
内蒙古典型草原小叶锦鸡儿灌丛地上净初级生产力和种群结构对火烧的响应
2010-06-08 07:52林燕白永飞
草业学报 2010年5期
林燕,白永飞
(中国科学院植物研究所植被与环境变化国家重点实验室,北京100093)
近一个世纪以来,全球范围内许多温带及稀树草原均发生了不同程度的灌丛化,群落中木本植物的密度不断增加,而草本植物的盖度和生产力不断降低,进而导致了生态系统服务功能的衰退[1-5]。草原灌丛化的因素很多,过度放牧、火烧频率降低、气候变暖、CO2浓度升高、灌木种子的传播,以及草本植物与灌木之间的竞争关系的改变等都可能造成灌丛化的发生[2,6,7]。而对于许多地区,导致草原灌丛化大面积发生的最直接因素是长期的过度放牧[8]。过度放牧使草本植物的生物量降低,减少了可燃性物质的数量,降低了火烧的频率和强度,进而加速了灌木对草原群落的入侵[9-11]。火烧被抑制后,群落中对火烧敏感的灌木幼苗则易于定居[12,13]。同时,由于灌木具有较发达的根系,能够利用更深层的土壤水分,使其比草本植物具有更强的竞争力,特别是干旱和过度放牧的交互作用进一步加速了木本植物对草原群落的入侵、定居和扩展,进而导致了灌丛化的大面积发生[14]。
内蒙古锡林河流域位于内蒙古高原中东部,是我国温带典型草原的核心分布区。该地区的代表性植被类型为羊草(Leymuschinensis)草原和大针茅(Stipagrandis)草原,旱生豆科灌木小叶锦鸡儿(Caraganamicrophylla)是这两类草原群落的伴生种之一[15]。自20世纪80年代以来,由于长期过度放牧,草原植被退化十分严重,草原火烧频率显著降低,导致小叶锦鸡儿灌丛大面积扩张,草原灌丛化现象不断加剧,进而对生态系统结构与功能产生了重要影响[16-20]。目前,关于小叶锦鸡儿种群的研究主要集中于自身生长和生理特征[21-25],以及对土壤等环境因子的影响[26,27],而对小叶锦鸡儿在草原中发生大面积扩张的原因及其控制因素的研究则相对薄弱。因此,开展火烧对草原群落灌丛地上净初级生产力和种群结构影响的研究,对于退化草地的恢复和合理利用,保护生态环境,实现畜牧业的可持续发展具有重要的理论和实践意义。
本研究以内蒙古锡林河流域退化草原群落中大面积生长的小叶锦鸡儿灌丛为研究对象,通过人为控制的火烧实验,研究小叶锦鸡儿地上净初级生产力和生物量分配对火烧的响应,探讨不同大小的小叶锦鸡儿株丛对火烧的敏感性,为进一步研究火烧对其种群动态的调控作用,制定合理的草原火管理对策提供科学依据。
1 材料与方法
1.1 实验地概况
本研究是在中国科学院内蒙古草原生态系统定位研究站进行的。该站位于内蒙古锡林河流域,属温带半干旱大陆性季风气候,冬季寒冷干燥、夏季温暖湿润,年平均气温为1.3℃,其中,1月平均气温为-21.0℃,7月平均气温为19.0℃,多年平均降水量为340.9 mm,主要集中于6-8月份,占全年降水量的60%~80%。植物生长季开始于4月末,结束于10月初,持续时间大约150 d[28]。
火烧实验在中国科学院内蒙古草原生态系统定位研究站的退化恢复样地(43°37′N,116°40′E)内进行。该样地位于锡林河二级阶地和丘陵坡麓之间,海拔1 187 m,地势微倾斜,地表较平整,样地于1983年围封以来没有家畜放牧采食[29]。土壤为暗栗钙土,质地属沙壤土,0~20 cm土层土壤颗粒组成为砂(0.05~2.0 mm)占81.06%、粉砂(0.002~0.05 mm)占11.29%、粘粒(<0.002 mm)占7.65%。土壤有机碳含量为1.22%,全氮含量为0.12%,全磷含量为0.02%。样地内的原生植被为羊草+大针茅草原,由于长期过度放牧,围封前已退化为小叶锦鸡儿+冷蒿(Artemisiafrigida)+丛生小禾草群落[29]。群落中常见植物种约50余种,主要包括:小叶锦鸡儿、羊草、大针茅、糙隐子草(Cleistogenessquarrosa)、冰草(Agropyroncristatum)、冷蒿、木地肤(Kochiaprostrata)、野韭(Alliumramosum)、菊叶委陵菜(Potentillatanacetifolia)、寸草苔(Carexduriuscula)、细叶白头翁(Pulsatillaturczaninovii)、猪毛菜(Salsolacollina)、轴藜(Axyrisamaranthoides)等。
1.2 实验设计
2005年,在实验区域内选择地势平坦、植被均匀且有代表性的地段,采用随机区组设计,将其划分为9个区组,每个区组中包含81个小区,每个小区面积为10 m×10 m;各区组间的间隔为3 m,各小区间过道为1 m,作为火烧隔离带。
2006年4月31日,在实验样地内进行了第1次火烧;2007年5月3日,对2006年火烧过的小区进行了第2次火烧。以2005年的本底调查为基础和此后连续2年的火烧实验数据,本实验拟分析和探讨第1次火烧处理和连续2年火烧对小叶锦鸡儿种群组成、株丛总地上生物量、老枝、当年枝和叶片生物量的影响。
1.3 小叶锦鸡儿灌丛大小及其分布特征
2005年8月,即火烧实验的前一年,调查了样地中所有小叶锦鸡儿灌丛的空间分布状况和生物学特性,为进一步研究小叶锦鸡儿灌丛对火烧处理的响应提供本底数据。在2006和2007年8月,分别对火烧后上述小叶锦鸡儿灌丛的形态学性状进行了连续2年的测定,用于对每一灌丛进行火烧处理前后的比较研究。具体方法为:在每个10 m×10 m的小区内,对所有的小叶锦鸡儿灌丛标号并定位,即以各小区的西北角作为坐标原点,测量灌丛中心距小区西边(X)和北边(Y)的垂直距离,并将小区内所有灌丛的分布按比例绘制于坐标图上。在记录灌丛坐标的同时,测量灌丛的形态学性状,具体测定指标包括:灌丛高度(H,height),即大部分枝条所处的平均高度;灌丛的最大直径(Max)以及与其垂直的最小直径(Min);以地面为基准的灌丛的枝条数(SN,stem number)。本底调查最终记录的小叶锦鸡儿灌丛数为6 588株。
1.4 小叶锦鸡儿净地上生物量的测定
2006年9月初,正值灌丛地上生物量的高峰期,在可用于刈割的小区内选择未经火烧与火烧处理的小区中进行小叶锦鸡儿灌丛生物量的测定,并建立生物量估测模型。具体测定方法为:在每个10 m×10 m的小区内,根据2005年绘制的坐标图对其中的小叶锦鸡儿灌丛逐个进行生物量的测定。记录完毕后将样方内出现的所有的小叶锦鸡儿灌丛齐地面剪下,带回室内,在65℃下烘干至恒重后,测量其老枝生物量、当年枝生物量、叶片生物量和果实生物量。根据小叶锦鸡儿灌丛体积及枝条数测定结果,建立了小叶锦鸡儿灌丛生物量最佳估测模型,用以估算非破坏性取样株丛的总生物量、老枝生物量、当年枝生物量和叶片生物量。在株丛水平上,当年枝和叶片生物量的合计为株丛的当年净地上生物量。在群落水平上,单位面积内所有株丛净地上生物量的合计为小叶锦鸡儿的净地上初级生产力。
1.5 统计分析
利用SPSS Version 13.0软件,对株丛总地上生物量、老枝生物量、当年枝生物量和叶片生物量的估算采用多元回归分析的方法。不同火烧处理间地上净初级生产力、株丛高度、枝条密度、茎生物量、叶片生物量,以及不同大小株丛净地上生物量间的差异,采用单因素方差分析(One-way ANOVA),多重比较采用Duncan新复极差法(shortest significant ranges,SSR)。
2 结果与分析
2.1 株丛总地上生物量、老枝、当年枝和叶片生物量与株丛大小和枝条数之间的关系
根据未经火烧处理和火烧处理后小叶锦鸡儿灌丛地上总生物量、老枝、当年枝和叶片生物量,以及株丛高度,株丛最大直径、最小直径和枝条数等形态学性状的实测数据,建立了株丛总生物量、老枝、当年枝和叶片生物量与各形态学性状之间关系的多元回归模型。统计分析结果表明,各形态学性状中,株丛总体积(CV,plant volume)和枝条数(SN)与其地上生物量(Y)之间存在着显著的线性正相关或对数线性相关关系,这2个变量可以解释57%~89%的方差。对于未经火烧处理的小叶锦鸡儿,其株丛总地上生物量、老枝和叶片生物量的估测精度均在81%以上,当年枝生物量的估测精度则相对较低(表1)。对于经过火烧处理的小叶锦鸡儿,其株丛总地上生物量、当年枝和叶片生物量均与株丛总体积和枝条数存在着极显著的对数正相关关系,株丛总体积和枝条数2个变量可以解释82%以上的方差(表2)。
2.2 火烧处理前不同大小株丛的频率分布
2005年实验区内本底调查测定的小叶锦鸡儿株丛共计为6 588株,其冠幅(最大直径与最小直径的平均值)的频率分布呈对数曲线分布(图1)。不同小叶锦鸡儿株丛个体之间存在着较大的差异,冠幅的变化范围为5~925 cm。总体而言,约80%的小叶锦鸡儿株丛其冠幅均小于150 cm。在所有调查统计的株丛中,冠幅为0~50 cm的株丛占30%、冠幅为50~150 cm的株丛占49%、冠幅大于150 cm的株丛占21%。这3个等级的株丛基本上呈正态分布,能够较好地反映实验区内小叶锦鸡儿株丛的实际分布情况,因此,采用这3个等级作为小叶锦鸡儿株丛个体大小的分组依据。

表1 未经火烧处理的小叶锦鸡儿株丛地上总生物量、枝条和叶片生物量最佳估测模型Table 1 The best fit equations for estimating the total aboveground biomass,stem biomass,and foliar biomass of C.microphylla before prescribed burning

表2 火烧处理后小叶锦鸡儿株丛地上总生物量、枝条和叶片生物量最佳估测模型Table 2 The best fit equations for estimating the total aboveground biomass,stem biomass,and foliage biomass of C.microphylla after prescribed burning

图1 不同大小的小叶锦鸡儿株丛频率分布 (n=6 588)Fig.1 The frequency distribution of C.microphylla with different crown size(n=6 588)
2.3 火烧对种群净初级生产力的影响
火烧处理对小叶锦鸡儿灌丛的生长具有显著的抑制作用,火烧显著地降低了群落中小叶锦鸡儿灌丛的地上净初级生产力(ANPP,annual aboveground net primary productivity)(图2A)。经过2006年第1年火烧和2007年连续2年火烧处理的小区,小叶锦鸡儿的地上净初级生产力均显著低于未经火烧处理的小区。ANPP与2005年对照相比,分别下降了22%和30%(P<0.05)。对年际间气候条件的影响进行校正后发现,火烧当年和连续2年火烧使小叶锦鸡儿的地上净初级生产力分别比火烧前下降了27%和66%(P<0.05)(图2B)。同时,连续2年火烧处理使小叶锦鸡儿的地上净初级生产力进一步降低,同火烧第1年相比,地上净初级生产力降低了53%。
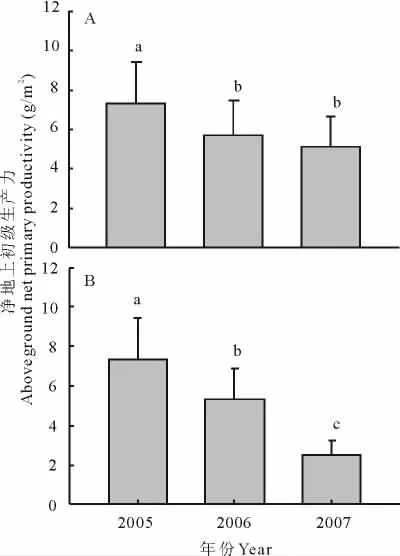
图2 火烧对小叶锦鸡儿地上净初级生产力的影响(mean±SE,n=9)Fig.2 The patterns of aboveground net primary productivity(ANPP)of C.microphylla in different burning treatments(mean±SE,n=9)
2.4 火烧对枝条密度、高度、茎叶生物量及其分配的影响
火烧处理几乎使小叶锦鸡儿的所有多年生老枝死亡,但刺激了休眠芽的再生,产生了大量当年生枝条。2006年火烧第1年,小叶锦鸡儿的新生枝条密度增加了约63%(P<0.05)。2007年经过连续2年火烧后,枝条数与火烧第1年相比,没有发生显著变化(P>0.05),但仍然显著高于对照(图3A)。同枝条密度的变化相反,火烧使小叶锦鸡儿灌丛的平均高度显著降低,株丛平均高度下降了7 cm(图3B)。同时,与2006年火烧第1年相比,连续2年火烧后,株丛高度又进一步下降了7 cm(图3B)。
火烧显著地提高了小叶锦鸡儿灌丛的当年枝生物量(图4A)。与对照相比,火烧第1年和连续2年火烧使小叶锦鸡儿的当年枝生物量分别增加了约1.5和1.3倍(P<0.05)。但是,火烧显著地降低了小叶锦鸡儿的叶片生物量,在火烧第1年和连续2年火烧后,其叶片生物量的下降幅度分别为42%和48%(图4B)。
小叶锦鸡儿灌丛地上净初级生产力是由当年生枝条、叶片和果实生物量三部分构成的。在没有进行火烧处理的小区(对照),当年生枝条的平均生物量约占地上总生物量的15%,叶片生物量占66%,果实生物量占19%,茎叶比(当年生枝条生物量与叶片生物量之比)为1∶4.4。经过火烧处理的小区,小叶锦鸡儿灌丛不能开花结实,地上净初级生产力均由当年生枝条和叶片生物量组成,两者所占比例分别为52%和48%,茎叶比为1∶1.1。这表明,火烧处理显著地增加了茎的生物量分配,减少了叶的生物量分配,改变了小叶锦鸡儿的茎叶比。
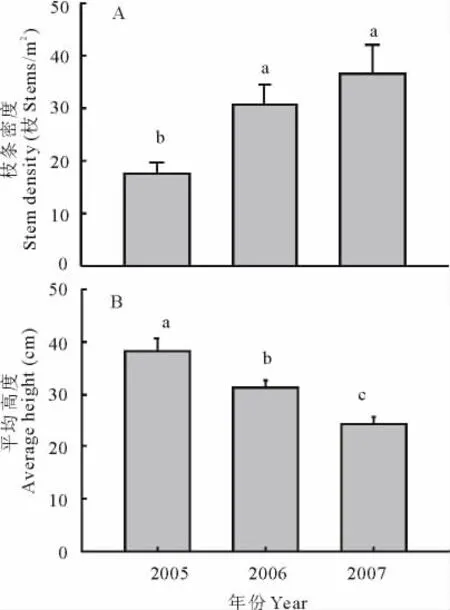
图3 火烧对小叶锦鸡儿枝条密度和高度的影响Fig.3 The effects of prescribed burning on stem density and height of C.microphylla
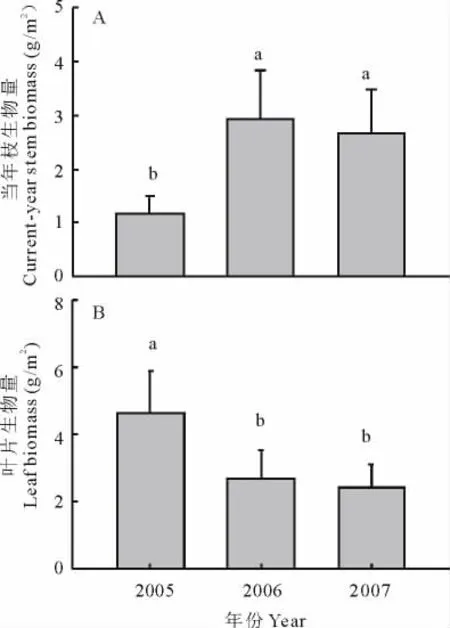
图4 火烧对小叶锦鸡儿当年枝和叶片生物量的影响Fig.4 The effects of prescribed burning on current-year stem and leaf biomass of C.microphylla
2.5 火烧对个体地上生物量的影响
在株丛水平上,经过2006年第1年火烧和2007年连续2年火烧处理,3个冠幅组小叶锦鸡儿个体的净地上生物量均显著地低于对照(P<0.05),这与群落水平上小叶锦鸡儿初级生产力的变化趋势是一致的(图5)。
3个不同冠幅组的小叶锦鸡儿个体净地上生物量的相对变化表明,同火烧前的本底值相比,连续2年火烧处理对冠幅为0~50 cm的小灌丛影响最大,株丛平均净地上生物量比火烧前下降了60%,下降幅度显著高于其他2个冠幅组(P<0.05,图6)。同时,第1年火烧处理与连续2年火烧表现出相同的变化趋势,即0~50 cm的小灌丛受火烧的影响最大,地上净初级生产力比火烧前降低了57%,明显高于其他2个冠幅组(P<0.05)。但第1年火烧和连续2年火烧处理的结果相比,3个冠幅组地上净初级生产力的下降幅度间差异并不显著(P>0.05)。
3 讨论
3.1 小叶锦鸡儿初级生产力、枝条密度、高度和生物量分配对火烧的响应
周期性火烧、干旱和放牧是全世界范围内草原生态系统维持的重要因子[5,30-32]。历史上,在蒙古高原的大部分草原地区,每隔3~5年就经历一次周期性的火烧。火烧曾作为草原管理的手段之一,用来有效地控制木本植物的入侵和大量繁殖,剔除凋落物的积累,因而对草原生态系统结构与功能稳定性的维持具有重要作用[33]。由于草本植物的分生组织一般位于地下的根茎分蘖处,而灌木的分生组织则位于地上叶腋和茎的交叉处,因此灌木比草本植物更容易遭受火烧的影响[34]。灌木个体在经历不同强度的火烧后有2种反应:死亡或者萌发。Heisler等[5]连续18年的长期研究结果表明,高频率的火烧可以有效地阻止灌木物种进入草原,但是侵入的灌木物种一旦定居后,即使高频率的火烧也不能将它们完全消除。火烧可以有效地阻止木本植物进一步入侵到草原群落中,但很难将木本植物占优势的草原群落恢复至原来的面貌,因为火烧后木本植物会迅速地通过发芽再生[35,36]。例如,灌木物种可以通过种子库中储存的种子萌发,产生新的幼苗,或者从火烧中存活的灌丛根部或枝条处再次萌发出新的枝条,适应火烧后的环境[37]。本研究中,火烧处理后灌木的死亡率为0,所有的灌丛均通过由根部或枝条处萌发产生新的枝条,2006年第1年火烧和2007年连续2年火烧均使群落水平和株丛水平上小叶锦鸡儿的地上净生物量显著降低。这说明在内蒙古典型草原群落中,短期的火烧对小叶锦鸡儿灌丛的进一步扩张具有明显的抑制作用,但从长期来看,是否可以通过控制性火烧的手段将内蒙古草原恢复到原来的草原群落,仍需进一步的深入研究。
Heisler等[34]研究发现,经过2年的火烧处理,粗叶梾木(Cornusdrummondii)灌丛枝条数增加了近6倍。在内蒙古草原,火烧处理显著地增加了小叶锦鸡儿的枝条密度,但使其平均高度显著降低。与对照相比,火烧第1年和连续2年火烧使小叶锦鸡儿的新生枝条密度分别增加了72%和95%,使其株丛平均高度分别下降了7和14 cm。同时,火烧改变了小叶锦鸡儿的生物量分配,显著地增加了茎的生物量分配,减少了叶的生物量分配,改变了小叶锦鸡儿的茎叶比。火烧第1年和连续2年火烧使小叶锦鸡儿地上生物量的生殖器官分配为0,与对照相比,当年枝生物量分别增加了约1.5和1.3倍,但叶片生物量分别下降了42%和48%,茎叶比由对照的1∶4.4升高为1∶1.1。这表明,尽管火烧处理使小叶锦鸡儿的枝条密度和当年枝生物量增加,但火烧显著地削弱了小叶锦鸡儿的生活力,因而对小叶锦鸡儿的进一步扩展具有明显的控制作用。
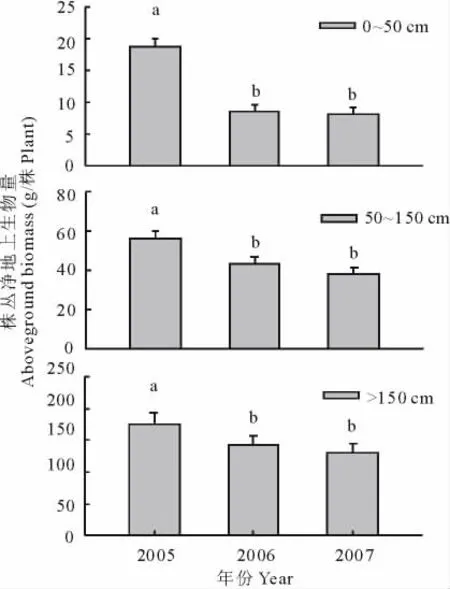
图5 火烧对小叶锦鸡儿不同大小株丛地上净生物量的影响(mean±SE)Fig.5 The effects of burning on aboveground net biomass of C.microphylla with different crown size(mean±SE)
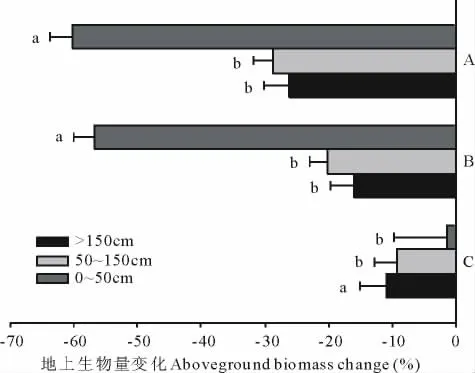
图6 小叶锦鸡儿株丛净地上生物量变化对不同火烧处理的响应 (mean±SE)Fig.6 Aboveground biomass change of different brown classes under different burning regimes(mean±SE)
3.2 不同大小的小叶锦鸡儿株丛对火烧的响应
火烧后,灌木需要利用大量火烧前所储存的能量来维持其生存和生长[37,38],灌木根部是储存能量的重要器官,因此,火烧前灌丛根部营养物质积累的多少直接影响火烧后灌木的再生[39-41]。有研究表明,灌木根部营养物质的积累与灌丛大小成正比,灌丛越小积累的能量越少,其在火烧后的再生能力就越弱,受火烧的影响就越大[42]。本研究结果表明,0~50 cm的小灌丛在1年火烧和连续2年火烧后,地上净初级生产力比未火烧时分别减少了57%和60%,明显高于50~150和>150 cm 2组灌丛,表明其受火烧的影响最为明显。
此外,由于在没有火烧抑制时,灌木随着个体的生长逐渐形成大小不同的“资源岛”(resource island)[43],与较大的灌木株丛相比,草本植物更容易生长于小灌丛的内部[34],能够为火烧提供更多的燃料,从而使得小灌丛比大灌丛更容易受火烧的影响。另外,随着围封时间的延长,凋落物量增加[44-46],通过火烧能够去除地表凋落物、提高土壤温度、增加草地表面的空斑面积,从而提高草种种苗的出苗和存活率[47],最终使草本植物多度和初级生产力增加,进一步增强草本植物的竞争力。与较大的灌丛相比,小灌丛的竞争能力较弱,因此,与草本植物之间的竞争作用也是小灌丛对火烧更敏感的原因之一。
3.3 火烧对小叶锦鸡儿种群动态的长期影响
Archibald和Bond[48]研究发现,湿润和干旱地区灌木物种的生长形态不同。湿润地区,光是植物生长的限制因子,因此,灌丛通常选择纵向生长,以增加高度来获取更多的光资源。而在干旱半干旱地区,水分是植物生长的主要限制因子,所以灌丛往往由基部的分枝较多,通过横向生长以增加冠幅来获取地表更多的水分。一般情况下,湿润地区的灌木随着年龄的增长,高度的增长相对较快,而干旱半干旱地区的灌木则随着年龄的增长,冠幅的增长相对较快。目前,已经有很多研究采用植物的形态特征作为判断其年龄的依据[37,42,48,49]。本研究中,小叶锦鸡儿是内蒙古典型草原中广泛分布的一种多枝条灌木物种,枝条的再生能力较强,侧根随着年龄的增长向水平方向生长。因此,可以利用灌丛冠幅大小作为判断其年龄的依据。
通过分析不同大小的小叶锦鸡儿株丛在整个种群中所占的比例,本研究表明,在内蒙古锡林河流域小叶锦鸡儿种群属于增长型的种群,中、幼龄个体的比例较大,而老龄个体的比例相对较少。这表明在未来几年内,若不通过有效的管理措施加以控制,该种群还将继续保持不断增长的趋势。火烧处理后,灌丛的幼年个体受火烧的影响最为明显。因此,利用火烧管理该草原,可以有效地控制小叶锦鸡儿种群的进一步增长,增强草本植物的竞争力。特别是采用火烧、施肥和割草等管理措施相结合的手段,有效地控制草原灌丛化的不断扩展,提高草原的生产力水平,恢复退化的草原群落和草原生态系统的生态服务功能,实现畜牧业生产的可持续发展。
[1]Schlesinger W H,Reynolds J F,Cunningham G L,etal.Biological feedbacks in global desertification[J].Science,1990,247:1043-1048.
[2]Knight C L,Briggs J M,Nellis M D.Expansion of gallery forest on Konza Prairie Research Natural Area,Kansas,USA[J].Landscape Ecology,1994,9:117-125.
[3]Archer S.Tree-grass dynamics in aProsopisthornscrubsavanna parkland:Reconstructing the past and predicting the future[J].Ecoscience,1995,2:83-99.
[4]Briggs J M,Knapp A K,Brock B L.Expansion of woody plants in tallgrass prairie:A fifteen-year study of fire and fire-grazing interactions[J].American Midland Naturalist,2002,147:287-294.
[5]Heisler J L,Briggs J M,Knapp A K.Long-term patterns of shrub expansion in a C4-dominated grassland:Fire frequency and the dynamics of shrub cover and abundance[J].American Journal of Botany,2003,90:423-428.
[6]Scholes R J,Archer S R.Tree-grass interactions in savannas[J].Annual Review of Ecology and Systematics,1997,28:517-544.
[7]Sturm M,Racine C,Tape K.Climate change:Increasing shrub abundance in the Arctic[J].Nature,2001,411:546-547.
[8]Briggs J M,Knapp A K,Blair J M,etal.An ecosystem in transition:Causes and consequences of the conversion of mesic grassland to shrubland[J].Bioscience,2005,55:243-254.
[9]Wright H A.The role and use of fire in the semi-desert grass-shrub type[A].General Technical Report[C].Ogden,Utah:USDA Forest Service,1980:85.
[10]Pyke G H.Optimal foraging theory:A critical review[J].Annual Review of Ecology and Systematics,1984,15:523-575.
[11]Bahre C J.A Legacy of Change:Historic Human Impact on Vegetation of the Arizona Borderlands[M].Tucson:University of Arizona Press,1991:231.
[12]Martin S C.Responses of semi-desert grasses and shrubs to fall burning[J].Journal of Range Management,1983,36:604-610.
[13]McPherson G R.The role of fire in the desert grassland[J].Renewable Energy Foundation,1995,110:131-151.
[14]Walter H.Ecology of Tropical and Subtropical Vegetation[M].Edinburgh:Oliver and Boyd,1971:207-236.
[15]Bai Y F,Han X G,Wu J G,etal.Ecosystem stability and compensatory effects in the Inner Mongolia grassland[J].Nature,2004,431:181-184.
[16]李博,雍世鹏,李忠厚.锡林河流域植被及其利用[A].草原生态系统研究(第三集)[M].北京:科学出版社,1988:84-183.
[17]赵献英,杨彦臣,杨汝蒙.锡林河流域天然草场的生态地理特征及其展望[A].草原生态系统研究(第三集)[M].北京:科学出版社,1988:184-226.
[18]李永宏.放牧影响下羊草草原和大针茅草原植物多样性的变化[J].植物学报,1993,35(11):877-884.
[19]熊小刚,韩兴国,白永飞,等.锡林河流域草原小叶锦鸡儿分布增加的趋势、原因和结局[J].草业学报,2003,12(3):57-62.
[20]熊小刚,韩兴国.内蒙古半干旱草原灌丛化过程中小叶锦鸡儿引起的土壤碳、氮资源的空间异质性分布[J].生态学报,2005,25(7):1678-1683.
[21]孙志蓉,翟明普,王文全,等.密度对小叶锦鸡儿播种苗生长的影响[J].北京林业大学学报,2007,29(1):42-46.
[22]赵爱芬.差巴嘎蒿和小叶锦鸡儿根系分布及生长动态的初步研究[J].中国草地,1994,3:15-19.
[23]周海燕,李新荣,樊恒文,等.极端条件下几种锦鸡儿属灌木的生理特性[J].中国沙漠,2005,25(2):182-190.
[24]阿拉木萨,慈龙骏,杨晓晖,等.科尔沁沙地不同密度小叶锦鸡儿灌丛水量平和研究[J].应用生态学报,2006,17(1):31-35.
[25]陶林,高洪文,樊奋成.小叶锦鸡儿根瘤固氮活性的动态变化[J].中国草地,2005,27(3):53-56.
[26]刘方明,郝伟,姜勇.科尔沁沙地小叶锦鸡儿对土壤有机碳积累的影响[J].辽宁工程技术大学学报,2006,25(2):294-296.
[27]闫德仁,赵春光,齐凯,等.小叶锦鸡儿灌丛土壤肥力变化的研究[J].内蒙古林业科技,2007,33(3):1-4.
[28]姜恕.中国科学院内蒙古草原生态系统定位研究站的简历和研究工作概述[A].草原生态系统研究(第一集)[M].北京:科学出版社,1985:75-82.
[29]王炜,梁存柱,郝敦元,等.内蒙古草原退化群落恢复演替的研究(Ⅰ.退化草原的基本特征与恢复演替动力)[J].植物生态学报,1996,20(5):449-459.
[30]周道玮,周以良,郑焕能.火生态学研究评述[J].世界林业研究,1993,6:38-44.
[31]Holechek J,Pieper R D,Herbel C H.Range Management:Principles and Practices.Fifth edition[M].New Jersey:Pearson Prentice Hall,2002.
[32]Bond W J,Keeley J E.Fire as a global‘herbivore’:The ecology and evolution of flammable ecosystems[J].Trends in Ecology and Evolution,2005,20:387-394.
[33]Erdensaikhan N,Erdentuya M.Fire situation in Mongolia[J].International Forest Fire News,2002,26:75-83.
[34]Heisler J L,Briggs J M,Knapp A K,etal.Direct and indirect effects of fire on shrub density and aboveground productivity in a mesic grassland[J].Ecology,2004,85:2245-2257.
[35]Flinn M A,Wein R W.Depth of underground plant organs and theoretical survival during fire[J].Canadian Journal of Botany,1977,55:2550-2554.
[36]Keeley J E,Keeley M B,Bond W J.Stem demography and post-fire recruitment of a resprouting serotinous conifer[J].Journal of Vegetation Science,1999,10:69-76.
[37]Hodgkinson K C.Sprouting success of shrubs after fire:Height-dependent relationships for different strategies[J].Oecologia,1998,115:64-72.
[38]Bond W J,Midgley J J.Ecology of sprouting in woody plants:The persistence niche[J].Trends in Ecology and Evolution,2001,16:45-51.
[39]Pate J S,Froend R H,Bowen B J,etal.Seedling growth and storage characteristics of seeder and resprouter species of Med-iterranean-type ecosystems of south-west Australia[J].Annals of Botany,1990,65:585-601.
[40]Bell T L,Pate J S,Dixon K W.Relationships between fire response,morphology,root anatomy and starch distribution in south-west AustralianEpacridaceae[J].Annals of Botany,1996,77:357-364.
[41]Bell T L,Ojeda F.Underground starch storage in Erica species of the Cape floristic region:Differences between seeders and resprouters[J].New Phytologist,1999,144:143-152.
[42]McCarron J K,Knapp A K.C3shrub expansion in a C4grassland:Positive post-fire responses in resouces and shoot growth[J].American Journal of Botany,2003,90:1496-1501.
[43]Lett M S,Knapp A K.Consequences of shrub expansion in mesic grassland:Resource alterations and graminoid responses[J].Journal of Vegetation Science,2003,14:487-496.
[44]左万庆,王玉辉,王风玉,等.围栏封育措施对退化羊草草原植物群落特征影响研究[J].草业学报,2009,18(3):12-19.
[45]呼格吉勒图,杨劼,宝音陶格涛,等.不同干扰对典型草原群落物种多样性和生物量的影响[J].草业学报,2009,18(3):6-11.
[46]单贵莲,徐柱,宁发,等.围封年限对典型草原植被与土壤特征的影响[J].草业学报,2009,18(2):3-10.
[47]刘桂霞,龚玉梅,韩建国.草地空斑对种苗建植影响的研究进展[J].草业科学,2008,25(2):110-115.
[48]Archibald S,Bond W J.Growing tall vs growing wide:Tree architecture and allometry ofAcaciakarrooin forest,savanna,and arid environments[J].Oikos,2003,102:3-14.
[49]Meyer K M,Ward D,Moustakas A,etal.Big is not better:SmallAcaciamelliferashrubs are more vital after fire[J].African Journal of Ecology,2005,43:131-136.
猜你喜欢
水土保持学报(2022年5期)2022-10-10
种子(2022年5期)2022-06-27
西北农林科技大学学报(自然科学版)(2022年3期)2022-04-01
防护林科技(2020年5期)2020-07-21
水土保持研究(2020年1期)2020-04-27
南方周末(2019-06-27)2019-06-27
草业科学(2019年5期)2019-06-04
西藏农业科技(2018年4期)2018-04-25
小学生作文(低年级适用)(2018年12期)2018-03-23
莫愁(2017年15期)2017-05-11
