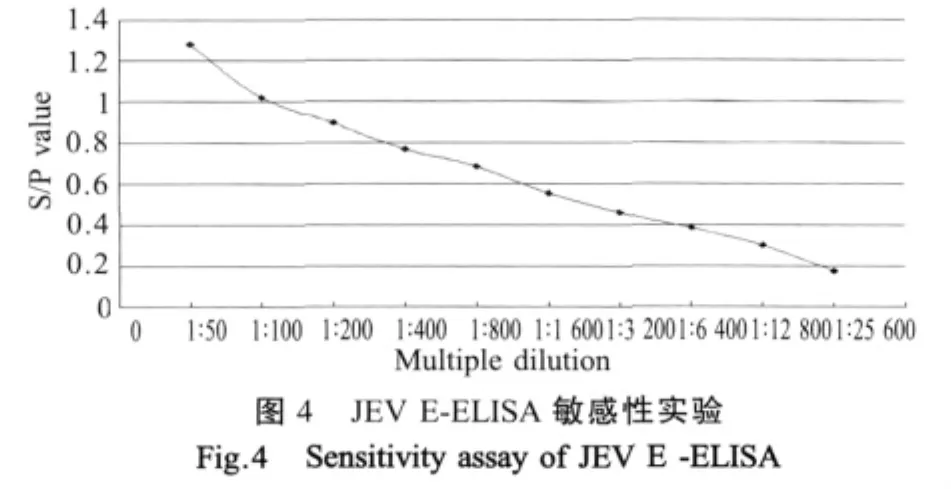
1.9.4 敏感性试验将JEV标准阳性对照血清按1∶50~1∶25 600倍稀释,每个稀释度平行做4个重复,用建立的JEV E-ELISA检测,测定OD450nm值,将其换算成S/P值,确定其抗体检出效价。
1.9.5 重复性试验(1)批内重复性试验,选取8份JEV抗体水平不同的血清,在相同试验条件下,每份血清样本平行做8个重复,用JEV E-ELISA检测,对检测结果进行统计学分析。(2)批间重复性试验,选取8份JEV抗体水平不同的血清,在相同试验条件下,在4个不同时间用JEV E-ELISA检测,对检测结果进行统计学分析;在相同试验条件下,用4批不同时间纯化的重组抗原进行JEV E-ELISA检测,对检测结果进行统计学分析。
1.9.6 符合性试验血凝抑制试验(HI)检测结果为阳性和阴性的血清样本共156份,再用JEV E-ELISA检测,以HI检测结果为标准,计算E-ELISA相对于HI的符合率、敏感性、特异性。
1.10 现地应用试验 应用本实验建立的JEV EELISA检测了来自山东、河南、河北等地的658份血清样本,确定JEV血清抗体阳性率。
2 结果
2.1 PCR扩增、克隆及重组表达质粒的鉴定PCR扩增产物经过1%琼脂糖凝胶电泳,在位于约1 000 bp处可见特异性DNA扩增条带,与预期大小相符。构建的pMD-E质粒经PCR及EcoRⅠ/HindⅢ双酶切鉴定后,测序结果表明克隆序列与参考毒株完全相同。构建的pET-E重组质粒经PCR及EcoRⅠ/HindⅢ双酶切鉴定后,测序结果表明其阅读框正确。
2.2 E重组蛋白的表达及纯化 以最佳条件(诱导温度为37℃、诱导剂浓度为1.0 mmol/L、诱导时间为5 h)诱导表达,SDS-PAGE表明所表达蛋白的分子量为45 ku,薄层扫描分析表明目的蛋白占菌体总蛋白的33.8%。目的蛋白大部分是以包涵体形式表达的,包涵体经8 M尿素变性后,采用镍离子亲和层析法纯化,纯化后的蛋白透析复性。薄层扫描分析表明复性后蛋白的纯度高达89.5%,Bradford法测定蛋白浓度为0.216 mg/mL(图1)。

2.3 E重组蛋白的活性鉴定 纯化后蛋白经SDS-PAGE电泳后,转印到硝酸纤维素膜上,与JEV阳性血清作用(图2)。表明该重组蛋白能与JEV阳性血清发生特异性反应,而空载体表达菌与JEV阳性血清没有反应。
2.4 JEV E-ELISA相关条件确定试验结果
2.4.1 JEV E-ELISA最佳反应条件的确定重组抗原最佳包被浓度为5.4 μg/mL;最佳封闭液为含1%BSA的PBST,最佳封闭时间为37℃作用1.5 h;血清样本最佳稀释倍数为1∶100,最佳作用时间为37℃作用1 h;二抗最佳工作浓度为1∶5 000,最佳作用时间为37℃作用1 h;底物最佳显色时间为15 min。
2.4.2 JEV E-ELISA判定标准的确定38份阴性血清,经JEV E-ELISA检测后,将血清样本分成5组,以S/P值为横坐标,以各组样品数量为纵坐标,得到JEV阴性的血清S/P数值分布直方图(图3),这些样本的S/P值主要分布在0.05~0.20之间,占血清样本的89.5%,数据分布合理,可以作为制定判定标准的依据。计算得出38份阴性血清S/P,当S/P(样本)≥0.294时判为阳性;S/P(样本)<0.239时判为阴性;0.239≤S/P(样本)<0.294时判为可疑。对可疑样本须重新检测,重检时,S/P(样本)≥0.239为阳性,S/P(样本)<0.239为阴性。

2.5 特异性试验结果 重组抗原与常见的PCV-2、PRV、PRRSV、PPV、PEDV、TGEV、CSFV 7种猪病阳性血清反应的S/P值均低于0.239,呈阴性反应。该结果表明重组蛋白与常见7种猪病的阳性血清不发生交叉反应,显示出了很好的特异性。
2.6 敏感性试验结果 检测JEV标准阳性对照血清,当血清稀释至12 800倍时,检测结果仍为阳性(图4),证明所建立的方法具有较好的敏感性。
2.7 重复性试验结果 批内重复性试验的变异系数小于5%(表1),不同时间、不同批次重组抗原的批间重复性试验变异系数均小于10%(表2、表3)。说明JEV E-ELISA具有良好的重复性。

表1 间接ELISA批内重复结果Table 1 Intro-batch repeatability test of the indirect ELISA

表2 不同时间批间重复性试验结果Table 2 Different time of inter-batch repetition test

表3 不同批次重组抗原批间重复性试验结果Table 3 Different batches recombinant antigens of inter-batch repetition test
2.8 比较试验结果 HI检测结果为阳性的121份血清样本,JEV E-ELISA检出阳性102份,阴性19份;HI检测结果为阴性的45份血清样本,JEV E-ELISA检出阴性36份,阳性9份。JEV E-ELISA相对于HI的符合率为83.1%,敏感性为84.3%,特异性为80.0%(表4)。
2.9 现地应用试验结果 应用本实验建立的JEVE-ELISA检测了来自山东、河南等地的658份血清样本,检出阳性448份,血清抗体阳性率为68.1%。

表4 JEV E-ELISA与HI符合率试验结果Table 4 Agreement test between JEV E-ELISA and HI
3 讨论
目前,黄病毒的疫苗大多数是针对E蛋白的抗原表位。Chen等研究认为,E蛋白是唯一能诱发机体产生抗JEV中和抗体的重要抗原[10]。Kolaskar等的研究表明,E蛋白含有3个抗原域:域Ⅰ,也称为中心域,由E1位~E51位、E137位~E196位和E293位~E311位的130个氨基酸残基组成,此域有糖基化位点和具有血清学及生物学活性的抗原表位;域Ⅱ,由E52位~E136位和E197位~E292位的181个氨基酸残基组成,具有中和活性和血凝活性的抗原表位;域Ⅲ,由E312位~E411位的100个氨基酸残基组成,参与受体结合过程,在黄病毒属中十分保守[11]。Chia等把截短的JEV E蛋白基因(不含跨膜区)分为“A”、“B”两段在大肠杆菌中表达,发现下游片段(B)表达产物的抗原性和免疫原性优于上游片段(A)[12]。本研究通过DNAStar生物学软件分析目的蛋白的生物学特性,截取抗原性较好的E1位~E343位的氨基酸残基,其包含了完整的域Ⅰ、域Ⅱ及“A”、“B”片段,所以表达的蛋白具有良好的抗原性。
本实验选择pET30a原核表达载体,表达了JEV的E蛋白,经免疫印迹试验证实所截短表达的E蛋白可与JEV阳性血清发生较强的反应,表明重组E蛋白具有良好的反应性与特异性。
目前在JEV的流行病学调查中,常用HI试验、补体结合试验、中和试验等检测方法,但常规的血凝抑制试验、补体结合试验、中和试验等血清学诊断方法费时费力,特异性和敏感性不高,ELISA诊断方法具有敏感性和特异性强、操作简单、检测快速、重复性好、高通量、成本低等特点,是当前动物传染病检疫、流行病学调查和免疫监测广泛采用的血清学诊断技术[13]。由于JEV的全病毒抗原制备过程繁琐、存在潜在的危险性及缺少标准化,因此,利用基因工程表达的重组抗原已逐步得到重视。同时,这种重组抗原只能与该病毒特定成份所诱生的抗体发生反应,因而可用于亚单位或标记疫苗免疫动物与自然感染动物的鉴别诊断。本实验所表达的重组E蛋白具有良好的反应活性,作为包被抗原建立的间接ELISA诊断方法具有很好的敏感性和特异性,为我国JEV的诊断、疫苗免疫水平监测、流行病学调查等提供一种简便、快速的血清学检测方法。
[1]Konishi E,Pincus S,Paoletti E,et al.Avipox virus-vectored Japanese encephalitis virus vaccines:use as vaccine candidates in combination with purified sub unit immunogens[J].Vaccine,1994,12(7):633-638.
[2]陈焕春.规模化养猪场疫病控制与净化[M].北京:中国农业出版社,2000,187.
[3]Wang J W,Fu S H,Wang H Y,et al.Isolation and identification of Japanese encephalitis virus in Liaoning province[J].中华实验和临床病毒学杂志,2006,20(1):61-65.
[4]McMinn P C.The molecular basis of virulence of the encephalitogenic flaviviruses[J].J Gen Virol,1997,78(11):2711-2722.
[5]Chen W R,Tesh R B,Rico-Hesse R.Genetic variation of Japanese encephalitis virus in nature[J].J Gen Virol,1990,71(12):2915-2922.
[6]Mandl C W,Guirakhoo F,Holzmann H,et al.Antigenic structure of the flavivirus envelope protein E at the molecular level,using tick-borne encephalitis virus as a model[J].J Virol,1989,63(2):564-571.
[7]Takegami T,Miyamoto H,Nakamura H,et al.Biological activities of the structural proteins of Japanese encephalitis virus[J].Acta Virol,1982,26(5):312-320.
[8]Li P,Zheng Q S,Wang Q,et al.Immune responses of recombinant adenoviruses expressing immunodominant epitopes against Japanese encephalitis virus[J].Vaccine,2008,26(46):5802-5807.
[9]Li Y,Ye J,Cao S,et al.Immunization with pseudotype baculovirus expressing envelope protein of Japanese encephalitis virus elicits protective immunity in mice[J].J Gene Med,2009,11(2):150-159.
[10]Chen H W,Pan C H,Liau M Y,et al.Screening of protective antigens of Japanese encephalitis virus by DNA immunization:a comparative study with conventional viral vaccines[J].J Virol,1999,73(12):10127-10145.
[11]Kolaskar A S,Kulkarni Kale U.Prediction of three-dimensional structure and mapping of conformational epitopes of envelope glycoprotein of Japanese encephalitis virus[J].Virology,1999,261(1):31-42.
[12]Chia S C,Leung P S,Liao C P,et al.Fragment of Japanese encephalitis virus envelope protein produced inE.coliprotects mice from virus challenge[J].Microb Pathog,2001,31(1):9-19.
[13]Konishi E,Mason P W,Shope R E.Enzyme-linked immunosorbent assay using recombinant antigens for serodiagnosis of Japanese encephalitis[J].J Med Virol,1996,48(1):76-79.
Development of an indirect ELISA for detecting antibodies to japanese encephalitis virus using recombinant truncated glycoprotein E expressed inE.coli
ZU Li-chuang1,WANG Jin-liang2,GUAN Yu2,SHEN Zhi-qiang1,2*,DONG Lin2,LI Jiao1
(1.Shandong Lvdu Bio-Sciences and Technology Co.Ltd.,Binzhou 256600,China;2.Shandong Binzhou Animal Science and Veterinary Medicine Academy,Binzhou 256600,China)
In this study,DNA fragment encoding the Japanese encephalitis virus(JEV)E protein was amplified by RT-PCR,cloned and expressed inE.coli.An indirect ELISA was successfully developed to detect anti-JEV antibody using the purified recombinant E protein as coating antigen.The assay was specific,sensitive and reproducible,which could be used as a rapid serology detection method for monitoring of anti-JEV antibody and epidemiologic survey of JEV.
japanese encephalitis virus;E protein;prokaryotic expression;indirect ELISA;diagnosis
S852.659.6
A
1008-0589(2010)03-0200-05
*Correspondingauthor
2009-07-21
祖立闯(1981-),男,黑龙江宾县人,硕士,从事动物病毒学研究.
*通信作者:E-mail:bzshenzq@vip.sina.com
(本文编辑:彭永刚)
猜你喜欢
家庭医学(下半月)(2019年10期)2019-11-16 08:59:52
中国医学影像学杂志(2018年9期)2018-10-17 01:26:58
英美文学研究论丛(2018年2期)2018-08-27 01:56:30
中国医药指南(2017年3期)2017-11-13 02:55:17
老年医学与保健(2017年6期)2017-02-06 05:29:54
西南医科大学学报(2015年1期)2015-08-22 13:01:46
医学研究杂志(2015年6期)2015-07-01 17:41:11
医学研究杂志(2015年12期)2015-06-10 06:57:46
癌变·畸变·突变(2015年3期)2015-02-27 06:15:07
中国药理学通报(2014年2期)2014-05-09 08:22:30
