
濒危植物白桂木的遗传多样性研究
2010-07-24 01:30范繁荣
浙江农林大学学报 2010年2期
范繁荣
(福建三明林业学校, 福建 三明365001)
白桂木Artocarpus hypargyreus 是桑科Moraceae 波罗蜜属Artocarpus 常绿乔木, 是一种经济价值很高的树种。 种子含有丰富的白桂木凝集素, 正引起医药学界的广泛注意。 白桂木树干高大, 树形优美, 枝叶繁茂, 遮荫面宽, 也是园林绿化的优良树种。 现主要零星分布于福建南部和西部、 江西南部、 湖南东南部和西南部、 广东北部、 海南、 广西、 云南东南部。 长期以来, 由于人们对白桂木天然林的破坏性经营, 加上白桂木自身生物学特性及自然环境条件变化等方面的原因, 致使白桂木野生资源接近枯竭, 处于濒危境地, 已被列为国家三级濒危保护树种[1]。 目前, 关于白桂木的研究很少, 仅局限于白桂木药用化学成分的研究[2-6]和白桂木形态特征、 生态特性及地理分布方面的报道[7], 对白桂木濒危的内在机制及保育技术研究还是空白。 遗传多样性是生物种内表现在从DNA 分子到形态特征各个层次上可遗传所有变异的总称, 是濒危植物保护生物学研究的核心之一[8]。 对濒危物种遗传多样性和群体遗传结构的研究是揭示其适应潜力的基础, 也为进一步探讨濒危物种的濒危机制和制定相应的保护措施提供科学依据[9]。 随机扩增多态性DNA(RAPD, random amplified polymorphic DNA)分析不受器官和组织种类、 发育阶段和生境条件等因素的影响, 需要的DNA 模板量少, 质量要求不高,多态性丰富, 具有简便、 快速、 易行、 价廉等优点, 为从分子水平研究生物遗传多样性中应用最广和最具代表性的方法之一[10]。 RAPD 分子标记用于属以下等级特别是种内各居群或品种的研究具有很强的优越性[11]。 本研究利用RAPD 分子标记技术对福建不同白桂木天然种群的遗传多样性进行分析, 为从遗传多样性角度揭示白桂木的濒危机制和更好地保护白桂木种群提供科学依据。
1 材料与方法
1.1 材料采集
实验材料分别采自福建白桂木保存较好的三明市梅列区陈大、 三元区莘口, 泉州市永春县牛姆林, 清流县沙芜, 永安市贡川的白桂木天然林。 由于各个种群的白桂木个体数量较少, 因此, 只能在各个采样区随机选取生长良好的3 株白桂木(株间间隔必须5 m 以上), 采集白桂木健康小枝, 并将它们迅速放入密封塑料袋中, 装入冰壶带回实验室, 放在- 80 ℃超低温冰箱保存备用。
1.2 DNA 提取
采用改进的十六烷基三甲基溴化铵(CTAB)法, 参照王关林等[12]的方法提取DNA。 称取叶片0.3 g左右, 加液氮快速充分研磨至粉末状, 加入0.1 g 聚乙烯吡咯烷酮(PVP), 将粉末移至1.5 mL 离心管中; 加入600 μL·管-1预热至65 ℃的2 × CTAB 提取缓冲液, 同时加入7 μL 2-琉基乙醇, 充分混匀,65 ℃水浴30 min, 其间颠倒几次; 取出离心管, 冷至室温, 加等体积的氯仿/异戊醇(24 ∶1), 轻缓颠倒混匀, 静置10 min, 室温下1.2 万r·min-1离心10 min; 取上清液至另一离心管, 加2/3 体积异丙醇, 轻缓颠倒混匀, 室温放置15 min, 于4 ℃条件下1.2 万r·min-1离心10 min; 用体积分数为70%乙醇洗涤DNA 2~3 次, 风干, 将DNA 溶于500 μL TE(tris-EDTA, 三羟甲基氨基甲烷-乙二胺四乙酸)溶液, 加入质量浓度为50 g·L-1的RNase A, 并于37 ℃保温1 h; 用等体积氯仿/异戊醇(24 ∶1)抽提1 ~3 次; 上清液中加入终浓度为0.2 ~ 0.4 mol·L-1的氯化钠, 2 倍体积的无水乙醇, 放置1 h 左右, 1.2万r·min-1离心10 min; 除去上清液; 用体积分数为70%乙醇洗涤沉淀2 ~ 3 次, 风干后溶于50 μL TE 中备用。
1.3 聚合酶链式反应(PCR)扩增与产物的检测
扩增体系采用25.0 μL 体系: 25 ng 模板DNA, 0.40 μmol·L-1引物, 0.15 mmol·L-1三磷酸碱基脱氧核苷酸(dNTP), 16.67 nkat TaqDNA 聚合酶, 1.50 mmol·L-1氯化镁, 2.5 μL 10 ×聚合酶链式反应(PCR)缓冲液, 加入灭菌的超纯水至25.0 μL。
PCR 扩增条件为: 94 ℃预变性5 min, 94 ℃变性1 min, 36 ℃1 min, 72 ℃延伸2 min, 共40 个循环, 72 ℃延伸10 min。 扩增完毕, 取经15.0 g·L-1琼脂糖凝胶电泳检查。
1.4 数据统计与分析
RAPD 是显性标记, 同一引物扩增产物中电泳迁移率一致的条带被认为具有同源性。 根据反应产物在凝胶对应位置上条带的有无, 有带记为1, 无带记为0, 得到RAPD 分析的原始数据。 使用POPGEN 32 软件计算多态位点百分率、 Shannon 指数、 Nei 指数和基因流(Nm); 运用Nei 指数法[13]计算遗传距离 (D)和遗传一致度 (F), 聚类分析运用DPS 软件采用非加权配对算术平均法 (UPGMA, unweighted pai-group method using an arithmetic average)分析。
2 结果与分析
2.1 引物筛选
随机引物购自上海生工生物工程有限公司。 用5 个DNA 样品(每个种群选1 个样品)对随机引物进行筛选, 从60 个引物中筛选出S237, S248,S249, S262, S263, S271 等6 个扩增条带清晰、 重复性好的引物(表1), 对所有个体进行扩增。

表1 RAPD 分析的6 个随机引物序列Table 1 Sequences of 6 random primers used in RAPD analysis
2.2 白桂木种群遗传多样性分析
2.2.1 多态位点百分率 利用6 个随机引物对5 个白桂木种群15 个个体DNA 样品进行RAPD 分析,共扩增出37 个条带, 其中多态条带30 个, 总的多态位点百分率为81.08%, 平均可获得多态位点5个·引物-1。 引物S263, S269 和S271 对白桂木不同个体扩增的带型见图1 ~ 3。 各居群间多态位点百分率存在着较大差异, 其中永安种群最高, 为24.32%, 梅列种群最低, 为8.11%, 平均多态位点百分率为15.67%, 其大小排序为永安种群>三元种群>永春种群>清流种群>梅列种群。
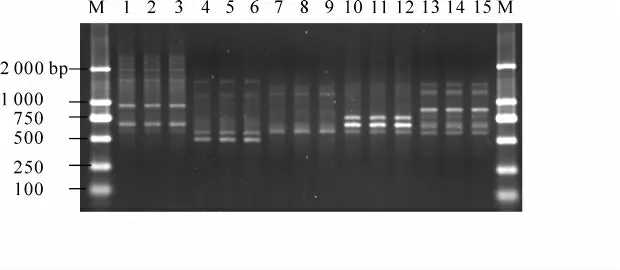
图1 引物S263 扩增的RAPD 带型Figure 1 RAPD amplification products from primer S263
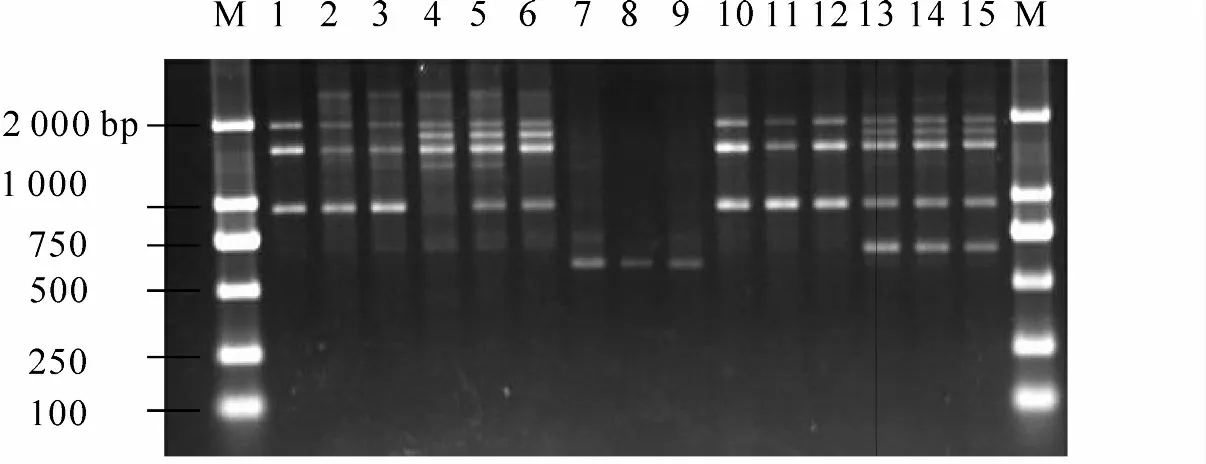
图2 引物S269 扩增的RAPD 带型Figure 2 RAPD amplification products from primer S269

图3 引物S271 扩增的RAPD 带型Figure 3 RAPD amplification products from primer S271
2.2.2 Shannon 多样性和Nei 基因多样性指数 Shannon 指数是生态学中用于度量物种多样性最常用的指标[14]。 从表2 的估算结果可看出, 白桂木总遗传多样性指数为0.231 9, 各种群平均多样性指数为0.219 3, 其中最高的是永安种群, 为0.261 7, 其大小排序为清流种群>三元种群>梅列种群>永春种群, 最小的永春种群仅为0.181 9。 从表2 中可见, 由Nei 指数估算的白桂木总遗传多样性为0.166 0, 各种群平均
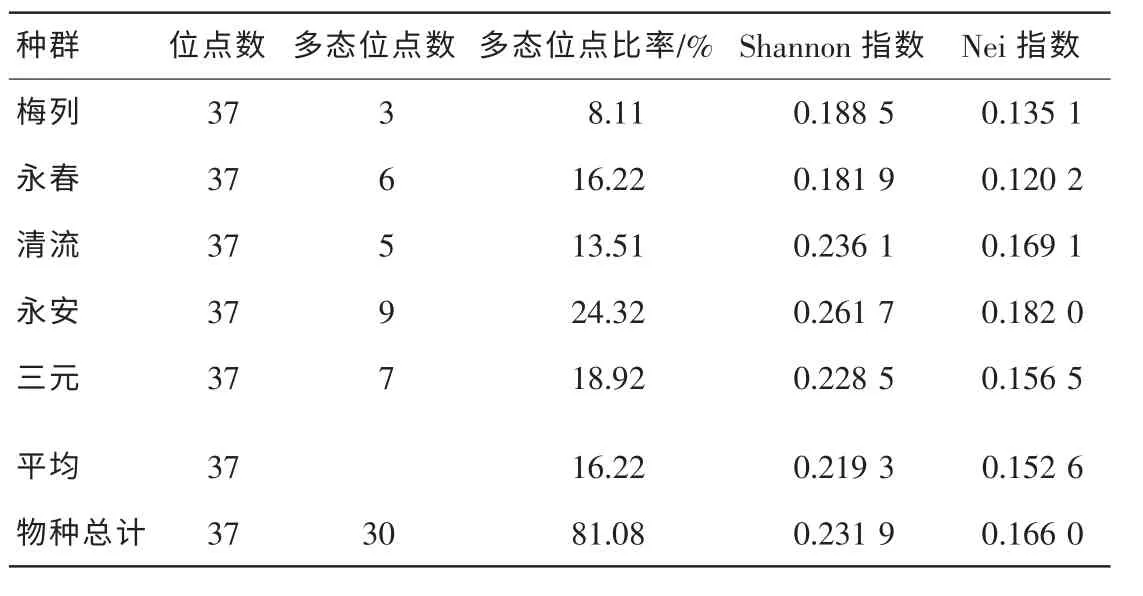
表2 不同白桂木种群的遗传多样性Table 2 Genetic diversity of Artocarpus hypargyreus
遗传多样性为0.152 6, 其中最高的是永安种群, 为0.182 0, 其他大小排序为清流种群>三元种群>梅列种群>永春种群, 最小的永春种群仅为0.120 2。 由该指数估算的遗传多样性较Shannon 指数估算的低, 但其大小排序与Shannon 多样性指数基本一致。 与濒危植物银杉Cathaya argyrophylla 和珙桐Davidia involucrata 相比, 白桂木的遗传多样性高于银杉[11], 低于珙桐[15]。
2.3 白桂木种群间的遗传分化
由Nei 指数计算的白桂木种群间的遗传分化系数结果见表3。 白桂木种群间的分化系数GST为0.737 8, 白桂木的遗传变异有73.78%存在于种群之间, 只有26.22%的遗传变异存在于种群内, 这与苏何玲等[16]对濒危植物冷杉Abies ziyuanensis 的研究结果相似。 可见白桂木种群间的分化程度较高, 种群内的分化程度较低。
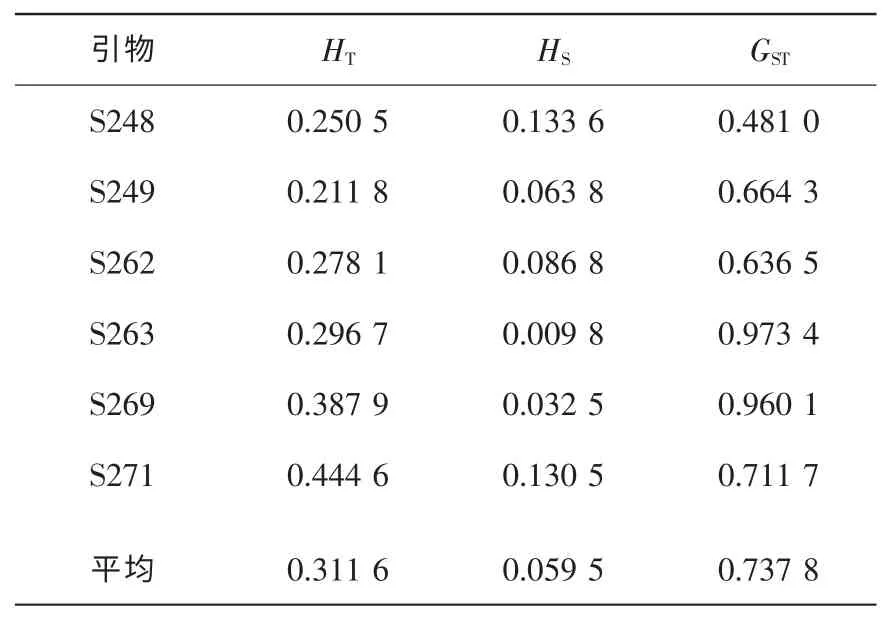
表3 Nei 指数估算的白桂木种群间遗传分化Table 3 Genetic differentiations among 5 populations estimated by Nei’ index
基因流Nm的计算结果为0.125 1, 表明白桂木种群间的基因流动较小, 这与由Nei 指数计算的种群间的遗传分化系数所得的大部分遗传变异存在于种群间的结论相一致。 Wright 认为种群间基因流大于1, 则能发挥其均质化作用; 反之若小于1, 则说明基因流成为遗传分化的主要原因[17]。 白桂木种群在自然状态下分布零散, 各种群呈“岛屿” 状, 种群间的空间距离远, 花粉的传播受限, 同时其生殖生态学研究发现白桂木个体数量少、 雄雌花序的分布不利于授粉、 开花时期雨水多等原因造成结实量低, 加上动物的取食, 自然状态下可供更新的有效种子数量少。 同时种子含水量高, 不易储存, 种子有后熟现象, 在自然条件下发芽困难, 种子成苗率低, 自然更新能力差。 有效种子流的形成受到极大的限制, 使得白桂木种群间的基因流极低, 只有0.125 1, 远小于1.000, 极低的基因流造成种群间很高的遗传分化。
2.4 白桂木种群的遗传距离与遗传相似度
遗传距离和遗传相似度是衡量植物变异水平的重要指标。 白桂木种群间的遗传距离和遗传相似度计算结果见表4。 从表4 中可看出, 梅列种群与三元种群的相似度最大, 为0.747 7, 它们之间的地理距离最近, 永春种群与清流种群的的相似度最小, 为0.528 7, 它们之间的地理距离也是最大的。 遗传距离则是永春种群与清流种群最大,为0.637 3, 梅列种群与三元种群的遗传距离最小, 为0.290 7, 5 个不同种群的平均相似度和遗传距离分别为0.6665 和0.411 8。
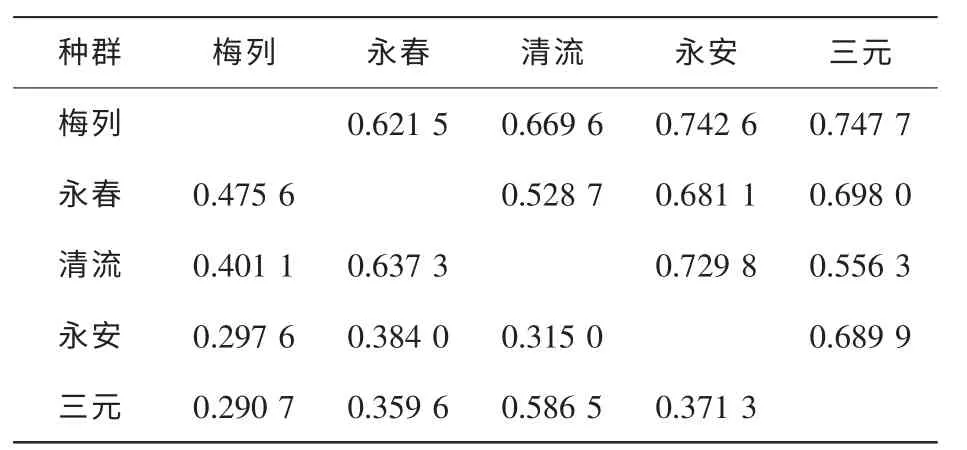
表4 不同白桂木种群的遗传相似度与遗传距离Table 4 Genetic distance and genetic identity of populations of Artocarpus hypargyreus
2.5 聚类分析
种群间的遗传距离一般随着地理距离的增大而增大[18]。 根据RAPD 基因表型矩阵, 通过UPGMA法进行不同白桂木种群的聚类分析, 得到白桂木个体间的树状图(图4), 进一步反映了白桂木种群间的亲缘关系。 从聚类结果可看出, 聚类结果与该种群所处的地理纬度位置比较一致, 地理位置相对最近的梅列种群和三元种群先聚在一起, 然后是清流种群和永安种群, 最后是地理位置最远的永春种群。
3 讨论
通常认为, 濒危物种因种群缩小而导致近亲繁殖和等位基因的随机固定, 从而降低了其种群生存能力, 遗传多样性水平较低, 如华木莲Sinomanglietia glauca 和银杉等[9,19], 而黄久香等[20]和恽锐等[21]对观光木Tsoongiodendron odorum 和辽东栎Quercus liaotungensis 的研究发现有些濒危物种也保持持着较高的遗传多样性水平。 白桂木遗传多样性水平的分析结果表明: 白桂木种群具有较高的遗传多样性, 总的多态位点频率为81.08%, 与观光木(84.62%)和辽东栎(79.00%)相当, 远高于银杉(32.00%)和华木莲(53.22%)。 平均Shannon指数为0.231 9, Nei 指数为0.166 0, 白桂木的遗传分化系数 (GST)为0.737 8, 基因流 (Nm)为0.125 1。 说明白桂木种群间的基因流受限严重,其遗传变异有73.78%存在于种群间, 只有26.22%发生于种群内, 可见白桂木种群间的分化程度较高, 种群内的分化程度较低, 这与濒危植物冷杉相似。
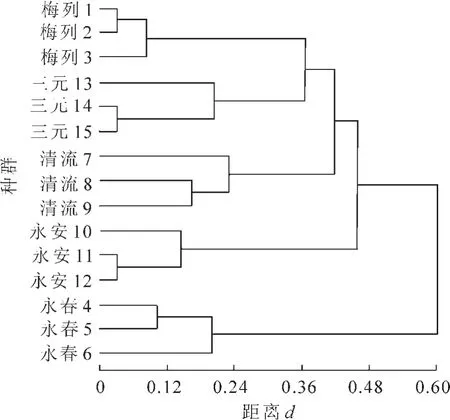
图4 不同白桂木种群的UPGMA 聚类图Figure 4 UPGMA dendrogram for populations of Artocarpus hypargyreus
通过UPGMA 聚类分析表明: 不同白桂木种群的聚类结果与其地理纬度位置基本一致。 地理位置最近的先聚一起, 其次相对较近的聚在一起, 最后是距离最远的。 可见不同白桂木种群间的基因交流少, 小种群分化严重, 种群内杂合度低,适应性弱, 是其种群衰退的重要原因。 同时, 白桂木的物种具有较高的遗传多样性, 因此在就地保护基础上, 更要注意迁地保护。 由于其种群间基因流很小, 遗传分化强烈, 应注意多种群采种, 进行人工繁殖, 尤其应在遗传多样性比较高的种群中采种, 采用扦插和组织培养等快繁途径来扩大和更新现有种群。 同时通过打破白桂木种子休眠, 创造条件提高白桂木种子发芽率, 在较大的空间范围进行播种, 增加该种群的幼苗数量, 创造基因交流和重组的条件, 进一步保护白桂木的遗传多样性。
[1] 傅立国. 中国植物红皮书[M]. 北京: 北京科学技术出版社, 1992: 98.
[2] 甘茂罗, 欧阳胜, 谢平, 等. 白桂木的化学成分研究[J]. 中草药, 2005, 36 (7): 988-989.GAN Maoluo, OUYANG Sheng, XIE Ping, et al. Study on chemical composition of Artocarpus hypargyreus [J].Chin Tradit Herbal Drugs, 2005, 36 (7): 988-989.
[3] 陈黎明, 谢平, 肖庆青, 等. 白桂木化学成分研究[J]. 中草药, 2007, 38 (6): 815-818.CHEN liming, XIE Ping, XIAO Qingqing, et al.Study on chemical composition of Artocarpus hypargyreus [J]. Chin Tradit Herbal Drugs, 2007, 38 (6): 815-818.
[4] 周德义, 杨尔滨, 邓勇, 等. 白桂木凝集素的纯化与性质的研究[J]. 生物化学与生物物理学报, 1995, 17 (1): 61- 66.ZHOU Deyi, YANG Erbin, DENG Yong, et al. Purification and characterization of lectin from the seeds of Artocarpus hypargyreus Hance [J]. Acta Biochem Biophys Sin, 1995, 17 (1): 61-66.
[5] 吴耀生, 张红, 周素芳, 等. 酶标白桂木凝集素糖蛋白结合特性的分析[J]. 中国生物化学与分子生物学报, 2000,16 (2): 210-214.WU Yaosheng, ZHANG Hong, ZHOU Sufang, et al. Characterization of Artocarpus hypargyreus Hance lectin interacting with glycoproteins [J]. Chin J Biochem Mol Biol, 2000, 16 (2): 210-214.
[6] 邓勇, 周德义, 关祺芳, 等.红桂木、 白桂木、 木菠萝种子凝集素生物学性质的比较分析[J].广西医科大学学报, 1995,12 (2): 183-185.DENG Yong, ZHOU Deyi, GUAN Qifang, et al. Biology characteristic analysis of seed agglutinin between Artocarpus lignanensis Merr, Artocarpus hypargyreus Hance and Artocarpus heterophyllus Lam. [J]. J Guangxi Med Univ, 1995, 12(2): 183-185.
[7] 陈开团, 张宗华, 蒋延生.福建省25 种珍贵树种的分布现状与保护对策的初步研究[J]. 福建林业科技, 2003, 30 (2):70-73.CHEN Kaituan, ZHANG Zhonghua, JIANG Yansheng. Preliminary on the distribution status and protective countermeasures of rare tree species in Fujian Province [J]. J Fujian For Sci Technol, 2003, 30 (2): 70-73.
[8] 祖元刚, 张文辉, 阎秀峰, 等. 濒危植物裂叶沙参保护生物学[M]. 北京: 科学出版社, 1999: 182-223.
[9] 葛颂, 王海群, 张大明. 八面山银杉林的遗传多样性和群体分化[J]. 植物学报, 1997, 39 (3): 266-271.GE Song, WANG Haiqun, ZHANG Daming. Genetic diversity and population differentiation of Cathaya argyrophylla Chun et Kuang in Bamian Mountain [J]. Chin Bull Bot, 1997, 39 (3): 266-271.
[10] 蒋志刚, 马克平, 韩兴国. 保护生物学[M]. 杭州: 浙江科学技术出版社, 1997.
[11] 汪小全, 邹喻苹, 张大明, 等. RAPD 应用于遗传多样性和系统学研究中的问题[J]. 植物学报, 1996, 38 (12):954-962 WANG Xiaoquan, ZOU Yuping, ZHANG Daming, et al. Problems in the use of RAPD to the study of genetic diversity and systematics [J]. Chin Bull Bot, 1996, 38 (12): 954-962.
[12] 王关林, 方宏筠. 植物基因工程[M]. 北京: 科学出版社, 2002: 744.
[13] NEI M, LI W H. Mathematical model for studying genetic variation in terms of restriction endonucleases [J]. Proc Natl Acad Sci USA, 1979, 76: 5269-5273.
[14] 罗美娟, 叶功富, 卢昌义. 短枝木麻黄群体的遗传分化和遗传结构[J]. 福建林学院学报, 2007, 27 (4): 343-348.LUO Meijuan, YE Gongfu, LU Cangyi. Genetic differentiation and structure of Casuarina equisetifolia populations [J]. J Fujian Coll For, 2007, 27 (4): 343-348.
[15] 宋丛文, 包满珠. 天然珙桐群体的RAPD 标记遗传多样性研究[J]. 林业科学, 2004, 40 (4): 75-79.SONG Congwen, BAO Manzhu. Study on genetic diversity of RAPD mark for natural Davidia involucrata population [J].Sci Silv Sin, 2004, 40 (4): 75-79.
[16] 苏何玲, 唐绍清. 濒危植物资源冷杉遗传多样性研究[J]. 广西植物, 2004, 24 (5): 414-417.SU Heling, TANG Shaoqing. Genetic diversity of the endangered plant Abies ziyuanensis in two populations [J]. Guihaia, 2004, 24 (5): 414-417.
[17] WRIGHT S. Evolution in Mendelian population [J]. Genetics, 1931, 16: 97-159.
[18] NEBAUER S G, CASTILLO-AGUDO L, SEGURA J, et al. RAPD variation within and among natural populations of outcrossing willow-leaved foxglove (Digitalis obscurea L.)[J]. Thror Appl Genet, 1999, 98: 985-994.
[19] 林新春, 俞志雄, 裘利洪, 等. 濒危植物华木莲的遗传多样性研究[J]. 江西农业大学学报: 自然科学版, 2003, 25(6): 805-810.LIN Xinchun, YU Zhixiong, QIU Lihong, et al. Studies on genetic diversity of endangered Sinomanglietia glauca(Magnoliaceae)[J]. Acta Agric Univ Jiangxi, 2003, 25 (6): 805-810.
[20] 黄久香, 庄雪影. 华南三地观光木遗传多样性的RAPD 分析[J]. 华南农业大学学报, 2002, 23 (2): 54-57.HUANG Jiuxiang, ZHUANG Xueying. Comparison of genetic diversity of Tsoongiodendron odorum in southern China by RAPD markers [J]. J South China Agric Univ, 2002, 23 (2): 54-57.
[21] 恽锐, 钟敏, 王洪新, 等. 北京东灵山辽东栎种群DNA 多样性的研究[J]. 植物学报, 1998, 40 (2): 169-175.YUN Rui, ZHONG Ming, WANG Hongxin, et al. Study on DNA diversity of Quercus liaotungensis Koidz population[J]. Chin Bull Bot, 1998, 40 (2): 169-175.
猜你喜欢
今日农业(2022年15期)2022-09-20
清华金融评论(2022年4期)2022-04-13
湖南电力(2021年1期)2021-04-13
国际放射医学核医学杂志(2021年10期)2021-02-28
房地产导刊(2020年7期)2020-08-24
新世纪智能(高一语文)(2020年12期)2020-06-01
红土地(2018年7期)2018-09-26
科技视界(2016年27期)2017-03-14
中国医药导报(2015年27期)2015-02-28
中国药理学通报(2014年2期)2014-05-09
