
犬蝠取食对叶榕果实的行为和相互适应关系研究
2010-09-08 06:38胡亚明唐占辉丁雪梅宋传涛马逊风
东北师大学报(自然科学版) 2010年3期
胡亚明,唐占辉,丁雪梅,宋传涛,曹 敏,马逊风
(1.东北师范大学城市与环境科学学院,吉林长春130024; 2.中国科学院西双版纳热带植物园,云南勐腊666303; 3.吉林大学畜牧兽医学院,吉林长春130062)
犬蝠取食对叶榕果实的行为和相互适应关系研究
胡亚明1,2,唐占辉1,丁雪梅3,宋传涛1,曹 敏2,马逊风1
(1.东北师范大学城市与环境科学学院,吉林长春130024; 2.中国科学院西双版纳热带植物园,云南勐腊666303; 3.吉林大学畜牧兽医学院,吉林长春130062)
在西双版纳地区,观察和测量了对叶榕(Ficus hispida)的结果特点、果实特征以及取食该果实的犬蝠(Cynopterus sphinx)的形态特征,并通过飞行笼实验考察了对叶榕果实提供的视觉和嗅觉信息及犬蝠取食时视觉和嗅觉的应用情况.结果表明:犬蝠利用视觉系统能成功地分辨出完整和破碎的果实,每小时访问完整果实的次数为(15.5±1.61)次,访问浆状果实的次数为(10.5±1.45)次;在访问有果实和没有果实的盒子时,每小时访问前者的次数为(37.4±7.62)次,显著高于后者(1.1±0.88)次.对叶榕果实的生物和生态学特征与犬蝠的行为学特征建立起了较好的相互适应性.
对叶榕;果蝠;飞行笼实验;西双版纳
植物果实的特征通常可以影响取食者(食果动物)的食物选择[1-2];果实特征包括果实的大小、颜色、营养成分和种子大小等[3-16].同时,食果动物作为种子传播者,生态习性和行为上的不同也会导致所取食果实特征相应的进化改变[17-19].在云南地区热带森林生态系统中,榕树是一类关键性树种[20-21],榕树的果实可以作为一些动物的食物,如鸟类、蝙蝠等[22-24].对西双版纳地区犬蝠(Cynopterus sphinx)食性的调查发现,榕果是它们食物中重要的组成部分,其中热带雨林常见的先锋树种——对叶榕(Ficus hispida)更是一种长期且稳定的食物资源[25].
关于果实特征和其取食者的关系,Silvia等人从果实的大小和颜色这两个角度分析了鸟类和哺乳动物各自所取食的果实的特点[26];也有学者研究了新大陆和旧大陆热带地区的果蝠所取食的榕果的一些特征[18].在中国西双版纳热带雨林也有学者对果实特征的几种模式进行了调查[27].本文以对叶榕为例,研究了它和果实取食者和种子传播者——犬蝠之间的相互适应关系.果实方面主要考察了挂果特点、大小、成熟颜色、气味、成熟的同步性和种子特征等方面,同时测量犬蝠在形态方面的一些适应特征,研究了果蝠在取食对叶榕果实时感官系统的应用,结合对叶榕果实营养成分的已知数据,探讨了对叶榕与犬蝠之间的相互适应关系.
1 材料和方法
1.1 研究地点
实验于2009年8-11月进行,研究地点在中国科学院西双版纳热带植物园内(N 21°08′、E 99°56′,海拔560 m),该园占地900 hm2.研究地点年平均气温21.17℃,年平均降雨量1500~1600 mm,年平均相对湿度86%[28].该地区干湿季变化明显,雨季为5-10月,旱季为11月至翌年4月[29].
1.2 研究对象
对叶榕属榕属(Ficus)植物,雌雄异株,全年结果,在中国海南、云南和贵州等地分布[30].成熟时雌果和雄果外观上几乎没有差别,但雌果蜂孔不明显,里面多是干净的种子;雄果则蜂孔明显,里面有大量榕小蜂的存在.实验喂养犬蝠时发现,它们取食的大部分是雌果,因此实验重点以对叶榕雌果为研究对象.
犬蝠体型中等,嗅觉系统较发达[31],其食物主要是植物果实和叶子[25],同时也取食爪哇木棉(Ceiba pentandra)等植物的花蜜[32-34].
1.3 研究方法
野外调查对叶榕的挂果方式、果实颜色、果实气味.我们在果实成熟前标记了1棵雄树的37个果实和1棵雌树东南西北4个方向的果实(分别为37,47,32和35个)以考察树间和树内的成熟同步性,研究对叶榕果实的成熟过程.从8月18日起,每天17:00观察并记录成熟果实个数,直至所有果实成熟.对叶榕果实特征的测量是从13棵树上采集果实,室内测量成熟果实的鲜重、干重,长轴、短轴长度,果肉湿重、果肉干重及种子数.
测量犬蝠体重、翼长等指标.在飞行笼中研究对叶榕果实和犬蝠相应感官应用的互利关系.
飞行笼(6 m×3 m×3 m)实验设置如下:
(1)研究果实形状和犬蝠的视觉关系:称取两份等量且足量的雌性对叶榕果实,一份为完整果实,一份将果实捣碎为浆状(看不出果实形状),分别放在笼内两个食物盘中(见图1).对犬蝠在笼内的取食行为进行观察记录,时间从20:00~24:00,每次观察结束后即放飞.本实验每天观察7只犬蝠,连续观察2 d.观察记录犬蝠每小时对每个果盘的访问次数,这里的“访问”是指吃到食物或徘徊、停留在果盘和果盒上至少1 s.为了不打扰犬蝠的访问行为,观察者在飞行笼外1.5 m左右借助月光和夜视仪观察.同时,为了消除犬蝠记忆行为的影响,每隔1 h调换两个果盘的位置,直到实验结束.
(2)研究果实气味和犬蝠嗅觉的关系:制作两个外形一样且封闭的纸盒,在每个纸盒前后左右4个面分别打上众多细孔(直径<2 mm,打孔部位在食物放置部位以上),以便于果实气味的释放,同时使犬蝠访问时得不到食物回报.在其中一个盒里放有果实,另一个不放果实(见图2).观察记录方式同(1).本实验每天观察4只犬蝠个体,时间从20:00~24:00,连续观察2 d.
在野外,以所研究的母树为中心,根据残留果渣来判断和搜寻犬蝠的进食地,确定对叶榕种子的传播距离.

图1 飞行笼中犬蝠取食对叶榕的视觉实验

图2 飞行笼中犬蝠取食对叶榕的嗅觉实验
1.4 数据分析与处理
数据分析利用 SPSS 13.0软件完成,飞行笼中视觉和嗅觉数据的正态分布性用单样本Kolmogorov-Smirnov来确定,经检验每组数据服从正态分布(P>0.05),在分析对叶榕雌雄果大小差异时利用独立样本t检验完成.同样在视觉和嗅觉实验中,完整果实和浆状果实访问次数的差异、装有果实和没有果实盒子的访问次数差异也用独立样本t检验完成.显著水平设置为 a=0.05.对叶榕雌树和雄树成熟曲线图利用SigmaPlot 8.0完成.
2 实验结果
2.1 对叶榕果实及犬蝠形态特征
实验期间,我们观察到在对叶榕的主要枝干上,大量果实成串状挂在上面,果实周围很少有树叶遮挡,每串果实数量不等,少则几个,多至几十个,果实暴露度很高.成熟时果实为黄色,具有浓郁的果香.雄果个体要大于雌果(df=138,t=8.640,p=0.00<0.05).成熟雌果含有很多小种子,数量可达到上千个(见表1).对叶榕果实及犬蝠的形态特征见表1、表2.雌性对叶榕果实鲜重与雌、雄犬蝠体重比分别为39.7%和39.4%,雄性对叶榕果实鲜重与雌、雄犬蝠体重比分别为53.2%和51.9%.

表1 对叶榕的果实特征
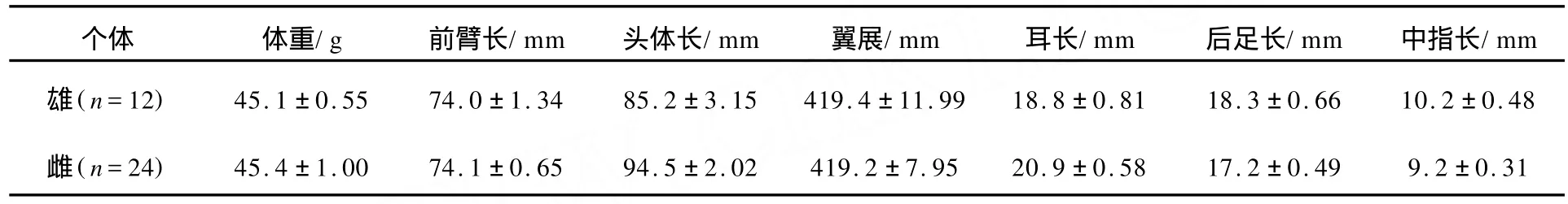
表2 犬蝠的形态特征
2.2 对叶榕果实成熟曲线
对叶榕果实成熟过程较平稳,成熟高峰期不明显,对叶榕雌树标记果实在树内4个方向上成熟比较同步,以32个果实(树西标记果实数,数目最少)为例,东南西北方向成熟天数分别为12,18,18和12 d,均在相差不到1个星期之内成熟.实验中所选的雄树标记果实从8月22日开始成熟,而标记的雌果则从9月4日开始成熟(见图3、图4).实验期间也观察到其他对叶榕树之间也存在这种现象,有的果实成熟期均已结束,有的还处于幼果期,树间果实成熟表现出较明显的不同步性.

图3 对叶榕雌果在四个方向上的成熟曲线
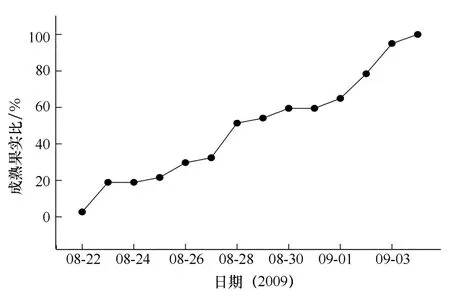
图4 对叶榕雄果的成熟曲线
2.3 犬蝠取食对叶榕的行为
在飞行笼实验中观察到,犬蝠在获取整个对叶榕果实后,快速飞到其他地点去进食,由于果实个体和重量相对较大,所以采取的是身体正立,用两前指勾住可悬挂物,负载身体及果实重量,仅用嘴部操作和进食果实(见图5).在进食过程中,犬蝠嚼食对叶榕果实,一部分果汁和果肉进入消化道,一些粒径细小的种子也混杂其中,一段时间后随粪便排出;而一部分果肉不进入消化道,嚼成团状果渣后,被犬蝠吐落在进食地及其周围,而这些果渣中也往往包含一部分种子.
在视觉实验中,每小时访问装有完整果实的果盘次数为15.5±1.61次(n=8),浆状果实为10.5± 1.45次(n=8),前者明显高于后者(df=14,t=2.303,p=0.037<0.05),说明在嗅觉信号相同的情况下,对叶榕果实所展示的特征可以使犬蝠在视觉上分辨出来完整个体.
犬蝠嗅觉实验中,每小时访问装有果实的盒子次数为37.4±7.62次(n=8),空盒子的次数为1.1± 0.88次(n=8),两者差异明显(df=14,t=4.726,p=0.00<0.05),说明在视觉信号相同的情况下,对叶榕果实的气味可以使犬蝠利用嗅觉找到食物位置,而且准确率相对更高.
飞行笼实验表明,犬蝠在取食时同时运用了视觉和嗅觉系统,对叶榕果实的形状和气味都是有效的信号.
2.4 种子传播范围
实验期间我们在母树周围500 m范围内搜寻,分别在正南、东南和西南方向发现3处进食地,树下发现大量对叶榕果渣,距离为154,197和359 m.传播生境分别为森林边缘、森林边缘和林地,而在进食地周围处于果实成熟期的只有实验样树,因此我们判定犬蝠携带母树的对叶榕果实到此进食.
3 讨论
对叶榕在主要枝干上成串挂果,这样使果实相对周围环境来说显得很突出,因此我们认为这对于在夜间觅食的犬蝠等果蝠来说,增加了它们视觉上的方便性,减轻了寻找食物所带来的压力.对叶榕果实成熟时为淡黄色且个体较大,果香浓郁,这对一些食果鸟类来说,可能会降低它们的取食机会,因为鸟类一般对鲜艳的颜色较敏感且嗅觉不发达,同时又受限于喙张开的角度,因而不会选择进食一些大的果实[35-37].有学者认为依靠鸟类传播种子的果实,一般颜色鲜艳,如红色、黑色等,并且气味较淡,果实个体较小[38-39].实验期间,我们发现偶尔有鸟类短暂停留在树上,但没有观察和找到鸟类进食的直接证据,同时也没发现其他个体较大的动物来取食对叶榕果实.这也暗示了对叶榕吸引的取食者可能主要是犬蝠等果蝠,因为它们在寻找食物时嗅觉和视觉发挥了主要作用,且具有相对锋利的牙齿[40],可以把果实咬成破碎的块状.有学者认为动物取食榕果的主要原因是其数量众多而不在于营养价值[41-43],甚至有人认为榕果是一种低营养果实[44-45].但Marcia等人证明榕果能为果蝠提供充足的蛋白质等营养[46].Snow认为含水量较少,且蛋白质含量<0.5%的果实为“贫营养”果实[47];Mattson则把含水量较多,且蛋白质含量在6%~7%之间的果实称为“富营养”果实[48].对叶榕果实营养成分分析表明,其蛋白质含量高达5.8%,对比上面提到的标准可以看出,对叶榕的果实是一种很接近于“富营养”的果实[49],因此能够补偿犬蝠等果蝠在飞行中带走果实所消耗的能量.有学者发现榕果的种子没有毒性、纤维很少或没有[50],因此更适合犬蝠和其他动物取食.对叶榕的种子较小[51],这样在犬蝠等果蝠进食时,有利于通过消化道,可以使其转播到更远的地方.对叶榕果实成熟期平稳,有学者认为这样既可以吸引特化的取食者,还可以减少同种个体间的竞争[52].对叶榕几乎全年挂果,而且树间成熟具有不同步性,这也满足了犬蝠等果蝠的长期食物需求,使其成为稳定的取食者.
犬蝠取食对叶榕,在得到营养的同时,又会对种子起到传播作用,对于其种群的分布和数量的维持起到了重要作用.果蝠在种子传播方面的作用已经得到了充分的肯定和证实[53-54],而且带到不同生境的种子萌发和生长情况也可能发生改变,有研究表明一些植物的种子通过果蝠的消化道后,种子的萌发率会提高[55].唐占辉等人在考察犬蝠取食对叶榕果实后对其种子萌发的影响时发现,虽然萌发率没有提高,但种子的萌发参数有所改变[56].野外在对叶榕母树和其他进食地都发现了果渣,因此我们推测,对于相对大的果实,为了减少飞行中带走食物消耗的能量,犬蝠在母树上进食;对于较小的果实,为了减少被捕食的风险[57],则带到相对安全的进食地.实验中果实鲜重与犬蝠体重之比在50%左右,这使得犬蝠有能力把大多数果实带到其他地方进食,飞行笼实验的观察也证明了这点.研究发现,有的果蝠一次甚至能负载体重1.8~2.5倍的果实[58].犬蝠具有较长和宽大的双翼,有学者认为宽大的翼加上较小的身体更有利于蝙蝠掌控它们的飞行方式[59],而且双翼的利用也有利于它们的取食活动[60].Jason等人更是对果蝠的双翼在操控食物和保持身体姿势方面的作用做了详细研究[61].犬蝠的牙齿相对较发达,这使它有可能采食个体较大的食物,如人心果(Manilkara zapota)[25],所以犬蝠可以比较容易地进食对叶榕果实.视觉和嗅觉在果蝠取食过程中具有重要作用,尤其是嗅觉[62-63].实验中研究的犬蝠具有比较发达的嗅觉和视觉,因此可以充分利用对叶榕果实的气味和果实形状等视觉和嗅觉信号,减少搜寻食物的时间,从而准确定位食物并进行取食活动.
研究结果表明了对叶榕果实所展现的一些特征,为取食者提供足够的信号和丰富的食物补偿.犬蝠则充分利用自身的身体条件和感官系统,来定位和进食其果实,同时它们给予对叶榕以“回报”——传播种子.二者有较好的互利共生关系.
[1] LADLEYJ J,KELL Y D.Dispersal,germination and survival of New Zealand mistletoes(Loranthaceae):dependence on birds [J].New Zealand Journal of Ecology,1996,20(1):69-79.
[2] SALLABANKS R,COURTNEY S P.On fruit-frugivore relationships:variety is the spice of life[J].Oikos,1993,68(3): 567-570.
[3] SNOW D W.Tropical frugivorous birds and their food plants:a world survey[J].Biotropica,1981,13(1):1-14.
[4] KNIGHT R S,SIEGFRIED W R.Inter-relationships between type,size and colour of fruits and dispersal in Southern African trees[J].Oecologia,1983,56:405-412.
[5] WHEELWRIGHT N T.Fruit size,gape width,and the diets of fruit-eating birds[J].Ecology,1985,66(3):808-818.
[6] JANSON C H.Adaptation of fruit morphology to dispersal agents in a neotropical forest[J].Science,1983,219:187-189.
[7] WHEELWRIGHT N T,JANSON C H.Colors of fruit displays of bird-dispersed plants in two tropical forests[J].Am Nat, 1985,126(6):777-799.
[8] WILLSON M F,IRVINE A K,WALSH N G..Vertebrate dispersal syndromes in some Australian and New Zealand plant communities,with geographic comparisons[J].Biotropica,1989,21(2):133-147.
[9] WILLSON M F,WHELAN C J.The evolution of fruit color in fleshy-fruited plants[J].Am Nat,1990,136(6):790-809.
[10] HERRERA C M.Long-term dynamics of Mediterranean frugivorous birds and fleshy fruits:a 12-year study[J].Ecological Monographs,1998,68:511-538.
[11] HERRERA C M.A study of avian frugivores,bird-dispersed plants,and their inter-action in Mediterranean scrublands[J]. Ecological Monographs,1984,54(1):1-23.
[12] HERRERA C M.Seasonal variation is the quality of fruits and diffuse coevolution between plants and dispersers[J].Ecology, 1982,63(3):773-785.
[13] JOHNSON R A,WILLSON M F,THOMPSON J N.Nutritional values of wild fruits and consumption by migrant frugivorous birds[J].Ecology,1985,66(3):819-827.
[14] MURRAY K G,WINNETT-MURRAY K,CROMIE E A,et al.The influence of seed packaging and fruit color on feeding preferences of American robins[G]∥FLEMING T H,ESTRADA A.Frugivory and seed dispersal:ecology and evolutionary aspects.Netherlands,Dordrecht:Kluwer Acadmic Publishers,1993:57-73.
[15] FLEMING T H,VENABLE D L,HERRERA M L G.Opportunism vs.specialization:the evolution of dispersal strategies in fleshy-fruited plants[G]∥FLEMING T H,ESTRADA A.Frugivory and seed dispersal:ecological and evolutionary aspects. Netherlands,Dordrecht:Kluwer Academic Publishers,1993:107-120.
[16] GAUTIER-HION A,DUPLANTIER J M,QURIS R,et al.Fruit characters as a basis of fruit choice and seed dispersal in a tropical forest vertebrate community[J].Oecologia,1985,65(3):324-337.
[17] SCHAEFER H M,LEVEYD J,SCHAEFER V,et al.The role of chromatic and achromatic signals for fruit detection by birds [J].Behaviour Ecology,2006,17(5):784-789.
[18] JANSON C H.Adaptation of fruit morphology to dispersal agents in a Neotropical forest[J].Science,1983,219:187-189.
[19] KAL KOE K V,HERRE E A,HANDLEY C O.Relation of fig fruit characteristics to fruit-eating bats in the new and old world tropics[J].Journal of Biogeography,1996,23:565-576.
[20] 许再富,朱华,杨大荣,等.滇南热带雨林格树类群多样性及生态学意义[G]∥热带植物研究论文报告集.昆明:云南大学出版社,1996:1-15.
[21] 杨大荣,李朝达,杨兵.西双版纳热带雨林中榕树动物群落结构与多样性研究[J].动物学研究,1997,18(2):189-196.
[22] HEITHAUS E R,FLEMING T H,OPLER P A.Foraging patterns and resource utilizetion in seven species of bats in a seasonal tropical forest[J].Ecology,1975,56:841-854.
[23] BONACCORSO F J.Foraging and reproductive ecology in a Panamanian bat community.Bull Florida Stute Mus[J].Biological Science,1979(4):359-408.
[24] BEEHLER B.Frugivory and polygamy in birds of paradise[J].Auk,1983,100(1):1-12.
[25] 唐占辉,盛连喜,曹敏,等.西双版纳地区犬蝠和棕果蝠食性的初步研究[J].兽类学报,2005,25(4):367-372.
[26] LOMA′SCOLO S B,SPERANZA P,KIMBALL R T.Correlated evolution of fig size and color supports the dispersal syndromes hypothesis[J].Oecologia,2008,156:783-796.
[27] CHEN J IN,FLEMING T H,ZHANGLING,et al.Patterns of fruit traits in a tropical rainforest in Xishuangbanna,SW China [J].Acta Oecologica,2004,26:157-164.
[28] 王宪芳,彭艳琼,杨大荣.对叶榕及其传粉榕小蜂的繁殖特点研究[J].云南农业大学学报,2005,20(5):632-635.
[29] 曹敏.热带季节雨林更新动态研究[D].昆明:云南大学,2001.
[30] 吴征镒,陈介,陈书坤,等.云南植物志(第6卷)[M].北京:科学出版社,1995.
[31] ADAMALAI ELANGOVAN,ELANGOVANYUVANA SATYA PRIYA,GANAPATHY MARIMUTHU.Olfactory discrimination ability of the short-nosed fruit batCynopterus sphinx[J].Acta Chiropterologica,2006,8(1):247-253.
[32] MARSHALL A G.Old world phytophagous bats(Megachiroptera)and their food plants:a survey[J].Zoological Journal of the Linnean Society,1985,83:351-369.
[33] SINGARAVELAN N,MARIMUTHU G.Nectar feeding and pollen carrying fromCeiba pentandraby pteropodid bats[J]. Journal of Mammalogy,2004,85:1-7.
[34] ELANGOVAN V,MARIMUTHU G,KUNZ T H.Nectar feeding behaviour of the short-nosed fruit batCynopterus sphinx [J].Acta Chiropterologica,2000(2):1-5.
[35] LEVEY D J.Seed size and fruit-handling techniques of avian fruigivores[J].American Naturalist,1987,129:471-485.
[36] REY P J,GUTIERREZ J E,ALCANTARA J,et al.Fruit size in wild olives:implications for avian seed dispersal[J]. Functional Ecology,1997,11:611-618.
[37] WHEELWRIGHT N T,JANSON C H.Colors of fruit displays of bird-dispersed plants in two tropical forests[J].American Naturalist,1985,126(6):777-799.
[38] BURNS K C,DALEN J L.Foliage color contrasts and adaptive fruit color variation in a bird-dispersed plant community[J]. Oikos,2002,96(3):463-469.
[39] 赵匠,万冬梅,王海涛,等.图牧吉自然保护区大鸨繁殖期种群数量动态和食性分析[J].东北师大学报:自然科学版,2007,39 (2):103-105.
[40] STORZJ F,KUNZ T H.Cynopterus sphinx[J].Mammalian Species,1995,613:1-8.
[41] MORRISON D W.Foraging and day-roosting dynamics of canopy fruit bats in Panama[J].Journal of Mammal,1980,61: 20-29.
[42] FLEMING T H,HEITHAUS E R.Frugivorous bats,seed shadows,and the structure of tropical forests[J].Biotropica,1981, 45:83-87.
[43] THOMAS D W.Fruit intake and energy budgets of frugivorous bats[J].Physiological Zoology,1984,57:457-467.
[44] JORDANO P.Fig-seed predation and dispersal by birds[J].Biotropica,1983,15(1):38-41.
[45] HERBST L H.The role of nitrogen from fruit pulp in the nutrition of the frugivorous batCarollia perspicillata[J]. Biotropica,1986,18(1):39-44.
[46] WENDELN M C,RUNKLE J R.Nutritional values of 14 fig species and bat feeding preferences in Panama[J].Biotropica, 2000,32(3):489-501.
[47] SNOW D W.Evolutionary aspects of fruit-eating by birds[J].Ibis,1971,113:194-201.
[48] MATTSON W J.Herbivory in relation to nitrogen content[J].Annual Review of Ecology and Systematics,1980,11:119-161.
[49] 魏作东,杨大荣,彭艳琼,等.榕树在西双版纳热带雨林生态系统中的作用[J].生态学杂志,2005,24(3):233-237.
[50] JANZEN D H.How to be a fig[J].Annual Review of Ecology and Systematics,1979,10:13-51.
[51] 陈辉,张霜,曹敏.光和温度对西双版纳地区先锋树种对叶榕种子萌发的影响[J].植物生态学报,2008,32(5):1084-1090.
[52] HOWE H F,Estabrook G F.On intraspecific competition for avian dispersers in tropical trees[J].American Naturalist,1977, 111:817-832.
[53] GALETTI M,MORELLATO L P C.Diet of the large fruit-eating batA rtibeus lituratusin a forest fragment in brasil[J]. Mammalia,1994,58:661-665.
[54] FOSTER R B,ARCE J,WACHTER T S.Dispersal and sepuential plant communities Amazonian Peru floodplain[G]∥ASTRATA A,FLEMING T H.Frugivores and seed dispersal.Netherlands:Junk W Publishers,1986:357-370.
[55] UTZURRUM R C B,HEIDEMAN P D.Differential ingestion of viable vs nonviableFicusseeds by fruit bats[J].Biotropica, 1991,23(3):311-312.
[56] 唐占辉,盛连喜,马逊风,等.两种果蝠取食对聚果榕和对叶榕种子萌发的影响[J].生态学报,2007,27(4):1343-1349.
[57] HOWE H F.Fear and frugivory[J].American Naturalist,1979,114:925-931.
[58] FUNAKOSHI K,ZUBAID A.Behavioural and reproductive ecology of the dog-faced fruit batCynopterus brachyotisandC. horsf ieldiin a Malaysian rainforest[J].Mammal Study,1997,22:95-108.
[59] NORBERG U M,RAYNER J M V.Ecological morphology and flight in bats a(Mammalia:Chiroptera):wing adaptations. flight performance,foraging strategy and echolocation[J].Philosophical.Transactions of Royal Society,1987,316:335-427.
[60] STAFFORD B J,THORINGTON R W JR.Carpal development and morphology in archontan mammals[J].Journal of Morphology,1998,235(2):135-155.
[61] JASON DEMETRI VANDOROS,ELIZAABETH RACHEL DUMONT.Use of the wings in manipulative and suspensory behaviors during feeding by frugivorous bats[J].Journal of Experimental Zoology,2004,301:361-366.
[62] LASKA M.Olfactory discrimination ability in short-tailed fruit bat,Carollia perspicillata(Chiroptera:Phyllostomidae)[J]. Journal of Chemical Ecology,1990,16:3291-3299.
[63] RIEGER J F,JAKOB E M.The use of olfaction in food location by frugivorous bats[J].Biotropica,1988,20(2):164-166.
(责任编辑:方 林)
The study on feeding behavior ofCynopterus sphinxonFicus hispida
HU Ya-Ming1,2,TANG Zhan-Hui1,DING Xue-Mei3,SONG Chuan-Tao1,CAO Min2,MA Xun-Feng1
(1.College of Urban and Environmental Sciences,Northeast Normal University,Changchun 130024,China; 2.Xishuangbanna Tropical Botanical Garden,CAS,Mengla 666303,China; 3.College of Animal Science and Veterinary Medicine,Jilin University,Changchun 130062,China)
Figs are very important food resource for many animals in Xishuangbanna,Southwest China, and there is mutually beneficial relation between figs and their frugivore.Observed the hung position fruits ofFicus hispida,and measured some morphological characters of fruits ofF.hispidaand Cynopterus sphinx,which is regular forager to their fruits.Moreover,tried to find out the relation between the information of vision and olfaction offered by fruits ofF.hispidaand the use of sensory system of fruit bats when they foraged the figs in flight cage.C.sphinxcould successfully distinguish the whole fruit ofF.hispida(15.5±1.61 times/h,n=8)from the meshed fruit(10.5±1.45 times/h, n=8)with their vision;When they visited the boxes with and without fruit,C.sphinxpreferred to visit the former box(37.4±7.62 times/h,n=8)rather than to pay visits to the other box(1.1±0.88 times/ h,n=8).The results showed thatF.hispidaand fruit bat adapted to each other very well.
Ficus hispida;fruit bats;flight cage;Xishuangbanna
Q 959.833[学科代码]180·5744
A
1000-1832(2010)03-0132-07
2010-03-23
国家自然科学基金资助项目(30800119);教育部博士点基金资助项目(200802001011);东北师范大学“东师学者”青年学术骨干培养计划项目(120401044);中国科学院西双版纳热带植物园热带森林生态学重点实验室资助项目.
胡亚明(1986—),男,硕士研究生;通讯作者:马逊风(1954—),男,教授,主要从事进化生态学、动物行为生态学研究.
猜你喜欢
云南画报(2021年8期)2021-12-02
故事作文·高年级(2019年6期)2019-06-20
Coco薇(2016年8期)2016-10-09
公民与法治(2016年8期)2016-05-17
爆笑show(2016年2期)2016-04-11
少儿科学周刊·儿童版(2015年6期)2015-11-24
云南画报(2015年10期)2015-01-09
爆笑show(2014年7期)2014-09-03
云南林业(2014年4期)2014-02-28

- 东北师大学报(自然科学版)的其它文章
- 小兴安岭凉水自然保护区土壤昆虫群落特征及多样性研究