
陆地生态系统净初级生产力的时空动态模拟研究进展
2010-05-30 05:45:54夏文韬梁天刚
草业科学 2010年2期
王 莺,夏文韬,梁天刚
(兰州大学草地农业科技学院 农业部草地农业生态系统学重点实验室,甘肃 兰州730020)
植被净初级生产力(Net Primary Productivity,NPP)是指绿色植物在单位面积、单位时间内由光合作用所产生的有机物总量中扣除自养呼吸后的剩余部分,不仅直接反映了植被群落在自然环境条件下的生产能力以及陆地生态系统的质量状况,也是判定生态系统碳源/碳汇和调节生态过程的主要因子,在全球变化以及碳循环中扮演着重要的角色[1-2]。陆地生态系统净初级生产力是人类赖以生存与发展的物质基础,是人类生存所需的粮食、纤维和燃料的来源,其中大约40%的生产力被人类直接或间接的利用[3]。因此对植被NPP的研究有助于自然资源的有效管理和合理利用以及为制定相应的社会和经济发展战略提供依据。
人类对植被生产力的研究已有很长的历史,Lieth是陆地植被 NPP研究的先驱者之一[4],1973年他第一次综合而全面地对NPP的研究历史作了详细的阐述,并首次估算了全球NPP[5]。20世纪80年代初期,NPP的理论和方法研究在国际生物学计划(IBP)的推动下得到了迅速的发展。到了90年代,随着联合国政府间气候变化专门委员会(Intergovernmental Panel on Climate Change,IPCC)会议的召开以及国际地圈-生物圈计划(IGBP)核心项目——全球变化与陆地生态系统(GCTE)的实施[6],将实地测量数据与卫星遥感信息相结合,联系植物生理生态学过程和环境因子的NPP模型研究成为了热点。
1 NPP研究概况
1.1 站点实测NPP 早期NPP的研究都基于站点实测,主要包括直接收割法、光合作用测定法、CO2测定法、pH 值测定法、放射性标记测定法、叶绿素测定法和原料消耗量测定法等[7]。这些方法是用所选站点的代表植被的观测数据进行NPP计算,然后用数学方法推广到区域或全球尺度上。如胡自治等[8-9]在天祝草原试验站测量了1980-1981年2种原生高山草甸群落总 NPP值。杨福囤等[10]在海北高寒草甸生态系统定位站测量了1980-1984年的矮嵩草Kobresia humilis草甸地上地下部分NPP值。方精云等[11]利用森林蓄积量推算森林生物量和净生物量的方法,系统研究了我国森林植被的生物生产力。沈禹颖等[12]对河西走廊几种盐渍化草地的NPP进行了研究。由于这些数据是通过实测得到的,因此是目前重要的模型验证数据。而在区域或全球尺度上人们无法用实际测量的办法获得真实的NPP值,因此利用模型估算NPP就成为了一种重要的研究方法[13-14]。近年来,随着各种新技术的支持,全球NPP的测量及其模式研究有了很多发展与突破。
1.2 NPP模型研究 模型是人们对真实世界的抽象或简化,是人们认识和研究自然现象不可缺少的工具[15]。生产力模型有助于人们去了解生态系统的结构和功能以及它们对环境因子变化的反应。由于“粮食安全”和“全球变暖”这2个问题越来越被人们所重视,NPP建模及其应用都得到了飞速的发展[16-17],出现了一系列基于区域和全球尺度的NPP模型[14]。根据模型对各种调控因子的侧重点以及对NPP调控机理的解释,Cramer等[14]和Ruimy等[18]将这些模型概括为气候相关统计模型、生态系统过程模型和光能利用率模型。
1.2.1 气候相关统计模型(Statistical climate-correlation models) 气候相关统计模型也叫气候生产潜力模型。以 Miami模型[19]、Thornthwaite Memorial模型[20]和Chikugo模型[21]为代表。这类模型主要利用气候因子与NPP之间的相关性原理,利用大量的实测数据建立简单统计回归模型[4,22-23]。因此大部分统计模型获得的结果是潜在植被生产力。如刘明春等[24]使用逐步回归法将气温和降水作为预报因子得出牧草生物量气候模型。周广胜和张新时[26]以植被表面的二氧化碳通量方程(相当于NPP)与水汽通量方程(相当于蒸散发)之比确定的植被对水的利用效率为基础,根据水量平衡方程和热量平衡方程从能量与水分对蒸发影响的物理过程出发推导出了联系能量平衡方程和水量平衡方程的区域蒸散模式,并利用叶菲莫娃在国际生物学计划工作期间取得的23组世界各地植物的NPP数据和相应的气候要素建立了以植物生理生态特性为基础的自然植被的NPP模型。林慧龙等[26]利用≥0℃年积温(∑θ)和湿润度(K)指标,在周广胜和张新时模型的基础上,推导出∑θ和K指标与NPP间的关系,为进一步研究地带性草地类型的生产潜力、草地NPP的区域分布和全球分布提供了可能。这类模型的输入参数简单易得,可以对不同陆地生态系统NPP进行估算和预测[19,27]。缺点是缺乏严密的植物生理生态学机制,同时由于受到取样密度的影响,由点到面甚至向区域外推时产生的尺度转换问题会影响NPP的估测精度,不能很好地反映出空间格局的异质性[28]。而且模型使用的输入参数很简单,导致模型忽略了其他环境因子的作用以及异常气候条件对NPP的影响[29]。
1.2.2 生态系统过程模型(Ecosystem process models) 生态系统过程模型又称机理模型(Mechanistic ecophysiological models)或生物地球化学模型(Bio-geochemistry models)。该模型是通过对植物的光合作用、有机物分解及营养元素的循环等生理过程的模拟而得到的,可以与大气环流模式耦合,因此可以用这类模型进行NPP与全球气候变化之间的响应和反馈研究。因为其较强的机理性和系统性,所以该类模型的可靠性比较高,在不同条件下都可以详细地描述生物学过程。目前已有的生态系统过程模型很多,如BEPS[30]、TEM[31]、Forest-BGC[32]和 BIOMEBGC[33]。周才平等[34]利用TEM 模型,并嵌入了适时的卫星遥感数据(MODIS),对青藏高原区域NPP进行了估算,得到该地区2001年NPP(以碳累积量计)为302.44×1012g/a与实测数据结果非常相符。冯险峰[35]等利用BEPS模型估算了我国2001年陆地生态系统NPP。
但由于过程模型设计复杂,要求输入参数多,并且一些随物候期变化的变量,如叶面积指数及其他相关的参数随时空分布的定量化也很困难,因此在参数的可获得性、可靠性和尺度转换方面问题较多[36],导致在有些情况下需对模型进行简化或人为设定一些参数,限制了模型的精度。
1.2.3 光能利用率模型(Light utilization efficiency models) 光能利用率模型又被称为生产效率模型(Production efficiency models)。这类模型是以植物光合作用过程和Monteith[37-38]提出的光能利用率(ε)为基础建立的,它基于资源平衡观点[39]。Monteith通过对多种农作物生物量的实验发现植物NPP和可吸收的光合有效辐射(Ab-sorbed Photosynthetically Active Radiation,APAR)有线性正相关关系,因此他认为植被累积的生物量实际就是太阳入射辐射被植冠截获、吸收和转化的结果。近年来大量研究发现总初级生产力(Gross Primary Production,GPP)与APAR的线性关系更稳定,因为GPP的光能利用率ε受气候条件和环境因子的影响小,不同植被间的差异也很小[40]。主要的模型有GLO-PEM[40]、CASA[41]和C-FIX等[42]。
光能利用率模型主要有三大优点[44]:1)模型比较简单,参数可以直接通过遥感方式获得全覆盖数据,适于向区域及全球推广;2)冠层绿叶所吸收的光合有效辐射比可以由遥感数据计算得到,不需要繁杂的野外试验测定;3)适时性好,时空分辨率高,可以获得确切的NPP季节、年际动态。
尽管如此,该模型也存在一定的不确定性和不一致性[43]:1)太阳辐射与光合有效辐射的关系,光合有效辐射和植物可吸收利用的光合有效辐射的关系,不同植被的光能利用率,光合作用固碳与生物量的积累和分配的关系,这些均存在不确定性;2)基于植被指数的NPP模型无法实现在假设条件下(如气候变化、CO2波动、营养物质的变化)的模拟预测,因为这些信息不能通过遥感获取;3)该模型有这样一个假设:过去产生的归一化植被指数(Normalised Difference Vegetation Index,NDVI)与植被未来的潜在生产相关,遥感植被指数是植物生长的测量参数和驱动因子。但是在环境条件迅速变化的情况下(如火灾、大面积病虫鼠害等),由遥感获得的NDVI无法代表真实的地表植被信息,这时模型模拟的可靠性就比较小,而生态机理模型可能对这种短时间NPP变化的模拟具有优势。
2 遥感参数模型
随着遥感、地理信息系统和计算机技术的快速发展,它们被逐渐引入到了NPP的模型设计中,使得NPP的计算免去了很多复杂的实验工作。因为卫星遥感具有覆盖面积广、重复观测周期性高、时空分辨率多样、数据质量较稳定和对植被没有损害等特点,所以很适合于全球性的观测和研究。如NOAA(美国国家大气与海洋管理局)系列卫星上搭载的甚高分辨率辐射计AVHRR,由于其数据时空分辨率较高,时间序列长达20多年,所以该数据在NPP模型研究中应用最广。而1999年美国国家宇航局(NASA)发射的中分辨率成像光谱仪MODIS因为它的高时间分辨率和高光谱分辨率而在陆地生态系统NPP和碳循环研究中具有重要的应用价值。
近年来,利用遥感数据设计的NPP模型很多,本研究主要讨论CASA模型、C-FIX模型和BIOME-BGC模型。
2.1 CASA模型 CASA模型是Potter等[41]在1993年建立的光能利用率模型。1995年,Field[44]对CASA模型中的光能利用率等问题进行了深入讨论和改进。后来,Potter等[45]为了探讨包括人为活动导致的土地覆盖变化而对CASA模型及某些参数做了调整,改善与植被吸收需求有关的土壤碳循环和总生态系统氮量可行性的计算。
CASA模型允许参数随时间(t)和地点(x)的变化而变化,并通过与之对应的温度和水分条件对参数进行校正。模型中植被NPP的具体表达式为:
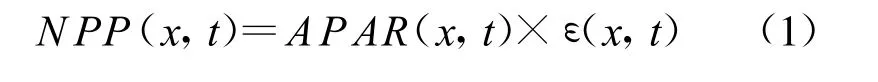
式中,x代表空间位置,t代表时间。NPP表示植被净第一性生产力;APAR表示植被所吸收的光合有效辐射;ε表示植被的光能转换率。
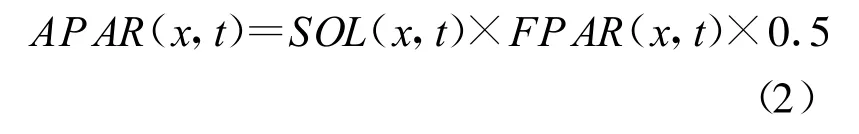
式中,SOL(x,t)是t时间象元x处的太阳总辐射(MJ/m2);FPAR(x,t)为植被层对入射光合有效辐射(PAR)的吸收比例;常数0.5表示植被所能利用的太阳有效辐射(波长为0.4~0.7 μ m)占太阳总辐射的比例。
Potter等[41]认为在理想条件下植被具有最大光能利用率,而在现实条件下光能利用率主要受温度和水分的影响,由式(3)表示:
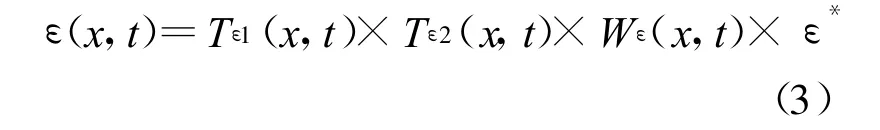
式中,Tε1反映在低温和高温时植物内在的生化作用对光合的限制而降低净第一性生产力,Tε 2表示环境温度从最适宜温度向高温和低温变化时植物的光能转化率逐渐变小的趋势[41,44];Wε是水分胁迫影响系数,反映水分条件的影响;ε*是理想条件下的最大光能转化率。Potter等[41]认为全球植被的ε*是 0.389(C,g/MJ),这也是CASA模型在运行全球NPP时用到的参数值。可是有研究表明这个值并不适合于中国的植被[46-47]。Running等[48]根据生态生理过程模型 BIOMEBGC模拟了不同的植被覆盖类型的最大光能利用率,具体取值如表1所示。
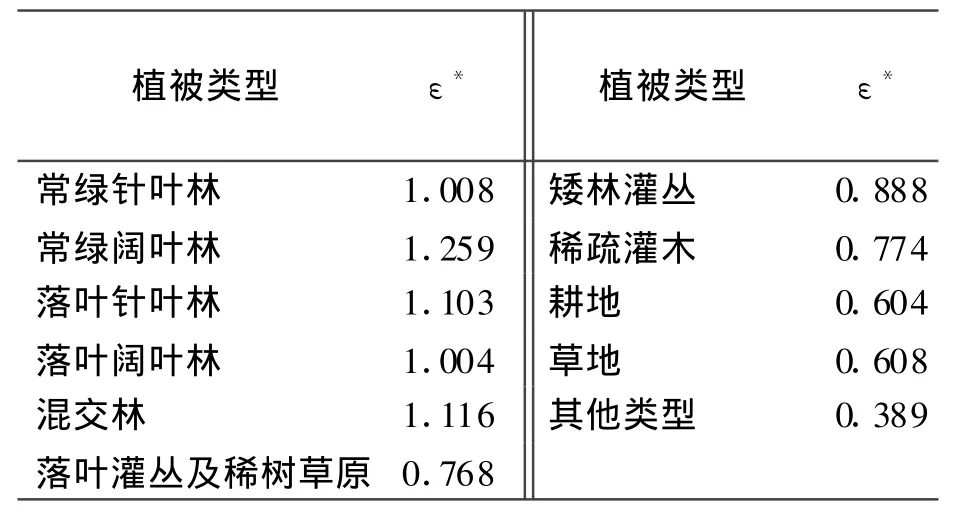
表1 不同植被类型的最大光能利用率 g/MJ
Lobell等[49]通过定量研究美国各县农作物的光能利用率,得到农田与天然植被具有不同的光能利用率的结论。Bradford等[50]指出目前在CASA模型中采用的光能利用率与地面实测数据相比较高,应该将其分为天然植被、C3农作物和C4农作物分别计算,草地具有低光能利用率,C4农作物具有高光能利用率。
CASA模型中的水分胁迫影响系数Wε的计算公式为:
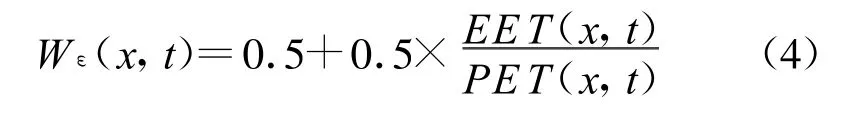
式中,PET为可能蒸散量(Potential Evapotranspiration),由Thornthwaite植被-气候关系模型求算,估计蒸散EET(Estimated Evapotranspiration)由土壤水分模型求算。需要的参数有月平均温度、月平均降水量、土壤中粘粒和砂粒所占的百分比以及土壤深度等变量,这些参数都需要大量野外实测工作的支持,并且在由点及面的尺度转换过程中会出现误差,数据精度无法保证。新近发展了一种遥感反演代表湿润指数(Land Surface Water Index,LSWI)的方法[52-54]。该方法在空间异质性方面表现较好,能够从时空尺度上较准确地反映地表湿润状况,计算公式如下:
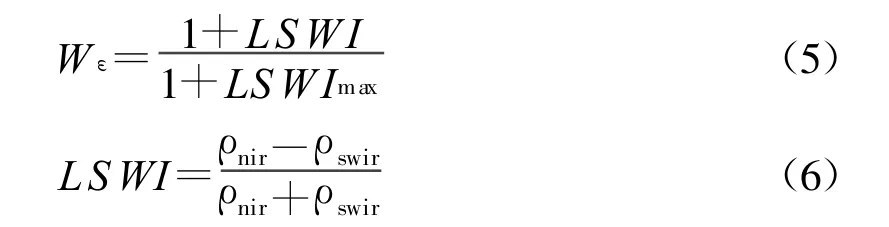
式中,ρnir和ρswir分别为近红外波段和短波红外波段,LSWImax为每种植被类型中全年最大的地表湿润度指数。
朴世龙[54]等利用CASA模型估算得到中国1997年植被NPP(以碳累积量计)为1.95 Gg,约为世界陆地植被年NPP的4.0%;其中海南岛南部、云南西南部、青藏高原东南部的热带雨林和季雨林地区植被NPP(以碳累积量计)最大,达到900 g/(m2·a)以上,而西部塔克拉玛干沙漠地区植被NPP不足10 g/(m2·a)。高清竹等[55]利用1981-2004年的遥感资料和气象数据,采用CASA模型估算藏北地区草地植被NPP,分析其变化趋势的空间格局及其对人类活动强度的响应。王艳艳等[56]利用改进的CASA模型对中国陆地生态系统1995年NPP进行了估算,并得到了国内NPP生产有机物质的总价值为 1.90×1012元/年。Yuan等[57]利用MODIS数据和其他地面基础观测数据,通过CASA模型模拟了河北省北部的植被NPP及其分布。张峰等[58]利用内蒙古草原连续13年的地上生物量资料在对CASA模型验证的基础上分析了该地区1982-2002年植被NPP的时间变异及其影响因子,模型得到的1982-2002年内蒙古草原平均年NPP(以碳累积量计)为 290.23 g/(m2·a),变化为 145.80~502.84 g/(m2·a),与降水呈极显著的相关关系,而与年均温无显著相关关系。张峰等[59]在中纬度半干旱区陆地样带之一的中国东北样带(Northeast China Transect,NECT)用CASA模型分析了1982-1999年植被NPP的时空变异及其影响因子,得到NECT植被NPP(以碳累积量计)为58~811 g/(m2·a),在过去 18年间整体呈显著增加趋势,其空间分布是由年降水量的分布格局决定,而时间变异则由年降水量和年太阳总辐射的变化驱动。
2.2 C-FIX模型 C-FIX模型也是基于Monteith理论的光能利用率模型,是近年来比较新的一个模型,可以在区域及全球尺度上估算3个基本的碳循环分量,即GPP、NPP和净生态系统生产力(Net Ecosystem Productivity,NEP)。它的主要设计者是比利时VITO(Flemish Institute for Technological Research)/TAP(Centre for Remote Sensing and Atmospheric Processes)研究所的Veroustraete Frank博士,其雏形是在1994年形成的[60-61]。C-FIX模型的第一版本主要考虑水分(包括降水和土壤水分)对光能利用率在较长时间序列(月/季节)上的制约作用和综合影响,第二版本将考虑把蒸散系数增加到模型之中。该模型在欧洲地区以及非洲大陆都已有了成功的应用[62-65]。经研究发现这种模型也适合于中国,如卢玲等[66]使用1998-2002年的 SPOT/VEGETATION遥感数据、全球格网化气象再分析资料以及黑河流域土地利用图估算了该流域不同生态系统净初级生产力(NPP)的年总量和平均生产力,输出了黑河流域NPP年累积量空间分布格局图、NPP季节动态图,分析了近5年来黑河流域NPP时空变化特征,评价了不同土地利用类型下的生态系统生产力水平差异。卢玲等[67]还应用C-FIX模型估算了中国西部地区的植被NPP。陈斌等[68]用C-FIX模型模拟了2003年中国陆地生态系统NPP,并对其空间格局进行了分析。
对于每一给定的点位置或格网,C-FIX模型使用以下公式来计算每天的NPP值(以碳累积量计),单位为g/(m2·d),下标d代表日值:
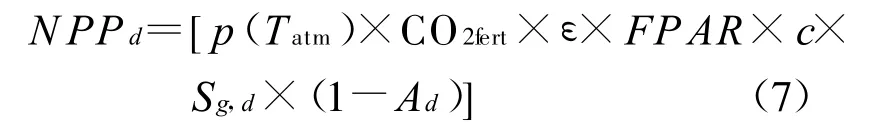
式中,p(Tatm)表示归一化气温依赖因子,CO2 fert表示归一化CO2施肥效应因子,它们的计算公式分别由Wang[69]和 Veroustraete[61]给出。ε表示植被在GPP阶段的光能利用率(以碳量计),其值约为1.1 g/MJ(APAR)[70]。FPAR表示植冠可吸收的光合有效辐射比,一般采用遥感植被指数推算。c表示光合有效辐射能占太阳入射总量的比例,在C-FIX模型中将之确定为常数0.48[71]。Sg,d表示太阳入射总辐射通量[MJ/(m2·d)]。Ad为植被自养呼吸率,模型采用Goward等[72]建立的依赖于温度的Ad简单算法。
C-FIX模型抓住了影响NPP的最主要的辐射、温度和水分因子,运用遥感数据计算FPAR,输出结果的时空分辨率很高。该模型的最大创新点在于它考虑了自工业革命以来全球大气CO2浓度显著增加对植被的光合作用和生产力产生的影响,即CO2施肥效应。大量试验表明C3植物对环境CO2浓度的变化最敏感[40,73],而全球C3植物占植物总数的95%以上;有实验证明在水分缺乏条件下植物对CO2浓度升高表现的正效应更大[73-74]。因此考虑CO2施肥效应对区域以及全球NPP的准确估算的重要性是显而易见的。但在该模型中对自养呼吸率的计算过于简化,只考虑了温度而忽略了其他的影响因素,不同的植被类型其自养呼吸率也有一定差异,但这个参数一般很难获得,这也是该模型今后将要改进的地方。
2.3 BIOM E-BGC模型 BIOME-BGC[31]模型是基于静态植被的生物地球化学模型,主要为了估算各类气候条件下生态系统的碳、氮和水在大气-群落-土壤间循环的特征以及温室气体(CO2、CH4和N2O)的交换通量,是从森林动力学模型发展而来的,以光合生物化学反应和土壤水平衡为基础,计算光合作用强度和NPP。在最新的模型版本中,许多生态系统行为/过程都是基于每日气象数据以天为时间步长进行模拟的。对于落叶的森林和草原植被类型,每年萌芽的物候期根据长期热量积累和土壤湿度来决定[75]。对于常绿植被类型来说,则需要根据当年的气象条件来决定生长期的开始和结束。在设计上它强调了下面2个基本原则:1)在全球范围内模型所需的输入数据易于取得或已有全球数据集;2)模型所需的生理生态参数已有公认的值或可以田间测量。在此基础上,该模型考虑了碳、水和能量在生态系统中输送通量的计算方法。
BIOME-BGC模型[76]采用光合酶促反应机理模型计算出GPP,减去生长呼吸和维持呼吸,得到碳的生物量积累,其中考虑了叶和细根的生物气候特征。通过植物体的凋落,特别考虑了大型凋落物的比例,植物积累的碳进入了枯枝落叶碳库。该模型模拟的水循环过程包括降雨、降雪、雪的升华、冠层蒸腾、土壤蒸发、蒸散、地表径流、土壤水分的变化及植被对水分的利用。该模型在土壤过程的模拟中考虑了凋落物分解进入土壤有机碳库的过程、土壤有机物矿化过程和基于木桶模型的水在土壤层间的输送关系。对能量过程主要考虑了净辐射、感热通量和潜热通量(基于Penman-Monteith方程)。另外,该模型还考虑了雪的融化和干扰。
BIOME-BGC模型的生理生态参数文件是按生态系统类型来进行参数化的,而不像其他模型那样按生物功能型或物种特性来参数化。分为以下几类:转换和死亡,分配比率,植物体各器官不稳定成分、纤维素、木质素组成比率,碳氮比,叶中核酮糖1,5-二磷酸加氧羧化酶中氮含量,比叶面积,叶片传导系数,树冠对水的截取和光的逃逸,水汽压亏缺和叶片水对传导的潜在控制。这些详细的参数可以使该模型在1 m2到全球范围内对主要生物群系的碳、水和氮每日的通量和状态进行模拟。
在国际上,BIOME-BGC模型已经被广泛地应用于碳循环估算及其对气候变化响应的研究中[33,77-82]。在国内,董文娟等[83]以小浪底-花园口区间洛河上游卢氏水文站以上流域作为研究区验证了BIOME-BGC模型在流域尺度的适用性,计算了研究区4种自然植被NPP的空间分布,与实测结果相比,常绿针叶林的日光合值接近模型的输出值,落叶阔叶林和灌木林的日光合值小于模型的输出值,草地的年NPP生产力大于模型的输出值。苏宏新[84]应用BIOME-BGC模型分析模拟了全球气候变化条件下新疆天山云杉林的生长。王超等[76]应用BIOME-BGC模型模拟了国际协调强化观测计划(CEOP)亚洲季风比较研究(CAMP)的半干旱地区吉林通榆地面观测基准站2002年10月-2003年9月草地和农田生态系统的潜热通量,并将模拟结果与通榆“干旱化和有序人类活动”长期观测站涡度相关法测定的观测值进行了比较,结果表明两者基本一致。曾惠卿等[85]通过BIOME-BGC模型估算了1993-2004年红壤丘陵区湿地松林GPP和NPP,并分析了GPP、NPP年际变化对气候的响应以及未来气候变化情景下的GPP、NPP响应。董明伟等[86]选取锡林河流域典型草原区沿水分梯度的4个代表群落,在野外实验测定和长期定位研究成果相结合的基础上,利用BIOME-BGC模型对代表群落的长期NPP动态进行了模拟和验证,对气候因子的敏感性分析及回归分析表明降水是该地区NPP最主要的决定因子,而温度决定作用相对较小。
MOD17产品是基于MODIS遥感参数,通过BIOME-BGC模型模拟的时间分辨率为8 d、空间分辨率为1 km的全球植被初级生产力产品数据集。目前已在全球不同区域对植被生长状况、生物量的估算、环境监测和全球变化等研究中得到广泛应用和验证[87-92]。但是针对全球设计的BIOME-BGC模型对区域初级生产力的估算误差较大,比如David等[93]用9个不同的生物类型和土地利用类型做了研究,得到MOD17产品在生产力水平比较低的区域因为输入的FPAR值较高而导致估测值偏高,在高生产力地区因为在GPP的算法中输入的光能利用率较低而导致估测值偏低的结论。Zhao等[94]在2005年探讨了MOD17数据集的缺陷,论证了该模型的输入数据MOD12Q1(全球土地覆盖分类数据集)、DAO(Data Assimilation Office)数据集和MOD15A2(FPAR和LAI数据集)的不确定性以及模型本身的设计缺陷,提出了修改方案。
2.4 其他遥感参数模型 其他应用遥感数据作为数据源的生态系统初级生产力模型还有BEPS[30]、Forest-BGC[32]、GLO-PEM[40]、RHESSYS[95]、SiB2[96]和TURC等[97]。
3 讨论与展望
由以上的论述可以看出遥感参数模型在研究大面积NPP方面所具有的优势和需要改进的地方,可归纳为以下几个方面。
3.1 遥感参数模型所需要的主要数据类型有:植被类型、气候数据和土壤质地数据。数据来源主要为卫星遥感和地面实测,对于点上的数据通常都是采用数据插值的方法推广到面上。现在国内存在的情况是数据严重不足,早期数据缺乏,地面观测站点分布不均匀而且比较分散,数据共享制度不完善。而数据精度对模型运行结果的影响明显。近年来随着一些新的传感器相继发射升空,如 SEA-WIFS 、POLDER、VEGETATION 、MODIS等,它们具有较NOAA/AVHRR更优化的光谱通道设计,可以提供更高分辨率和更高数据质量的土地覆盖、叶面积和FPAR等一系列与NPP模型输入有关的参数产品。同时,新技术的出现也为模型的设计和模拟结果的优化提出了新的思路,比如Chasmer等[98]利用空载激光雷达的数据去估测加拿大短松Pinusbanksiana的初级生产力,通过与实测数据和MOD17数据的对比,模拟结果良好。陈新芳等[99]利用微波可以穿透树冠的特点,提出可以将微波遥感引入到森林生物量的计算中去。Yang等[100]用支持向量机(Support Vector Machine,SVM)方法以美国为研究区设计了一个新的初级生物量模型。Daniel等[101]为了避免地面实测数据的误差和尺度转换的问题,设计了完全基于遥感数据EVI(Enhanced Vegetation Index)和LST(Land Surface Temperature)的TG(Temperature and Greenness)模型,在和涡度通量观测数据进行比较后发现,此模型的模拟结果优于MOD17产品。
因此如何更好地将新的遥感数据、计算机技术与模型相结合,改善模型的算法和模拟的精度将是未来的一个研究重点。
3.2 长期以来对NPP的研究大多集中在森林和农田,用现代技术对草地NPP的研究较少。利用遥感技术将草地与其他植被分开,准确研究草地NPP的变化及气候因子对草地畜牧业的发展有重要的意义。这是今后NPP研究的一个方向。
3.3 植被NPP过程模型大多是针对北美地区建立的,因为各个地方的差异,这些模型参数是否对中国有效,如何根据具体条件选取合适的模型和参数,是研究植被NPP的一个重要问题,需要严格的实验与论证。
3.4 遥感参数模型都是基于大尺度范围的,模型的精度验证是一个很困难的问题[102]。地面实验站点的数据很难推广到区域尺度,比如现在认为最准确的测定植被-大气间CO2湍流通量的方法是涡度相关技术,并在全球范围内成立了通量观测研究网络,但这也是基于点的数据集。目前模型验证主要通过模型间的互相比较,虽然可以说明一定问题,但是并不准确。因此,寻找合适的模型验证方法也是今后的一个重要研究方向。
[1] Ruimy A,Saugier B.Methodology for the estimation of terrestrial net primary production from remotely sensed data[J].Journal of Geophysical Research,1994,99:5263-5283.
[2] Field C B,Behrenfeld M J,Randerson J T,et al.Primary production of the biosphere:integrating terrestrial and oceanic components[J].Science,1998,281:237-240.
[3] Vitousek P M,Ehrlich P R,Ehrlich R H,et al.Human appropriation of the products of photosynthesis[J].Bio.Science,1986,36:368-373.
[4] 方精云,唐艳鸿,林俊达,等.全球生态学——气候变化与生态响应[M].高等教育出版社&施普林格出版社,2000.
[5] Lieth H.Promary production:terrestrial ecosystem[J].Human Ecology,1973(1):303-332.
[6] IGBP.The terrestrial carbon cycle:implications for Kyoto protocol[J].Science,1998,280:1393-1394.
[7] 李高飞,任海,李岩,等.植被净第一性生产力研究回顾与发展趋势[J].生态科学,2003,22(4):360-365.
[8] 胡自治,孙吉雄,张映生,等.高山线叶蒿草草地的第一性生产和光能转化率[J].生态学报,1988,8(2):183-190.
[9] 胡自治,孙吉雄,李洋,等.甘肃天祝主要高山草地的生物量及光能转化率[J].植物生态学报,1994,18(2):121-131.
[10] 杨福囤,王启基,史顺海.矮蒿草草甸生物量季节动态与年间动态[A].高寒草甸生态系统国际学术讨论会文集[C].北京:科学出版社,1988:61-70.
[11] 方精云,刘国华,徐嵩龄.我国森林植被的生物量和净生产量[J].生态学报,1996,16(5):497-508.
[12] 沈禹颖,阎顺国,朱兴运,等.河西走廊几种盐化草地第一性产生力的研究[J].草业学报,1995,4(2):44-50.
[13] Alexandrov G A,Oikawa T,Yamagata Y.The scheme for globalization of a process based model explaining gradations in terrestrial NPP and its application[J].Ecol Mod.,2002,148(3):293-306.
[14] Cramer W,Kicklighter D W,Bondeau A,et al.Comparing global models of terrestrial net primary productivity(NPP):Overview and key results[J].Global Change Biology,1999,(1):1-15.
[15] 邬建国.略谈理论和模型在生态学中的应用[J].生态学杂志,1994,13(3):76-79.
[16] Prince S D,Justice C D,Moore III B.Monitoring and Modeling of Terrestrial Net and Gross Primary Production[R].International Geosphere Biosphere Program(IGBP),Data and Information System(DIS),Global Analysis,Interpetation and Modeling(GAIM)Working Paper 1,1994:56.
[17] IPCC.Second assessment report of the Intergovernmental Panel on Climate Change(IPCC)[J].Environ Policy Law,1996,26(5):234-235.
[18] Ruimy A,Kergoat L,Bondeau A,et al.Comparing global models of terrestrial net primary productivity(NPP):analysis of differences in light absorption and light-use efficiency[J].Global Change Biology,1999,5(1):56-64.
[19] Lieth H.Modeling the primary production of the world[A].In:Lieth H,Whittaker R H.Primary Productivity ofthe Biosphere[C].New York:Springer-Verlag,1975:237-263.
[20] Box E.Quantitative evaluation of global primary productivity models generated by computers[A].In:Lieth H,Whittaker R H.Primary Productivity of the Biosphere[C].New York:Springer-Verlag,1975:305-328.
[21] Uchijima Z,Seino H.Agroclimatic evaluation of net primary productivity of natural vegetations(1)Chikugo model for evaluation net primary productivity[J].Journal of Agricultural Meteorology,1985,40:343-352.
[22] Bazilevich N I,Drozdov A V,Rodin L E.World forest productivity,its basic regularities and relationship with climatic factors[A].In:Duvigneard P III.Productivity of Forest Ecosystems[C].Paris:UNESCO,1971:345-353.
[23] Esser G.Osnabruck biosphere model:structure,construction,results[A].In:Esser G,Overdieck D.Modern Ecology:Basic and Applied Aspects[C].Amsterdam:Elsevier,1991:210-235.
[24] 刘明春,马兴祥,尹东,等.天祝草甸、草原草场植被生物量形成的气象条件及预测模型[J].草业科学,2001,18(3):65-68.
[25] 周光胜,张新时.自然植被净第一性生产力模型初探[J].植物生态学报,1995,19(3):193-200.
[26] 林慧龙,王军,徐震.草地净初级生产力与>0℃年积温、湿润度指标间的关系[J].草业科学,2005,22(6):8-10.
[27] Esser G.Modelling global terrestrial sources and sinks of CO2with special reference to soil organic matter[A].In Bouwman A F.Soils and the Greenhouse Effect[C].New York:John Wiley&Sons,1990:247-262
[28] Goetz S J,Prince S D,Goward S N,et al.Satellite remote sensing of primary production:an improved production efficiency modeling approach[J].Ecological Modelling,1999,122:239-255.
[29] Melillo J M,McGuire A D,Kicklighter D W,et al.Global climate change and terrestrial net primary production[J].Nature,1993,363:234-240.
[30] Liu J,Chen JM,Cihlar J,et al.A process-basedboreal ecosystem productivity simulator using remote sensing inputs[J].Remote Sensing of Environment,1997,62:158-175.
[31] Melillo J M,McGuire A D,Kicklighter D W,et al.Global climate change and terrestrial net primary production[J].Nature,1993,363:234-240.
[32] Running S W,Nemani R R,Peterson D L,et al.Mapping regional forest Evapotraspiration and photosynthesis by coupling satellite data with ecosystem simulation[J].Ecology,1989,70:1090-1101.
[33] Hunt E R,Piper S C,Nemani R,et al.Global net carbon exchange and intra-annual atmospheric CO2concentrations predicted by an ecosystem simulation modeland three-dimensionalatmospheric transport model[J].Global Bipgeochemical Cycles,1996(10):431-456.
[34] 周才平,欧阳华,王勤学,等.青藏高原主要生态系统净初级生产力的估算[J].地理学报,2004,59(1):74-79.
[35] 冯险峰.基于过程的中国陆地生态系统生产力和蒸散遥感研究[D].北京:中国科学院地理科学与资源研究所,2004.
[36] Chen J M,Liu J,Cihlar J,et al.Daily canopy photosynthesis model through temporal and spatial scaling for remote sensing applications[J].Ecological M odeling,1999,124:99-119.
[37] Monteith J L.Solar radiation and productivity in tropical ecosystems[J].Journal of Applied Ecology,1972(9):747-766.
[38] Monteith J L.Climate and efficiency of crop production in Britain[J].Philosophical Transactions of the Royal Society of London Series B,Biological Science,1977:277-294.
[39] Field C B,Randerson J T,Malmstrom C M.Global net primary production:Combining ecology and remote sensing[J].Remote Sensing Environment,1995,51:74-88.
[40] Prince S D,Goward S N.Global primary production:A remote sensing approach[J].Journal of Biogeography,1995,22:815-835.
[41] Potter C S,Randerson J,Field C B,etal.Terrestrial ecosystem production:a process model based on global satellite and surface data[J].Global Biogeochemical Cycle,1993(7):811-841.
[42] Veroustraete F,Sabbe H,Eerman E.Estimation of carbon mass fluxes over Europe using the C-FIX model and Euroflux data[J].Remote Sensing of Environment,2002,83:376-399.
[43] 朱文泉,陈云浩,徐丹,等.陆地植被净初级生产力计算模型研究进展[J].生态学杂志,2005,24(3):296-300.
[44] Field C S,Randerson J T,Malmstrom C M.Global net primary production:combining ecology and remote sensing[J].Remote Sensing of Environment,1995,51:74-88.
[45] Potter C S,Klooster S A.Global model estimate of carbon and nitrogen storage in litter and soil pools:response to changes in vegetation quality and biomass allocation[J].Tellus,1997,49:1-17.
[46] 彭少麟,郭志华,王伯荪.利用GIS和 RS估算广东植被光利用率[J].生态学报,2000,20(6):903-909.
[47] 张娜,于贵瑞,于振良,等.基于3S的自然植被光能利用率的时空分布特征的模拟[J].植物生态学报,2003,27(3):325-336.
[48] Runing S W,Thornton P E,Nemani R.Global terrestrial gross and net primary productivity from the earth observing system[A].In:Sala O E,Jackson R B,M ooney H A,et al.Methods in ecosystem science[C].New York:Springer Verlag,2000:44-57.
[49] Lobell D B,Hicke J A,Asner G P,et al.Satellite estimates of productivity and light use efficiency in United States agriculture,1982-1998[J].Global Change Biology,2002(8):722-735.
[50] Bradford J B,Hicke J A,Lauenroth W K.The relative importance of light-use efficiency modifications from environmental conditions and cultivations for estimation of large-scale net primary productivity[J].Remote Sensing of Environment,2005,96:246-255.
[51] Ceccato P,Flasse S,Tarantola S,etal.Detecting vegetation leaf water content using reflectance in the optical domain[J].Remote Sensing of Environment,2001,77:22-33.
[52] Boles S,Xiao X,Liu J,et al.Land cover characterization of Temperate East Asia:Using multi-temporal image data of VEGETATION sensor data[J].Remote Sensing of Environment,2004,90:477-489.
[53] Xiao X M,Zhang Q Y,Saleskab S,et al.Satellitebased modeling of gross primary production in a seasonally moist tropical evergreen forest[J].Remote Sensing of Environment,2005,94:105-122.
[54] 朴世龙,方精云,郭庆华.利用CASA模型估算我国植被净第一性生产力[J].植物生态学报,2001,25(5):603-608.
[55] 高清竹,万运帆,李玉娥,等.藏北高寒草地NPP变化趋势及其对人类活动的响应[J].生态学报,2007,27(11):4612-4619.
[56] 王艳艳,杨明川,潘耀忠,等.中国陆地植被生态系统生产有机物质价值遥感估算[J].生态环境,2005,14(4):455-459.
[57] Yuan J G,Niu Z,Wang C L.Vegetation NPP Distribution Based on MODIS Data and CASA Model-A Case Study of Northern Hebei Province[J].Chinese Geographical Science,2006,16(4):334-341.
[58] 张峰,周广胜,王玉辉.基于CASA模型的内蒙古典型草原植被净初级生产力动态模拟[J].植物生态学报,2008,32(4):786-797.
[59] 张峰,周广胜.中国东北样带植被净初级生产力时空动态遥感模拟[J].植物生态学报,2008,32(4):798-809.
[60] Veroustraete F,Patyn J,Myneni R B.Forcing of a simple ecosystem model with FAPAR and climatic data to estimate regional scale photosynthetic assimilation[A].Vegetation,Modelling and Climate Change Effects[C].The Hague,Netherlands:Academic Publishing,1994:151-177.
[61] Veroustraete F.On the use of ecosystem modeling for the interpretation of climate change effects at the ecosystem level[J].Ecological Modelling,1994,75-75:221-237.
[62] Veroustraete F,Patyn J,Myneni R B.Estimating net ecosystem exchange of carbon using the Normalized Difference Vegetation Index and an Ecosystem Model[J].Remote Sensing of Environment,1996,58:115-130.
[63] Sabbe H,Veroustraete F.Estimation of the carbon balance ofEuropean terrestrialecosystems by means of the C-Fix model[C].Copenhagen:Proceedings of The 1999 EUMETSAT Meteorological Satellite Data Users’Conference,1999:271-278.
[64] Sabbe H,Veroustraete F.Demonstration of a standard Net Primary Productivity product for the SPOT4-VEGETATION instrument[R].Vito report:9 ex.DWTC:3 ex.TAP/062,2000.
[65] Veroustraete F,Sabbe H,Herman E.Estimation of carbon mass fluxes over Europe using the C-FIX model and Euroflux data[J].Remote Sensing of Environment,2002,83:376-399.
[66] 卢玲,李新,Frank V.黑河流域植被净初级生产力的遥感估算[J].中国沙漠,2005,25(6):823-830.
[67] 卢玲,李新,Frank V.中国西部地区植被净初级生产力的时空格局[J].生态学报,2005,25(5):1026-1032.
[68] 陈斌,王绍强,刘荣高,等.中国陆地生态系统NPP模拟及空间格局分析[J].资源科学,2007,29(6):45-53.
[69] Wang F K Y.Canopy CO2exchange of Scots pine and its seasonal variation after four year exposure to elevated CO2and temperature[J].Agricultural and Forest Meteorology,1996,82:1-27.
[70] Wofsy F S C,Goulden F M L,Fan F S M,et al.Net exchange of CO2in midlatitude forests[J].Science,1993,260:1314-1317.
[71] McCree K J.Test of current definitions of photosynthetically active radiation against leaf photosynthesis data[J].Agricultural Meteorology,1972(10):442-453.
[72] Goward S N,Dye D G.Evaluating North-American net primary productivity with satellite observation[J].Advanced Space Research,1987,7(11):165-174.
[73] 蒋高明,韩兴国,林光辉.大气CO2浓度升高对植物的直接影响——国外十余年来模拟实验研究之主要手段及基本结论[J].植物生态学报,1997,21:489-502.
[74] 蒋高明,林光辉,Marino B D V.美国生物圈二号内生长在很高CO2浓度下的10种植物气孔导度、蒸腾速率及水分利用效率的变化[J].植物学报,1997,39:546-553.
[75] White M A,Thornton P E,Running S W.A continental phenology model for monitoring vegetation responses to inter-annual climatic variability[J].Global Biogeochemical Cycles,1997,11(2):217-234.
[76] 王超,延晓冬,黄耀,刘辉志.应用BIOME-BGC模型研究典型生态系统的潜热通量——半干旱地区吉林通榆的模拟[J].气候与环境研究,2006,11(3):404-412.
[77] Korol R L,Running S W,Milner K,et al.Testing a mechanistic carbon balance model against observed tree growth[J].Canadian Journal of Forest Resources,1991,21:1098-1105.
[78] Pan Y,Melillo J M,McGuire A D,et al.Modeled responses of terrestrial ecosystems to elevated atmospheric CO2:a comparison of simulations by the biogeochemistry models of the Vegetation/Ecosystem Modeling and Analysis Project(VEMAP)[J].Oecologia,1998,114:389-404.
[79] Keyser A R,Kimball J S,Nemani R R,etal.Simulating the effects of climate change on the carbon balance of North American high-latitude forests[J].Global Change Biology,2000,6(S1):185-195.
[80] Thornton P E,Law B E,Gholz H L,et al.Modeling and measuring the effects of disturbance history and climate on carbon and water budgets in evergreen needle leaf forests[J].Agricultural and Forest Meteorology,2002,113(1-4):185-222.
[81] Churkina G,Tenhunen J,Thornton P,et al.Analyzing the ecosystem carbon dynamics of four european coniferous forests using a biogeochemistry model[J].Ecosystems,2003(6):168-184.
[82] Law B E,Turner D,Campbell J,et al.Disturbance and climate effects on carbon stocks and fluxes across Western Oregon USA[J].Global Change Biology,2004,10(9):1429-1444.
[83] 董文娟,齐晔,李惠民,等.植被生产力的空间分布研究——以黄河小花间卢氏以上流域为例[J].地理与地理信息科学,2005,21(3):105-108.
[84] 苏宏新.全球气候变化条件下新疆天山云杉林生长的分析与模拟[D].北京:中国科学院研究生院,2005.
[85] 曾慧卿,刘琪璟,冯宗炜,等.基于BIOM E-BGC模型的红壤丘陵区湿地松(Pinus elliottii)人工林GPP和NPP[J].生态学报,2008,28(11):5314-5321.
[86] 董明伟,喻梅.沿水分梯度草原群落NPP动态及对气候变化响应的模拟分析[J].植物生态学报,2008,32(3):531-543.
[87] Zhao M,Heinseh F A,Nemani R R,etal.Improvements of the MODIS terrestrial gross and net primary production global data set[J].Remote Sensing of Environment,2005,95(2):164-176.
[88] Rasmus F,Inge S,Michael S R,et al.Evaluation of satellite based primary production modeling in the semi-arid Sahel[J].Remote Sensing of Environment,2006,105:173-188.
[89] 郭晓寅,何勇,沈永平,等.基于 MODIS资料的2000-2004年江河源区陆地植被净初级生产力分析[J].冰川冻土,2006,28(4):512-518.
[90] 何勇,董文杰,季劲均,等.基于 AVIM 的中国陆地生态系统净初级生产力模拟[J].地球科学进展,2005,20(3):345-349.
[91] 张晓浩,黎夏,施迅,等.广东省水稻生物质能的估算[J].遥感信息,2007(1):26-29.
[92] 国志兴,王宗明,张柏,等.2000-2006年东北地区植被NPP的时空特征及影响因素分析[J].资源科学,2008,30(8):1226-1235.
[93] David P T,William D R,Warren B C,et al.Evaluation of MODIS NPP and GPP products across multiple biomes[J].Remote Sensing of Environment,2006,102:282-292.
[94] Zhao M S,Faith A H,Ramakrishna R N,et al.Improvements of the MODIS terrestrial gross and net primary production global data set[J].Remote Sensing of Environment,2005,95:164-176.
[95] Band L E,Peterson D L,Running S W,et al.Forest ecosystem process at the watershed scale:Basis for distributed simulation[J].Ecological Modelling,1991,56:171-196.
[96] Sellers P J,Los S O,Tucher C J,et al.A revised land surface parameterization(SiB2)for atmospheric GCMS.Part II:The generationof global fields of terrestrial biophysical parameters from satellite data[J].Journal of Climate,1996(9):706-737.
[97] Ruimy A,Dedieu G,Saugier B.TURCL a diagnostic model of continental gross primary productivity and net primary productivity[J].Global Biogeochemical Cycle,1996(10):269-285.
[98] Chasmer L,Hopkinson C,McCaughey H,et al.Scaling and assessment of GPP from MODIS using a combination of airborne lidar and eddy covariance measurements over jack pine forest[J].Remote Sensing of Environment,2009,113:82-93.
[99] 陈新芳,安树青,陈镜明,等.森林生态系统生物物理参数遥感反演研究进展[J].生态学杂志,2005,24(9):1074-1079.
[100] Yang F H,Kazuhito I,Michael A W.Developing a continental-scale measure of gross primary production bycombining MODIS and AmeriFlux data through Support Vector Machine approach[J].Remote Sensing of Environment,2007,110:109-122.
[101] Daniel A S,Abdullah F R,Vicente D C,et al.A new model of gross primary productivity for North American ecosystems based solely on the enhanced vegetation index and land surface temperature from MODIS[J].Remote Sensing of Environment,2008,112:1633-1646.
[102] 崔霞,冯琦胜,梁天刚.基于遥感技术的植被净初级生产力研究进展[J].草业科学,2007,24(10):36-42.
猜你喜欢
童话王国·奇妙逻辑推理(2024年5期)2024-06-19 16:03:38
河北地质(2022年2期)2022-08-22 06:24:04
中学生数理化·七年级数学人教版(2020年10期)2020-11-26 08:24:50
数学物理学报(2020年2期)2020-06-02 11:29:24
现代园艺(2017年23期)2018-01-18 06:58:12
红土地(2016年3期)2017-01-15 13:45:17
光学精密工程(2016年6期)2016-11-07 09:07:19
应用海洋学学报(2015年2期)2015-11-22 07:36:28
海峡姐妹(2015年9期)2015-02-27 15:13:07
杭州科技(2014年3期)2014-02-27 15:26:46
