
非主要生长季节白桦人工林林隙内伐根对土壤呼吸速率的影响1)
2010-08-09 11:07:18孟春王俭
东北林业大学学报 2010年11期
孟 春 王 俭
(森林持续经营与环境微生物工程黑龙江省重点实验室(东北林业大学),哈尔滨,150040)
伐根在森林生态系统中占有重要的地位,对林内微环境会产生影响[1]。国内外对伐根的研究较多[2],包括伐根与养分循环[3]、伐根与更新[4-5]、伐根与动物及真菌[6-8]等;但是,伐根对林地的碳循环的研究却未见报道,而林地碳循环对于目前全球气候变化有重要的影响[9-11]。因此,笔者对白桦人工林非主要生长季节林隙内白桦伐根对土壤呼吸速率水平空间分布影响进行了研究,以揭示林隙内土壤呼吸速率水平空间分布特点及主要影响因素,掌握不同腐朽程度伐根对林隙土壤呼吸速率的影响规律。
1 研究样地概况
东北林业大学哈尔滨实验林场位于哈尔滨市区内马家沟河西岸(45°43.463'~45°43.464'N,126°37.457'~126°37.458'E),地形平缓,土壤为地带性黑钙土,水分条件良好。属于温带季风性气候,年平均气温3.5℃,年积温2 757℃,年降水量534 mm。
本次试验选取该林场内的白桦(BetulaplatyphyllaSuk.)林作为研究对象,该林分于1960年春季采用2年实生苗造林。初植密度0.5 m×1 m,当年成活率95%。前3 a按常规方法进行除草、抚育等田间管理。1964年隔一行去一行,变株距为1 m×1 m,1990年进行疏伐。以后根据试验及教学的需要,对林内白桦有不定期的采伐。目前,每0.5 hm2林地上,活立木493株,平均高19 m,平均胸径16 cm。活立木蓄积53.81 m3。下草盖度20%。
2 研究方法
2.1 林隙的选取
经踏查后,本次研究选取该林分内5个林隙作为研究对象(A、B、C、D、E)。其中:4 个林隙为有伐根林隙(A、B、C、D);1个林隙为无伐根林隙(E),作为对照。4个有伐根林隙中,各伐根腐朽程度不同的伐根分别为:无腐朽、初步腐朽和严重腐朽。各林隙状况见表1。
2.2 土壤呼吸速率的测定
土壤呼吸速率存在显著的日变化特征[12-14],昼夜内不同时间观测,土壤表面呼吸速率差异较大。为准确观测各观测点的土壤呼吸速率,消除不同观测时间及观测时间延续对观测结果的影响,于2008年10月6日,对该林分进行了24 h土壤呼吸连续观测。结果表明,土壤呼吸速率昼夜均值发生在9:00时,而7:00—11:00时,土壤呼吸速率在土壤呼吸速率昼夜均值的±5%之间。所以,本次研究中林隙内土壤呼吸速率的观测时间选择在8:00—10:00时。
为观测林隙土壤呼吸速率,于2008年10月9日上午,在5个林隙内分别沿东西和南北正交方向选取相距约1 m的观测点。其中:A、B林隙选取11个土壤观测点和1个伐根表面观测点;C林隙选取12个土壤观测点和1个伐根表面观测点;D林隙选取10个土壤观测点和1个伐根表面观测点;E林隙选取12个土壤观测点,其位置见图1。在每个土壤观测点插入直径为10.5 cm的PVC环,在每个伐根表面观测点上方置1个直径为10.5 cm的PVC环,并用玻璃胶将PVC环与伐根表面胶合并使二者接触处密闭。其中:无伐根林隙的南北向为林隙的长轴方向,东西向为林隙的短轴方向;4个有伐根林隙的南北方向为林隙的短轴方向,东西向为林隙的长轴方向。2008年10月10日、11日、12日和13日9:00—10:00时,使用LI-8100型全自动土壤CO2排放通量的全自动测量系统[15],对设定的各林隙内观测点进行土壤CO2呼吸速率的观测,同时观测地表温度以及地表下10 cm处的温度和湿度。每日观测1个林隙。
在进行林隙土壤呼吸速率观测的同时,测定伐根表面呼吸速率。
土壤呼吸观测完毕后,取出各观测点PVC环,用环刀分别取各观测点地表下0~5 cm和10~15 cm处土样,混合后供土壤有机质分析用。土壤有机质含量采用水合热法测定[16]。

表1 伐根性状
2.3 数据处理
观测数据经LI-8100系统软件导出后,使用SPSS11.0统计软件计算土壤呼吸速率均值、最大值、最小值、标准差、方差和变异系数;进行单因素方差分析和各林隙土壤呼吸速率与温度和湿度的相关分析。使用ARCVIEW3.1软件绘制各林隙土壤呼吸速率等值线。
3 结果与分析
3.1 伐根腐朽程度及伐根表面呼吸速率
木材腐朽有白色腐朽、褐色腐朽和软腐朽3种类型,分别由不同种的真菌引起,结果造成木材纤维、木质素等的分解[17]。腐朽发生的不同阶段,真菌的种类和数量不同[18],并产生不同的宏观构造特征。
真菌侵蚀木材必须具备一定的条件,这些条件包括营养体、水分、温度和空气等。伐根表面裸露于大气中,在近地表的空间范围内,水分和空气充足,温度适宜,一定体积的伐根提供了真菌生存的营养体条件,真菌侵蚀伐根成为必然。这一侵蚀将从裸露于大气中的伐根表面开始,随着侵蚀程度的加剧,伐根表面有机体被分解,结构松散,为真菌侵蚀地下部分伐根创造了条件。因此,伐根的地下根系将逐渐被侵蚀,直至全部被分解完毕。
A林隙伐根是活的生命体,自养呼吸在进行。B林隙伐根作为生命体已开始死亡,自养呼吸作用微弱,表面开始被真菌侵蚀,少量真菌分解有机体的作用产生了一定量的异养呼吸,但呼吸总量不及活生命体的自养呼吸量,所以,A林隙伐根表面的呼吸速率略高于B伐根表面呼吸速率(表2)。C和D林隙伐根表面已严重腐朽,真菌数量众多,异养呼吸强度高,伐根表面呼吸速率分别达到15.195、31.869 μmol·m-2·s-1。
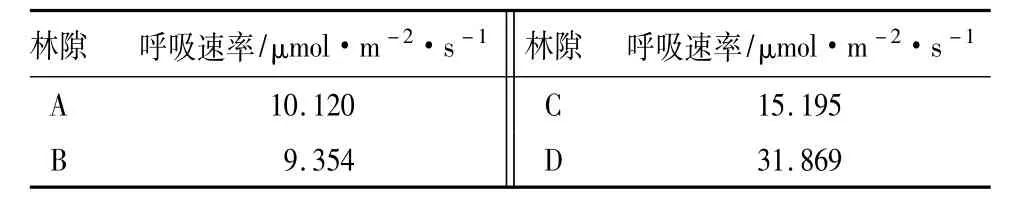
表2 伐根表面呼吸速率
3.2 土壤呼吸速率均值
观测结果表明,各林隙土壤呼吸速率不同,经单因素方差分析,各林隙土壤呼吸速率差异显著(F=5.373,F(α=0.05)=0.001,F>F(α=0.05))。有伐根林隙土壤呼吸速率均值高于无伐根林隙土壤呼吸速率均值(表3),4个有伐根林隙土壤呼吸速率高于无伐根林隙土壤呼吸速率6.18%~40.70%。
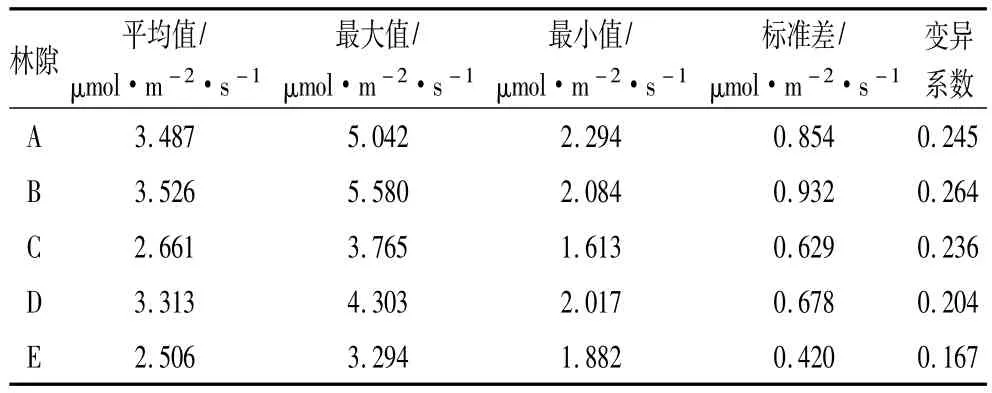
表3 林隙土壤呼吸速率
有伐根林隙,由于伐根的存在,增加了单位面积地下根生物量,碳源增加。因此,在观测期内土壤呼吸速率高于无伐根林隙土壤呼吸速率。初步腐朽伐根地下根系自养呼吸作用仍然存在,而严重腐朽伐根地下根系自养作用消失,木腐菌分解根系的异养呼吸作用还很微弱。因此,4个有伐根林隙中,初步腐朽伐根林隙(B)土壤呼吸速率最高,其值为3.526μmol·m-2·s-1;严重褐腐伐根林隙(C和D)土壤呼吸速率低于初步腐朽伐根林隙(B)土壤呼吸速率,其值分别为2.661、3.313 μmol·m-2·s-1。
3.3 土壤呼吸速率空间分布特点
各林隙内土壤呼吸速率均存在变异性,即水平空间异质性。有伐根林隙土壤呼吸速率异质性高于无伐根林隙土壤呼吸速率异质性,有伐根林隙土壤呼吸速率变异系数介于0.204~0.264之间,无伐根林隙土壤呼吸速率变异系数仅为0.167(表3)。
有伐根林隙中,B林隙土壤呼吸速率变异系数最大,其值为0.264;A、C和D林隙土壤呼吸速率变异系数依次为0.245、0.236和0.204。即:伐根初步腐朽林隙土壤呼吸速率变异性高于无腐朽伐根林隙土壤呼吸速率变异系数,而无腐朽伐根林隙土壤呼吸速率变异系数又高于严重腐朽伐根林隙土壤呼吸速率变异系数。初步腐朽伐根地下根系占据了一定的土壤空间,使土壤异质性增加,但是,由于地下根系自养呼吸消失,异养呼吸甚微,便造成林隙土壤呼吸速率具有较高的变异系数。未腐朽伐根地下根系虽增加了土壤异质性,但由于自养呼吸仍然存在,所以,土壤呼吸变异系数略小于初步腐朽伐根林隙土壤呼吸变异系数。严重腐朽伐根由于地下根系已经开始分解,根系与土壤间的物质交换使土壤异质性降低,因此,土壤呼吸速率变异系数较小。
对各林隙土壤呼吸速率等值线分析表明,有伐根林隙土壤呼吸速率等值线有规律可循,无伐根林隙土壤呼吸速率则无规律可循(图1)。
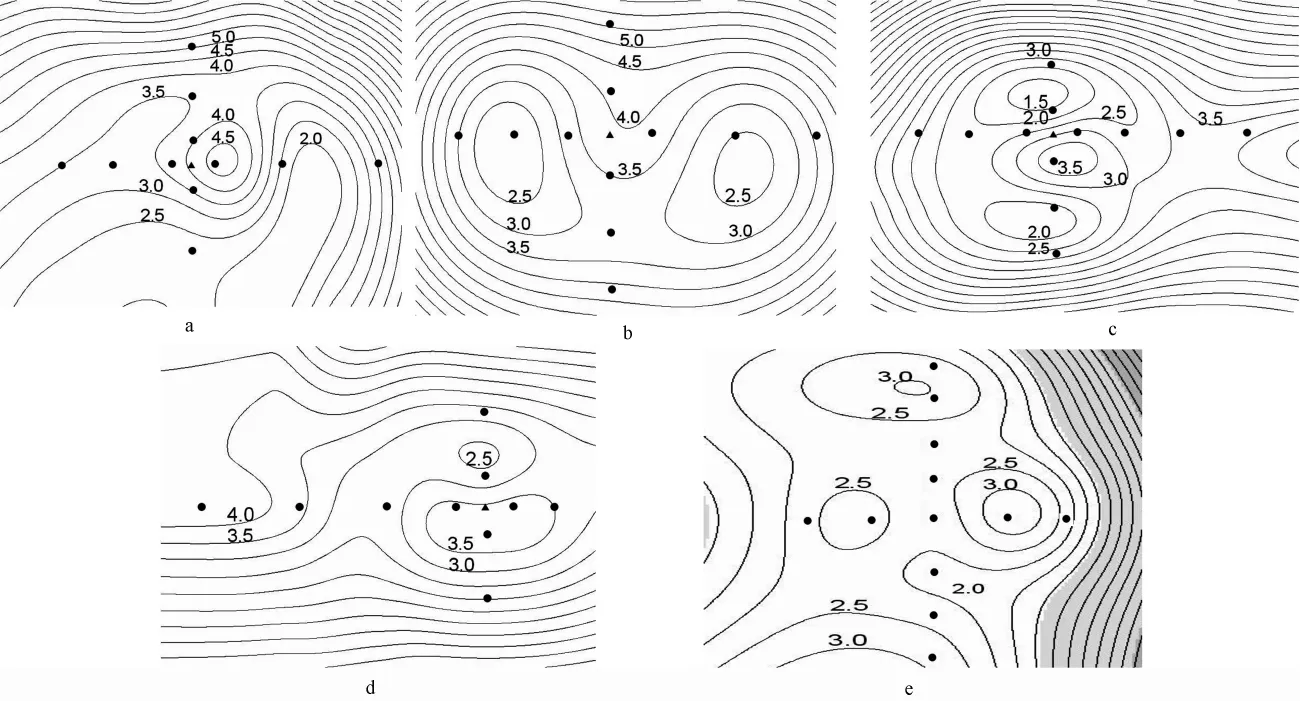
图1 林隙土壤呼吸速率等值线图
有伐根林隙土壤呼吸速率呈现以伐根为中心向四周辐射,土壤呼吸速率由高变低、再变高的变化规律。这一规律在长轴方向比短轴方向体现得明显。从土壤呼吸速率数值上看,伐根附近的土壤呼吸速率略低于林隙边缘处土壤呼吸速率。有伐根林隙土壤呼吸速率的这种水平分布格局,源于林木伐根根系以伐根为中心向四周伸展,在伸展方向上,随伸展半径的增加,单位面积根系生物量减少,因此,由根系分解产生的异养呼吸速率在降低,使土壤呼吸速率降低。当伸展半径再增加,即接近或到达林隙边缘时,由于林隙边缘活立木根系自养呼吸作用的增强,又使土壤呼吸速率增加。无伐根林隙内根系由林隙边缘立木根系伸展而来,随机性较大,因此,无伐根林隙土壤呼吸速率异质性无规律可循。
4个有伐根林隙中,土壤呼吸速率变化梯度不同(图2)。A、B、D林隙长轴方向土壤呼吸速率变化梯度大于短轴方向,A 林隙变化梯度最大值在 0.868~0.992μmol·m-2·s-1之间,B 林隙次之,为0.724~0.904 μmol·m-2·s-1,D 林隙为0.528~0.615 μmol·m-2·s-1,即随伐根腐朽程度的增加,变化梯度最大值降低,这一现象的原因如前所述;C林隙短轴方向土壤呼吸速率变化梯度大于长轴方向,变化梯度达0.824~0.962 μmol·m-2·s-1。

图2 有伐根林隙土壤呼吸速率变化梯度图
3.4 土壤呼吸速率与主要影响因素的关系
研究表明,土壤温度和湿度是决定土壤呼吸速率变异的两个主要因素,通常情况下,可解释土壤呼吸速率变异的60%~80%[19]。但是,本研究中各林隙土壤呼吸速率水平空间变化趋势和土壤温度及湿度水平空间变化趋势没有出现一致的现象[20],各林隙土壤呼吸速率变异性与土壤温度(土壤表面温度、土壤表面下10 cm处温度)和湿度的相关性并不高,相关系数(R2)介于0.001~0.601 8之间,低于通常研究的结果,表明土壤温度和湿度不是影响林隙内土壤呼吸速率水平空间分布的主要原因。
土壤有机质是土壤呼吸的基质[21],其含量的多少,可影响呼吸强度的大小[22-23]。但是,本研究中各林隙内土壤呼吸速率水平空间变化趋势和土壤有机质含量的水平空间变化趋势不同。对各林隙内各观测点土壤表面呼吸速率与有机质含量进行线性和二次曲线相关分析,结果表明:采用线性相关模型,相关系数(R2)介于0.001~0.163之间;采用二次曲线模型,相关系数(R2)介于0.012~0.410之间,表明土壤有机质含量并不是决定林隙内土壤表面呼吸速率水平异质性的决定因素。
以伐根表面呼吸速率表征伐根腐朽程度,对4个有伐根林隙土壤呼吸速率均值(Rs)与伐根呼吸速率(Rg)进行相关分析,其结果采用三次曲线表示:Rs=5.548-0.224Rg+0R3g,相关系数(R2)为0.989。表明伐根的不同程度腐朽是影响有伐根林隙土壤呼吸速率水平分布的主要因素。
4 结论
采伐作业遗留在林地的伐根,成为林隙微环境的一个重要环境因子。伐根的自养呼吸和异养呼吸作用在一个较长的时期内存在,影响了林隙内土壤呼吸速率的大小和水平空间分布,并且,由于伐根的“碳源”作用,使有伐根林隙土壤呼吸速率高于无伐根林隙土壤呼吸速率。
有伐根林隙土壤呼吸速率相对于无伐根林隙土壤呼吸速率呈现出以伐根为中心向四周辐射,土壤呼吸速率由高变低、再变高的规律变化。有伐根林隙随伐根腐朽程度的增加,土壤呼吸速率水平空间异质性变化梯度最大值降低。
伐根腐朽程度可以表征整个伐根被木腐菌侵蚀的状况,若以伐根表面呼吸速率来表征伐根腐朽程度,则伐根腐朽程度是影响有伐根林隙土壤呼吸速率的主要因素。
目前,尚未见到其他树种伐根对林隙土壤呼吸影响研究的报道。作为森林采伐活动大量留在林地上的各个树种的伐根,有必要就其对林隙的影响,尤其是目前敏感的碳循环的影响作全面、深入的研究。
[1]孟春,庞凤艳.伐根研究进展[J].森林工程,2005,21(4):11-13.
[2]法蕾,李文宁.试述粗木质残体及伐根分解的研究进展[J].地质技术经济管理,2004,26(6):54-56.
[3]Hendrickson OQ.Abundance and activity of N2-fixing bacteria in decaying wood[J].Canadian Journal of Forest Research,1991,21(9):1299-1304.
[4]Jobidon R.Stump height effects on sprouting of mountain maple,paper birch and pin cherry—10year results[J].Forestry Chronicle,1997,73(5):590-595.
[5]Mclaren K P,McDonald M A.Coppice regrowth in a disturbed tropical dry lime stone forest in Jamaica[J].Forest Ecology and Management,2003,180(1/3):99-111.
[6]Safranyi L,Shore T L,Carroll A L,et al.Bark beetle(Coleoptera:Scolytidae)diversity in spaced and unmanaged mature lodgepole pine(Pinaceae)in southeastern British Columbia[J].Forest Ecology and Management,2004,200(1/3):23-38.
[7]Lindhe A,LindelöwÅ.Cut high stumps of spruce,birch,aspen and oak as breeding substrates for saproxylic beetles[J].Forest E-cology and Management,2004,203(1/3):1-20.
[8]田子珩,张春雨,赵秀海.红松伐根分解过程中土壤动物动态[J].生态学杂志,2007,26(2):286-290.
[9]Jenkinson DS,Harkness D D,Vance E D.Calculating net Primary Production and annual input of organic matter to soil from the amount and radiocarbon content of soil organic matter[J].Soil Biology and Biochemistry,1992,24(4):295-308.
[10]Davidson E A,Savage K,Verehot L V,et al.Minimizing artifacts and biases in chamber-based measurements of soil respiration[J].Agricultural and Forest Meteorology,2002,113(1/4):21-37.
[11]杨玉盛,董彬,谢锦升,等.森林土壤呼吸及其对全球变化的相应[J].生态学报,2004,24(3):583-591.
[12]冯朝阳,吕世海,高吉喜,等.华北山地不同植被类型土壤呼吸特征研究[J].北京林业大学学报,2008,30(2):20-26.
[13]肖复明,汪思龙,杜天真,等.杉木人工林林地土壤呼吸研究[J].江西农业大学学报,2005,27(4):580-584.
[14]陈光水,杨玉盛,吕萍萍,等.中国森林土壤呼吸模式[J].生态学报,2008,28(4):1748-1761.
[15]赵广东,王兵,杨晶,等.LI-8100开路式土壤碳通量测量系统及其应用[J].气象科技,2005,33(4):363-366.
[16]邵则瑶,黄业海,李酋开.水合热法测定土壤有机质的研究[J].土壤通报,1980(3):29-31.
[17]魏玉莲,戴玉成.木材腐朽菌在森林生态系统中的功能[J].应用生态学报,2004,15(10):1935-1938.
[18]法蕾,赵秀海.阔叶红松林山杨伐根分解过程中真菌动态研究[J].北京林业大学学报,2006,28(增刊2):90-94.
[19]Rodeghiero M,Cescatti A.Main determinants of forest soil respiration along an elevation/temperature gradient in the Italian Alps[J].Global Change Biology,2005,11(7):1024-1041.
[20]孟春,王立海,王俭.人工林白桦伐根腐朽程度对林隙内土壤温度的影响[J].东北林业大学学报,2009,37(9):57-60.
[21]Sikora L J,McCoy JL.Attempts to determine available carbon in soil[J].Biology and Fertility of Soils,1990,9(1):19-24.
[22]Bazzaz FA,Williams WE.Atmospheric CO2concentrations within a mixed forest:Implications for seedling growth[J].Ecology,1991,72(1):12-16.
[23]Russell C A,Voroney R P.Carbon dioxide efflux from the floor of a boreal aspen forest.I.Relationship to environmental variables and estimates of C respired[J].Canadian Journal of Soil Science,1998,78(2):301-310.
猜你喜欢
现代企业(2021年2期)2021-07-20 07:57:10
四川劳动保障(2021年3期)2021-06-09 07:08:56
智能城市(2021年3期)2021-04-12 04:40:50
绿色科技(2019年12期)2019-07-15 11:13:02
现代园艺(2017年13期)2018-01-19 02:28:05
西南农业学报(2016年5期)2016-05-17 05:42:36
水利科技与经济(2016年10期)2016-04-26 08:39:58
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:09
邯郸职业技术学院学报(2016年2期)2016-02-27 13:39:26
山东建筑大学学报(2015年4期)2015-05-11 09:05:22
