
耐力训练对大鼠心肌JAKs和SOCSs表达的影响
2010-08-20 08:02王蕴红赵明华梁蕾张丽琴席利利崔秀亭唐朝枢
中国运动医学杂志 2010年2期
王蕴红 赵明华 梁蕾 张丽琴 席利利 崔秀亭 唐朝枢
1首都体育学院生理生化教研室(北京 100088) 2北京大学医学部生理病理系
长期大负荷训练可引起心脏功能和形态发生一系列改变[1]。研究表明,细胞因子 -6(IL-6)家族成员具有调节心肌功能和刺激心肌细胞肥大的作用,而JAK/信号转导和转录激活因子(signal transducer and activator of transcription,STAT)信号通路是IL-6家族成员发挥诱导心肌肥大的主要信号通路[2]。我们以往的研究也提示,急性运动可引起心肌IL-6家族成员之一的心肌营养因子(cardiotrophin-1,CT-1)及其作用通路JAK/STAT的信号分子STAT3表达的改变[3],这提示JAK/STAT3信号通路可能参与了对运动心脏的功能调节。但是JAK/STAT信号通路在长期训练中是否起调节作用,是否参与了运动性心肌肥大的发生还未见报道。本实验通过测定耐力训练大鼠心肌JAK/STAT3信号通路的信号分子JAKs以及JAK/STAT信号通路内源性的调节蛋白——细胞因子信号抑制因子(suppressors of cytokine signaling,SOCS)的表达,为探讨JAK/STAT信号通路在运动诱发心肌肥大的作用机制提供参考。
1 材料与方法
1.1 实验动物
清洁级雄性SD大鼠40只(购自北京维通利华实验动物中心),体重230±5克,二级动物房常规饲养。大鼠适应环境后随机分为安静对照组(n=10)和耐力训练组(n=30,在运动训练过程中死亡11只)。
1.2 运动方案
参照常芸[4,5]等的大鼠耐力训练方案,采用 10周跑台训练。具体方案是:耐力训练组大鼠每周训练6天,周日休息,第一周起始训练速度15m/min,15min/d,跑台坡度为10°,之后速度每天平均递增1m/min,运动时间递增为2min/d。第2~4周训练从22m/min,30min/d开始,速度平均增加1m/min,时间增加5min/d直至增加到28m/min,60min/d。第 5~6周:速度保持在 28m/min,运动时间保持在60min/d,分别在10°和15°跑台上各运动30min。第7~10周:速度从28m/min开始,每天速度平均约增加0.5~1m/min,直至速度增加到34m/min。运动时间保持40~60min/d,跑台坡度为10°。末次定量负荷运动结束后,分别在运动后即刻和运动后24小时处死,安静对照组大鼠同时处死。三组中共有11只大鼠在各自运动后不同时间段死亡,未解剖确认具体死因,但可排除运动因素以外的传染病或环境因素引起的死亡。
1.3 指标测定
大鼠处死后立即取出心脏,称全心重量后,计算心系数(心重/体重)。于大鼠左心室切取心肌置于10%中性甲醛溶液中固定,常规脱水,石蜡包埋,4μm切片,分别做HE染色和免疫组化染色,免疫组化方法按照试剂盒操作步骤进行。JAK1、JAK3和 SOCS1、SOCS2、SOCS6多克隆抗体购自美国Santa Cruz公司,JAK2购自美国 Cell Signaling公司,其他试剂购自北京中杉金桥生物技术有限公司。
JAK1、JAK2、JAK3 和 SOCS1、SOCS2、SOCS6 一 抗工作效价分别为 1∶50~1∶100,DAB显色。阴性对照片以PBS代替一抗,其余步骤相同。应用Leica DMLB显微镜(SPOT图像分析软件v4.0.2)图像分析仪,在光镜10×20倍视野下阅片照相。每张切片分别在心内膜下、心肌层和心外膜下随机选取3个视野,以细胞核呈棕色染色为阳性细胞[6]。双盲情况下,计数JAK1、JAK2、JAK3和SOCS1、SOCS2、SOCS6细胞的阳性染色细胞占总细胞数的比率。
1.4 统计学分析
2 结果
2.1 耐力训练后大鼠心系数的变化
本研究结果显示,大鼠10周耐力训练后心系数(3.78±0.29)较安静对照组(3.32±0.34)显著增加(P<0.01)。
2.2 大鼠心肌 JAK1、JAK2、JAK3表达
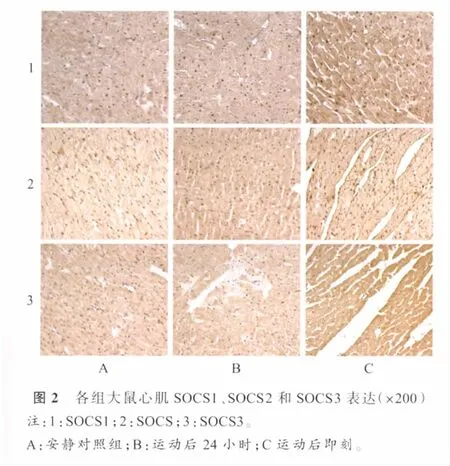
安静对照组大鼠心肌JAK1、JAK2、JAK3即有少量表达,可见于心内膜下、心肌层和心外膜下的心肌细胞以及血管内皮的表达,表达部位分布于胞浆和胞核(图1)。表2显示,耐力训练大鼠运动后即刻心内膜下、心肌层和心外膜下心肌JAK1阳性细胞率显著高于安静对照组(P<0.01)和运动后 24小时组(P<0.05),训练后24小时组未见明显升高(P>0.05)。耐力训练大鼠运动后即刻JAK2阳性细胞率显著高于运动后24小时组(P<0.05)和安静对照组(P<0.01),运动后24小时组也显著高于安静对照组(P<0.05)。耐力训练大鼠运动后即刻和24小时JAK3阳性细胞率与安静对照组比较均无显著差异(P>0.05)。
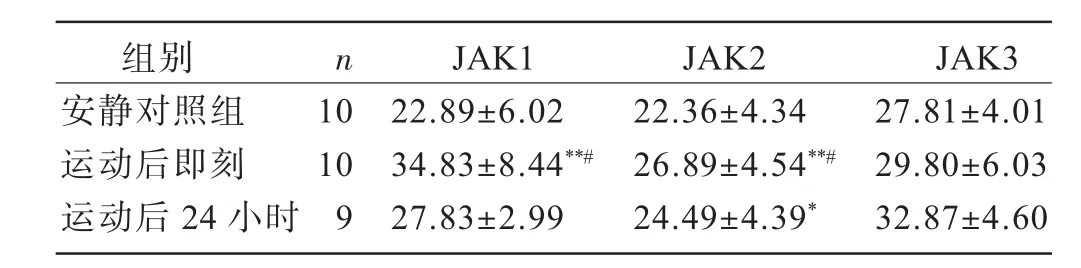
表2 各组大鼠心肌JAK1、JAK2、JAK3阳性细胞率比较(%)

2.3 大鼠心肌 SOCS1、SOCS2、SOCS6表达
安静对照组大鼠心肌SOCS1、SOCS2表达较少。阳性细胞可见于心内膜下、心肌层和心外膜下(图2)。表3显示,耐力训练大鼠心肌SOCS1阳性细胞率在运动后即刻和24小时显著高于安静对照组(P<0.05;P<0.01),运动后即刻组与运动后24小时组之间无显著差异(P>0.05)。SOCS2阳性细胞率在运动后即刻和24小时均显著高于安静对照组(P<0.05),运动后即刻与运动24h后无显著差异(P>0.05)。SOCS6阳性细胞率无论运动后即刻还是运动后24h与安静对照组相比均无显著差异(P>0.05)。
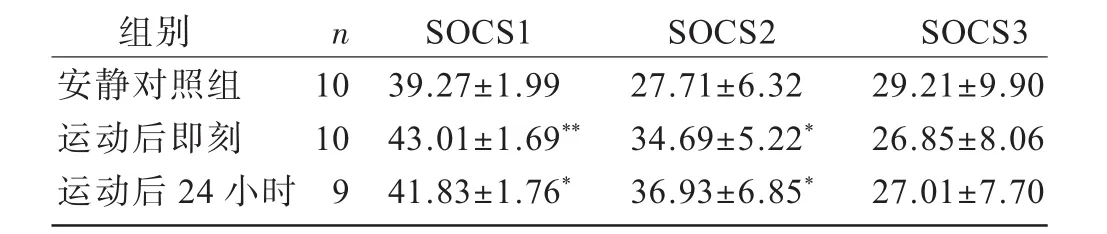
表3 各组大鼠心肌SOCS1、SOCS2、SOCS6阳性细胞率比较(%)
3 讨论
离体实验显示,JAK/STAT信号通路介导了IL-6家族成员调节心肌细胞功能和促心肌细胞肥大的作用。当IL-6等细胞因子刺激心肌细胞时,可见JAK/STAT通路的信号分子mRNA和蛋白表达水平显著升高,而使用JAK2阻断剂AG490后,信号分子mRNA和蛋白表达明显减弱[7]。这一作用在病理性心肌肥大发生中也得到证实。在慢性压力负荷诱发的心脏肥大的发生中,心肌JAK1、p-JAK1、JAK2、p-JAK2 表达明显升高,而当心脏功能失代偿时,它们的表达又明显降低。这提示JAKs的表达及激活变化在诱发心肌肥大中起着重要的作用,并且能反映心脏功能的变化[8]。我们以往的研究显示,急性运动也可导致JAK1、JAK2表达以及p-STAT3/STAT3比值的升高[3,4],表明运动应激也可激活 JAK/STAT通路,并且JAK/STAT信号通路可能参与了运动诱发的心肌功能和代谢变化的调节过程。
本实验结果显示,耐力训练大鼠运动后即刻心肌JAK1、JAK2阳性细胞率显著升高,说明急性运动仍能诱导耐力训练大鼠心肌JAK1、JAK2表达提高,耐力训练所致的心肌功能和形态的变化可能有JAK/STAT信号通路参与调节。此外,耐力训练大鼠心肌JAK1表达与一次急性运动后JAK1的表达规律有所不同,一次急性运动后JAK1阳性细胞率升高一直持续到运动后 24小时[4],而耐力运动大鼠在运动后24小时心肌JAK1阳性细胞率已下降到安静对照水平。这提示耐力训练后大鼠心肌JAK1的表达以及JAK/STAT信号通路对心肌细胞功能的调节发生了适应性改变。而耐力训练大鼠JAK2表达则持续较高,提示训练后JAK2在诱发心肌功能形态改变中作用可能是持续的。
本实验还观察到,JAKs家族的另一个成员JAK3的表达不受运动训练的影响,提示JAK3与JAK1、JAK2在运动中对心肌的作用可能不同。有研究显示JAK3参与了心脏移植排异反应[9]。
JAKs/STAT信号通路受内源性的负性调节机制调节[10]。在对 JAK/STAT通路起负性调节的蛋白质家族中,SOCSs是研究最为广泛的调节蛋白家族。正常情况下,SOCS1、SOCS2等 SOCS mRNA在组织和细胞中均以很低水平存在,而当细胞受到细胞因子等因素的刺激后,SOCS mRNA表达上调,并且同一种细胞因子可诱导SOCS1、SOCS3 等多种 SOCS 家族成员的表达[11]。体外研究还表明[11],SOCSs的过度表达可以抑制许多细胞因子的信号传导。
本实验结果显示,耐力训练大鼠无论在运动后即刻还是运动后24小时,心肌SOCS1、SOCS2阳性细胞率均显著提高(P<0.05),说明耐力训练导致其表达长时间维持在较高水平,这与急性运动后心肌SOCS1表达规律不同[4],同时,也与疾病诱发的心肌肥大中的表达规律不同:发生慢性压力负荷导致心脏肥大时,SOCS1表达无明显变化,只在心衰时明显升高[8]。上述结果是否提示了运动性心肌肥大与疾病因素诱发的心肌肥大预后机制不同,还有待进一步研究证实。
SOCSs对JAK/STAT通路的负反馈调节机制包括:通过SOCS1、SOCS2结合 JAK1、JAK2等 JAK家族成员,竞争性抑制STAT与活化受体的结合而抑制STAT的激活,以及介导信号蛋白依赖蛋白酶的降解等环节,对细胞因子信号传导起负反馈作用[11]。但这种 SOCS1、SOCS2蛋白表达的升高是否意味着其对JAK/STAT信号通路的抑制作用的提高,从而起到减弱细胞因子诱导心肌肥大的作用,仍需进一步研究。
综上所述,长期大负荷耐力训练可引起心肌JAKs和SOCSs表达的改变,这种改变可能在耐力训练影响心肌功能、代谢变化的调节中发挥作用。
[1] Maron BJ,Pelliccia A.The heart of trained athletes:cardiac remodeling and the risks of sports,including sudden death.Circulation,2006,114(15):1633-1644.
[2]KurdiM,Booz GW.Can the protective actionsof JAK-STAT in the heart be exploited therapeutically?Parsing the regulation of Interleukin-6-type cytokine signaling.J Cardiovasc Phyarmaol,2007,50(2):126-141.
[3]王蕴红,王志强,梁蕾,等.运动对心肌营养因子-1与信号转导和转录激活因子3表达的影响.中国运动医学杂志,2009,28(1):85-87.
[4]王蕴红,王杨,席利利,等.急性递增负荷运动对大鼠JAKs和 SOCS1表达的影响.基础医学与临床,2008,28(6):53-57.
[5]常芸,林福美,陈小同,等.运动心脏结构可复性的研究.中国运动医学杂志,1999,18(3):202-207.
[6]杜宏,王扬天,赵明,等.糖尿病大鼠心肌 NF-κB、iNOS、COX-2 表达的研究. 中国病理生理杂志,2008,24(1):81-83.
[7]武利军,赵连友,郑强荪,等.心肌营养素-1诱导大鼠心肌细胞肥大与JAK激酶/信号转导转录活化因子信号通路的关系.高血压杂志,2006,14(3):13-16.
[8]李莹洁,柏树令.慢性压力负荷所致心肌肥厚及心衰大鼠左心室肌JAK/STAT的变化.中国老年学杂志,2004,24(5):444-446.
[9]Changelian PS,Flanagan ME,Ball DJ,et al.Prevention of organ allograft rejection by a specific Janus kinase 3 inhibitor.Science,2003,302(5646):875-878.
[10] Krebs DL,Hilton DJ.SOCS:physiological suppressors of cytokine signaling.JCell Sci,2000,113(Pt 16):2813-2819.
[11]Krebs DL,Hilton DJ.SOCS proteins:negative regulators of cytokine signaling.Stem Cells,2001,19(5):378-387.
猜你喜欢
中学生天地(A版)(2022年9期)2022-10-31
小哥白尼(军事科学)(2022年5期)2022-08-17
动漫界·幼教365(小班)(2020年8期)2020-07-04
野生动物学报(2020年1期)2020-02-21
散文诗(2017年18期)2018-01-31
车迷(2017年10期)2018-01-18
中国高原医学与生物学杂志(2017年4期)2017-03-08
中国组织化学与细胞化学杂志(2016年3期)2016-02-27
军事体育学报(2014年3期)2014-02-27
卒中与神经疾病(2011年3期)2011-06-14
