
高地省藤幼苗光合作用日变化特征
2010-05-08 15:25官凤英范少辉刘亚迪邓旺华
浙江林业科技 2010年5期
关键词:午休
官凤英,范少辉,刘亚迪,邓旺华,彭 颖
(国际竹藤网络中心 竹藤科学与技术重点实验室 生物资源利用科学研究院,北京 100102)
高地省藤幼苗光合作用日变化特征
官凤英,范少辉*,刘亚迪,邓旺华,彭 颖
(国际竹藤网络中心 竹藤科学与技术重点实验室 生物资源利用科学研究院,北京 100102)
摘要:利用Li-6400便携式光合测定仪研究了高地省藤苗期光合作用日变化特征,结果表明:①高地省藤5个月幼苗Pn、Tr日变化曲线均呈双峰型,12:00左右出现明显的光合“午休”现象,峰值出现在10:00和14:00左右,“午休”过后,气孔扩张,且蒸腾增加量高于Pn增加量;②水分利用效率WUE和气孔限制值Ls也呈双峰趋势,在10:00均达最大值,WUE光合“午休”时降至最低,16:00出现次高峰,Ls变化较WUE平缓,光合“午休”时略有下降,18:00时降到最低值;③相关性分析结果表明:对Pn影响最大的因子是Ls,其次是Gs、WUE、Tr、RH、PAR、VPD、Ca、Ta,这些因子与Pn均呈正相关,但均未达到显著水平,高地省藤光合速率变化是多因素综合作用的结果。
关键词:高地省藤;净光合速率;日变化;光合“午休”
棕榈藤(rattan)是热带和亚热带森林中集材用、药用、食用于一体的多用途植物,具有较大的开发利用价值[1~2],一些学者在藤苗光照与生长方面开展了部分研究[3~5]。Aminuddin认为棕榈藤的光强适应范围较广;尹光天,许煌灿等通过不同光照条件下藤苗生长调查研究分析了白藤、黄藤、单叶省藤等藤种最适的光照条件,认为在全光照下藤苗生长不良,不同藤种在苗期需要适当遮荫。Dransfield研究得出多数藤种幼苗时需要遮荫,抽茎快速生长时又需要较强的光照,乃至全光照。这些研究都只是从不同栽培措施的角度分析了光照与苗木生长的关系,对于苗木本身的光合作用机理及其对环境条件适应性的相关研究尚未见报道。
高地省藤(Calamus nambariensisvar.alpinus)是棕榈科(Palmae)省藤亚科(Calamoideae)省藤族(Calameae)省藤属(Calamus)植物,产于我国云南省南部,生于海拔1 400~1 900 m的常绿阔叶林中[6~7],因其藤茎(藤条)质地柔韧,外形美观素雅,抗弯抗拉性强,常用于编织各种藤器、家具[8~9]。光合作用是研究植物生长生理过程的基本内容,是构成植物生产力的主要因素,本文通过对高地省藤叶片光合生理特性的研究,探索其光合作用的规律,以期为苗木培育提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为栽培于北京中国林科院温室的高地省藤盆栽实生苗,苗龄5个月,培养基质为腐殖土:珍珠岩:蛭石(体积比)= 3:1:1,培养期间相对光照强度为65%,正常水肥管理。
1.2 测定方法
2007年9月初,选择晴天,用Li-6400便携式光合测定仪(Li-Cor,Inc., Lincoln, USA)测定光合速率和环境因子日变化。选生长条件一致的苗木3株,每株测定上部3~5片完整成熟的叶片。测定时间8:00-18:00每2 h测1次,测定参数包括叶片净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)、大气CO2浓度(Ca)、饱和蒸汽压差(VPD)、大气温度(Ta)、大气相对湿度(RH)、光合有效辐射(PAR)等。气孔限制值Ls和叶片瞬时水分利用效率WUE(μmolCO2·mmol-1)利用以下公式计算:

1.3 数据处理
试验数据采用Excel进行统计处理,均采用各测量指标的平均值,采用SPSS 13.0软件系统进行相关性分析和方差分析。
2 结果与分析
2.1 环境因子的日变化2.1.1 光合有效辐射和大气CO2浓度日变化 植物光合作用受各种环境因素影响,尤其是光照强度,当植物吸收的光能超过其利用能力时会引起叶片光合作用的光抑制[12]。如图1所示,在8:00-18:00,光合有效辐射(PAR)的日变化呈先升高后降低的“单峰”曲线,变化范围为118.917~1178.556 μmol·m-2·s-1,在12:00时达到最高峰值,之后逐渐下降,18:00降到最低值。大气CO2变化幅度不大,浓度在8:00时最高,然后逐渐下降,12:00时最低,14:00以后又有缓慢上升的趋势。

图1 高地省藤苗PAR和Ca的日变化曲线Figure 1 Diurnal variations ofPARandCainC. nambariensisvar.alpinusseedlings

图2 高地省藤苗RH和Ta的日变化曲线Figure 1 Diurnal variations ofRHandTainC. nambariensisvar.alpinusseedlings
2.1.2 大气温度和相对湿度日变化 大气温度(图2)呈单峰曲线变化,变化范围为24.9~36.6℃,在12:00到14:00温度高达36℃以上。大气温度与相对湿度呈明显的负相关(r =-0.909*)。大气相对湿度(RH)呈一个低缓的先下降后上升的变化趋势,湿度变化范围为28.9%~53.8%,在8:00最大,14:00时左右最小,16:00以后随着温度的下降又逐渐上升。
2.2 光合参数日变化
2.2.1 光合速率日进程5个月生盆栽高地省藤苗叶片的净光合速率日变化呈“双峰”曲线(图3),在10:00时净光合速率达到第一个高峰,峰值为2.713 μmol·m-2·s-1,在12:00出现光合“午休”现象,Pn为0.829 μmol·m-2·s-1,14:00时达到第二个高峰,峰值为1.714 μmol·m-2·s-1,第二个峰值约为第一个峰值的63%,达到第2个峰值后,Pn值基本上保持平稳近2 h,到18:00时才开始有下降的趋势。Pn处于第二个峰时强光照和高温条件促进了叶片的光呼吸强度,增加了呼吸消耗,因而导致净光合速率比第一个峰低[13]。从图1、图2还可以看出,上午的光合速率明显高于下午的光合速率,是由于上午环境中的CO2浓度较高,而下午由于植物的呼吸作用使得环境中CO2浓度偏低所致。
2.2.2 蒸腾速率的日变化 植物通过蒸腾作用调节叶面温度、供应光合作用的水分,与光合速率关系密切。图3显示Tr的日变化与光合速率相似也呈规则的“双峰”曲线,两个主峰分别出现在10:00和14:00,14:00的峰值(1.164 mmol·m-2·s-1)高于10:00(0.754 mmol·m-2·s-1)。在12:00达到最低谷(0.587 mmol·m-2·s-1),Tr在达到第2个高峰后则开始下降。Tr的日变化趋势并不是随着Pn变化而变化,在14:00温度升高、湿度下降的环境下Tr值升高的幅度大于Pn,一些研究显示这可能与植物叶片角质层蒸腾较大有关[14]。
2.2.3 气孔导度、饱和蒸气压差日变化 图4显示高地省藤苗气孔导度日变化趋势与Pn一致,最大值出现在10:00,随着PAR和Ta的增大,Gs迅速降低,12:00时Gs值最低(0.014 cm·s-1)。14:00以后又逐渐增高。饱和蒸汽压差呈“单峰”曲线,在12:00-14:00出现最高值(4.246 kPa),与光合“午休”时间一致,12:00以后,Gs仍保持相对较高的水平,但是高地省藤的Pn已经开始下降(图3),VPD也开始下降,可见高地省藤在发生光合“午休”一直到下午光合作用的整个变化趋势是Gs和VPD综合作用的结果,其中叶片水分失衡,水蒸气压亏缺增大,Gs降低两种生态因子共同作用导致植物光合“午休”[15~16]。14:00时气孔导度起决定作用,随着时间的推移,VPD成为关键因子。

图3 高地省藤苗Pn和Tr的日变化曲线Figure 3 Diurnal variations ofPnandTrinC. nambariensisvar.alpinusseedlings

图4Gs和VPD的日变化Figure 4 Diurnal variations ofGsandVPDinC. nambariensisvar.alpinusseedlings
2.2.4 水分利用效率、气孔限制值日变化 水分利用效率(WUE)是指消耗单位重量的水分植物所固定的CO2的量,这由植物的Pn和Tr两方面所决定[17]。图5显示了高地省藤对光合作用的两种底物H2O和CO2的利用情况,在10:00之前,高地省藤的WUE值处于很高的水平,之后发生光合“午休”时,WUE降至最低值(1.412 μ molCO2·mmol-1),此时耗水量大,随后虽有上升的趋势,但是仍处于较低水平,16:00时出现次高峰。
气孔限制值(Ls)可以反映植物叶片对大气CO2的相对利用效率[18],由图5可见,Ls的日变化出现“双峰”趋势,变化较WUE平缓,Ls在Pn达到最高峰的10:00时也达到全天的最高值(0.393 %),可见此时植物充分消
耗利用光合底物进行光合作用,之后在光合“午休”时Ls虽然也有降低的趋势,但并不明显,直到18:00植物的Pn最低时Ls也降到最低。
2.2.5 高地省藤光合特征参数的相关性分析 由表1可知,从相关系数的大小来看,环境因子中对高地省藤Pn影响作用由大到小顺序为:Ls>Gs>WUE>Tr>RH>PAR,且这些因子与Pn均呈正相关,但均未达到显著水平。环境因子对Tr的影响同样未达到显著水平,从相关系数的大小来看,RH与Tr呈较大的负相关,Ta和VPD与其呈较大的正相关,相关系数均达到0.600以上水平。Ca与Ta呈显著负相关(r =-0.869*),RH与VPD呈极显著负相关(r =-0.931**),Ta与VPD呈极显著正相关(r = 0.983**);RH与Ta呈显著负相关(r =-0.909*),与WUE呈显著正相关(r = 0.907*)。

图5 水分利用效率与气孔限制值的日变化曲线Figure 5 Diurnal variations ofWUEandLsin C. nambariensis var. alpinus seedlings
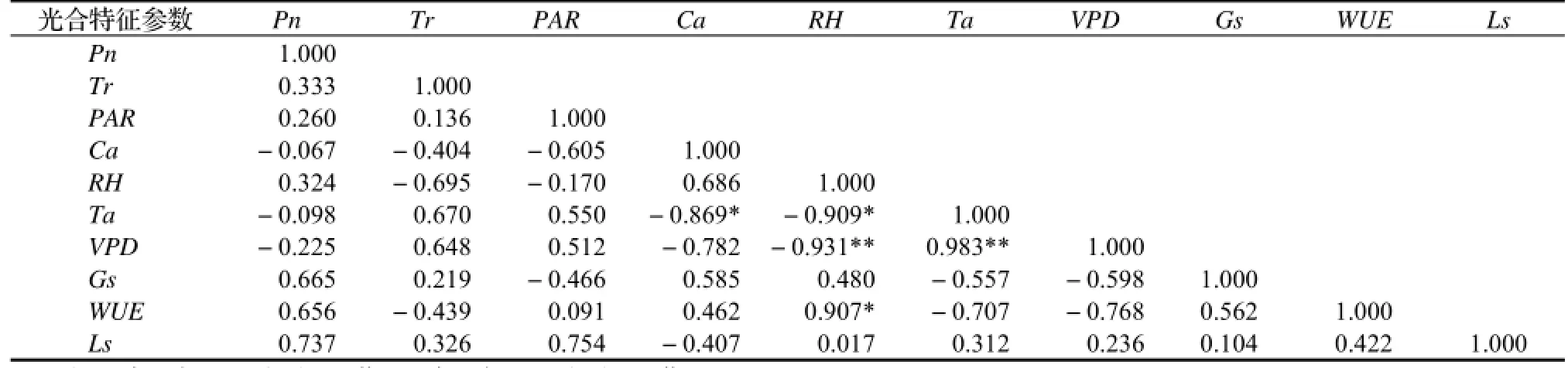
表1 高地省藤各光合特征参数与环境因子的相关性分析Table 1 Correlation analysis between photosynthetic characteristic parameters and environmental factors forC. nambariensisvar.alpinusseedlings
3讨论
高地省藤Pn日变化呈双峰曲线,“午休”现象比较明显,发生光合“午休”时,随着PAR、Ta达到全天的最高值,VPD增大,进而降低叶片水势,引起气孔关闭,发生光合作用的底物供应不足,Pn降至最低,水分供应不足,Tr也瞬时降低。有研究认为几乎所有的中生和旱生植物都是可以通过气孔部分关闭即降低气孔导度来适应午间叶片过度失水的环境,这样不仅可以直接减少水分蒸腾,还能有效地减少到达光合组织表面被吸收的辐射总量,从而减轻光化学损伤以及热负荷带来的负面影响[19]。而对于高地省藤来说,光合“午休”表现出它在长期的进化过程中对高温环境的一种适应,这也与他自身羽状全裂的叶片形态结构以及热带亚热带的地理分布特征相吻合。总的说来,植物发生光合“午休”现象,一般认为是由气孔因素和非气孔因素两方面原因造成的。Farquhar和Sharkey(1982)研究得出只有当Pn与Ci的变化方向相同,即两者同时减小(本文Ci无明显变化),且Ls增大时(本文Ls降低),才可认为Pn的下降主要是由Gs引起的,否则Pn的下降要归因于植物叶肉细胞羧化能力的降低[20]。因此高地省藤是否由气孔因素导致光合“午休”还有待进一步研究。
“午休”过后,在14:00时,PAR有所下降,此时Gs随即上升到一个高峰值,并维持一个最大程度气孔扩张来提高蒸腾(图2)以降低叶温,由于一部分水分用于蒸腾,发生光合作用的瞬时水分利用效率虽然比“午休”时有所缓和,但仍不高(图4)。从理论上讲,CO2的扩散阻力是水蒸气的0.64倍[21],因此气孔导度对Pn的影响比Tr大,然而14:00时,Tr的上升幅度比Pn的大,这一方面是光呼吸增大的原因,另一方面可能与叶片角质层结构蒸腾较大有关。
省藤属的植物一般是C3植物[22],而一般来说,C3植物的水分利用效率比景天酸植物和C4植物的都低[23~24]。本文的研究结果显示其WUE值在光合“午休”过后一直较低,这也与前人的研究结果有一定的相似性。此外用便携式光合测定仪测定并计算得到的水分利用效率只能代表某特定时间内植物部分叶片的行为[25],因此WUE仅反映高地省藤对测定环境的一种适应能力,至于高地省藤长期的水分利用情况仍需进一步研究。
参考文献:
[1] 江泽慧,萧江华,许煌灿. 世界竹藤[M]. 沈阳:沈阳科技出版社,2002. 509-612.
[2] 尹光天,许煌灿,张伟良,等. 棕榈藤物种的收集和引种驯化的研究[J]. 林业科学研究,1993,6(6):609-617.
[3] Aminuddin M. Performances of some rattan trial plots in Peninsular Malaysia[M] //Wong K M, Manokaran N. (Eds.). Proceedings of the Rattan Seminar, Kuala Lumpur, Malaysia: RIC, 1985: 49-56.
[4] 许煌灿,尹光天,曾炳山. 棕榈藤的研究[M]. 广东科技出版社,1994. 94-99.
[5] Dransfield J. The rattan taxonomy and ecology. In:Rao A.N. and V. Ramanatha Rao(eds.)Rattan-Taxonomy, ecology, silviculture, conservation, genetic improvement and biotechnology. Proceedings of training courses curn workshop,14-26 April 1996, Sarawak, Sabah. IPGRI-APO Serdang, Malaysia.1997,1-14.
[6] 江泽慧,范少辉,张昌顺,等. 棕榈藤研究进展[J]. 江西农业大学学报,2007,29(6):957-964, 1 005.
[7] 中国科学院中国植物志编辑委员会. 中国植物志(第13卷)[M]. 北京:科学出版社,1991. 60-100.
[8] 江泽慧,吕文华,费本华,等. 3种华南商用藤材的解剖特性[J]. 林业科学,2007,43(1):121-126.
[9] 李旸,腰希申.棕榈藤的电镜观察[J]. 林业科学,2002,38(1):173-174.
[10]Berry J A, Downton W J S. Environmental Regulation of Photosynthesis[M]. New York: Academic Press, 1982. 263-343.
[11]FischerR A,Turner N C. Plant Productivity in the Arid and Semiarid Zones[J]. Ann Rev Plant Physiol,1978(29):227-317.
[12]PearcyR W. Sunflecks and photosynthesis in plant canopies[J]. Ann Rev Plant Physiol,1990(41):421-453.
[13] 万素梅,贾志宽,杨宝平. 苜蓿光合速率日变化及其与环境因子的关系[J]. 草地学报,2009,17(1):27-31
[14] Larcherw. 李博(译). 植物生态生理学[M]. 北京:科学出版社,1980. 192-194.
[15] 高辉远,邹琦,程炳嵩. 大豆光合日变化与内生节律的关系[J]. 植物生理学通讯,1992,28(4):262-264.
[16] 郑国生,邹琦. 不同天气条件下田间大豆光合作用日变化的研究[J]. 中国农业科学,1993,26(1):44-50.
[17] 孔蓓蓓,刘超,尹伟伦,等.沙柳、黄柳和杞柳光合作用的日变化[J]. 河南科技大学学报(自然科学版),2009,30(3):79-83.
[18] 徐炳成,山仑,黄占斌. 黄土丘陵区柳枝稷与白羊草光合生理生态特征的比较[J].中国草地,2003,25(1):1-4.
[19] 鞠强,贡璐,杨金龙,等. 梭梭光合生理生态过程与干旱环境的相互关系[J]. 干旱区资源与环境,2005,19(4):201-204.
[20] Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis[J]. Ann Rev Plant Physiol, 1982,(33):317-345.
[21] 赵平,曾小平,彭少麟,等. 海南红豆夏季叶片气孔交换、气孔导度和水分利用效率的日变化[J]. 热带亚热带植物学报,2000,8(1):35-42.
[22] 李荣生. 华南地区3个棕榈藤种水分利用效率和抗旱能力的研究[D]. 北京:中国林业科学研究院,2003.
[23] Nobel P S. Ac hievable productivities of certain CAM plants: Basis for high values compared with C3 and C4 plants[J]. New Phy tol,1991(119):183-205.
[24] 蒋高明,何维明. 毛乌素沙地若干植物光合作用、蒸腾作用和水分利用效率种间及生境间差异[J]. 植物学报,1999,41(10):1 114-1 124.
[25] Martin B J, Thorstenson Y R. Stable carbon isotope composition (δ 13C) water-use efficiency, and biomass productivity ofLycoperscon esculentum,Lycoperscon pennelliiand the F1 hybrid[J]. Plant Physiol, 1988(88):213-217.
[26] 王 瑞,陈永忠,王湘南,等. 油茶优良无性系光合作用的日变化[J]. 中国农学通报,2009,25(24):236-239.
[27] 冯建灿,张玉洁. 喜树光合速率日变化及其影响因子的研究[J]. 林业科学,2002,38(4):34-39.
[28] 何文兴,易津,李洪梅. 根茎禾草乳熟期净光合速率日变化的比较研究[J]. 应用生态学报,2004,15(2):205-209.
[29] 王会肖,刘昌明. 作物光合、蒸腾与水分高效利用的试验研究[J]. 应用生态学报,2003,14(10):1 632-1 636.
中图分类号:S718.45
文献标识码:A
文章编号:1001-3776(2010)05-0033-05
收稿日期:2010-06-30;修回日期:2010-08-20
基金项目:林业科学技术推广项目“棕榈藤种质资源培育及利用技术推广与示范”(〔2005〕86);国际竹藤网络中心科研专项“优质棕榈藤苗期生长及光合作用研究”(06/07-B18)
作者简介:官凤英(1974-),女,吉林前郭人,助理研究员,博士,从事竹藤资源培育、经营与管理技术研究;*通讯作者。
Diurnal Variations in Photosynthesis of
Calamus nambariensis var. alpinus at Seedling Stage
GUAN Feng-ying,FAN Shao-hui,LIU Ya-di,DENG Wang-hua,PENG Ying
(International Centre of Bamboo and Rattan, The Key Laboratory for Bamboo and Rattan, Academy of Bioresources Utilization, Beijing 100102, China)
Abstract:Diurnal variation in photosynthesis ofCalamus nambariensisvar.alpinusat seedling stage was studied by Licor-6400 portable photosynthetic system. The results demonstrated that 5-monthC. nambariensisvar.alpinusseedling had an obvious double peaks curve of diurnal variation both in net photosynthesis rate(Pn) and transpiration rate(Tr) with obvious midday depression at 12 and the peak value appeared at 10 to 14. For the reason of the stretch of stoma, the increment of Tr was higher than that ofPnafter the midday depression. The diurnal variation trend of water use efficiency(WUE) and stomatal limitation value(Ls) were also double peaked with the peake at 10.WUEreached the second peak at 16. The changing trend of Ls was lower than that ofWUEand got the lowest value at 18. Correlation analysis resulted that Pn had relationships with many environmental factors but not a significant level. The influencing factors to Pn were ordered as follows:Ls>Gs>WUE>Tr>RH>PAR>VPD>Ca>Ta.
Key words:Calamus nambariensisvar.alpinus; net photosynthetic rate; diurnal variation; midday depression of photosynthesis
猜你喜欢
初中生学习指导·中考版(2020年11期)2020-09-10
当代工人(2020年11期)2020-07-06
保健与生活(2020年9期)2020-05-28
汉语世界(The World of Chinese)(2019年5期)2019-11-11
青少年科技博览(中学版)(2018年10期)2018-11-29
科学导报(2018年2期)2018-05-14
新天地(2018年3期)2018-03-21
家庭医药·快乐养生(2017年11期)2017-11-21
作文大王·中高年级(2016年11期)2016-12-09
学生天地·小学中高年级(2015年9期)2015-11-05
