
小鼠早期胚胎发育过程中胚胎发育相关功能基因 mRNA的表达
2010-04-23 03:39柴保国赵卫东郑振宇
河南农业大学学报 2010年6期
周 鑫,柴保国,赵卫东,郑振宇,
(1.河南农业大学生命科学学院,河南郑州 450002;2.河南农业大学牧医工程学院,河南郑州 450002)
“DDK综合症”(DDK syndrome)是 DDK近交系小鼠的特异繁殖现象,同品系交配或 DDK雄鼠能够使其它近交系雌鼠正常受孕,但 DDK雌鼠与同亚种内其它近交系雄鼠交配时,胚胎在发育至妊娠 3~5 d因胚泡形成障碍而死亡,导致 DDK雌鼠几乎不孕[1~3].研究证明,这种极性不育的特性由位于小鼠第 1l条染色体的卵子突变(Ovum Mutant或 Om)位点上的 1对等位基因所控制[4,5].在小鼠早期胚胎发育过程中,主要表达的功能基因有原癌基因、生长因子、生长发育及凋亡基因、白细胞抑制因子等 5大类.其中各个功能基因表达量的变化会对胚胎的正常发育造成不同的影响,并且与 Om基因引发的胚胎致死现象在胚胎学及生理学上极为相似,杂交胚胎与本交胚胎在发育过程中候选基因片段的表达差异,可认为是直接或间接导致胚胎死亡的因素[6~10].本研究运用半定量RT-PCR技术对候选功能基因在表达水平上检测不同交配组合胚胎早期发育过程中存在的表达差异,分析并阐明这些已知基因与 Om基因作用机理的联系,为卵子突变基因作用机理及卵子突变基因导致的不孕症研究提供依据.

表1 候选基因序列及引物Table1 Sequence and primer of candidate gene
1 材料与方法
1.1 试验动物及胚胎
试验所用小鼠为 DDK和 BALB/c品系,每个品系选 120只,雌雄各 60只.DDK小鼠是从日本理化研究所购得,BALB/c小鼠是从河南省实验动物中心购买,试验小鼠进行繁殖扩群获得试验所需样本.采用正常交配方式,收集不同交配组合(试验组:DDK♀ ×BALB/c♂,对照组 1:DDK♀×DDK♂,对照组 2:BALB/c♀×BALB/c♂)小鼠的早期胚胎细胞,并进行观察,2细胞期、4细胞期、8细胞期受精卵各取 90~110枚.PBS缓冲液冲洗,-70℃保存.
1.2 主要试剂及仪器
RNA提取试剂盒购自 TIANGEN公司;反转录试剂盒购自 TaKaRa公司;TaqDNA聚合酶(5 U◦μL-1);10×PCR缓冲液;MgCl2(25 mmol◦ L-1);dNTP(每种 10 mmol◦ L-1);6×电泳上样缓冲液;Marker(DL2000)(大连宝生物公司);紫外分光光度计(尤尼柯仪器有限公司);台式高速冷冻离心机(美国基因公司);梯度 PCR仪(Eppendorf公司);UVP凝胶成像系统(美国 UVP公司).
1.3 引物设计及合成
以候选基因在 NCBI中的 cDNA序列为模板,用 Oligo 6.0软件设计,序列见表 1.
以持家基因 β-actin部分序列为模板,用 Oligo 6.0软件设计 1对引物,进行模板cDNA的质量监控,退火温度 56℃,扩增片段长度 316 bp,引物序列为:F:5'-TGGCATCCATGAA ACTACAT-3',R:5'-AACGCAGCTCAGTAACAGTC-3',由 TaKaRa公司(大连)合成.
1.4 候选基因的 RT-PCR检测
瞬时离心混合均匀,放入预热至 94℃的 PCR仪中,每个候选基因的退火温度如表 1所示.按照试剂盒说明书分别提取不同时期胚胎总 RNA及合成 cDNA,以 cDNA为模板,利用相应引物 PCR扩增目的片段,引物如表 1所示.PCR反应体系:50 μL反应体积,含 10×PCR buffer(含有 Mg2+)5 μL,2 mmol◦ L-1d NTP 4.0μL,上下游引物(10 μmol◦ L-1)各 1.0μL,cDNA 2.5μL和 ExTaqDNA聚合酶(5 U◦ L-1)0.3μL,DEPC水补足至50μL;扩增反应条件为:94℃ 5 min;50℃3 min;72℃ 1 min;1个循环.94℃ 40 s;退火 40 s(退火温度见表1);72℃50 s;40个循环.72℃10 min;4℃保存.扩增产物在含 EB的质量分数为 2%琼脂糖凝胶、0.5×TBE缓冲液,10 V◦ mL-1电泳 50 min检测结果.采用克隆测序法,将鉴定为阳性的质粒送到 TIANGEN公司测序,确定扩增片段的序列.使用 DNAMan软件中的序列相似性分析功能,比较所获得的扩增片段,检查每一个扩增片段的 2到 5个阳性克隆的测序结果是否一致,确定扩增基因片段的准确性.
1.5 半定量分析候选基因的表达情况
同一份样本中分别取 20μL候选基因的扩增产物和 β-actin扩增产物,DNA marker 3μL,用质量分数 2%琼脂糖凝胶电泳检查,在凝胶成像分析系统(UVP)上摄像,并对电泳条带利用图像分析系统 Glyko BandScan 5.0进行灰度分析,以候选基因扩增产物与相应的 β-actin产物的 LOD的比值来表示候选基因 mRNA的相对含量.
1.6 统计分析
采用 SAS 8.0统计软件对各指标进行单因素方差分析,数据用来表示 .
2 结果与分析
2.1 不同交配组合早期胚胎的采集
从不同孕期的怀孕雌鼠中获得不同时期的早期胚胎,选择发育良好、形态正常的胚胎进行试验.一般形态正常的 2细胞期胚胎有 1个清晰可见的极体,或已经将极体完全排除,质地均匀.发育良好、形态正常的分裂期胚胎显示各阶段特异性的细胞分割,具有相等大小的分裂球,无细胞质碎片.通过对不同交配组合早期胚胎细胞的观察,2组对照组交配方式的胚胎能够在 2细胞期之后继续正常发育.但是,试验组交配组合的胚胎细胞在发育至8细胞期后即桑葚期开始,出现大量凋亡现象,胚胎细胞核质弥散,确定为死亡胚胎细胞,无法进行后续试验.因此,胚胎细胞的采集以发育正常的 2细胞期、4细胞期及 8细胞期胚胎细胞为主要对象.分别采集不同交配组合中 3个时期的胚胎细胞进行研究.
2.2 候选基因的 RT-PCR扩增结果
2.2.1 β-actin检测结果 胚胎总 RNA反转录为第 1链 cDNA后,用内参照 β-actin(管家基因)引物进行扩增,通过产物电泳分析,可见明显的 316 bp的片段,说明 RNA反转录效果较好(图 1).2细胞期、4细胞期、8细胞期 β-actin的表达量在试验组与对照组胚胎中基本相同,说明 2种交配组合反转录产物浓度基本一致,可以进行候选基因差异表达研究.
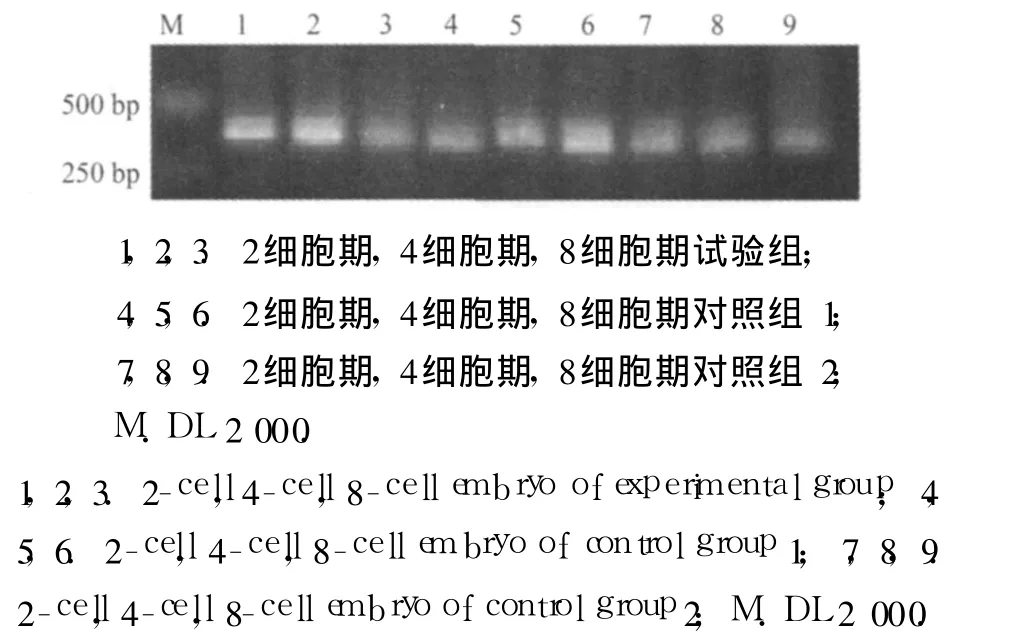
图1 小鼠早期胚胎 2细胞,4细胞,8细胞 cDNA检测结果Fig.1 cDNA of early embryo of mice examined 2-cell,4-cell,8-cell embryo by agarose eletrophoresis
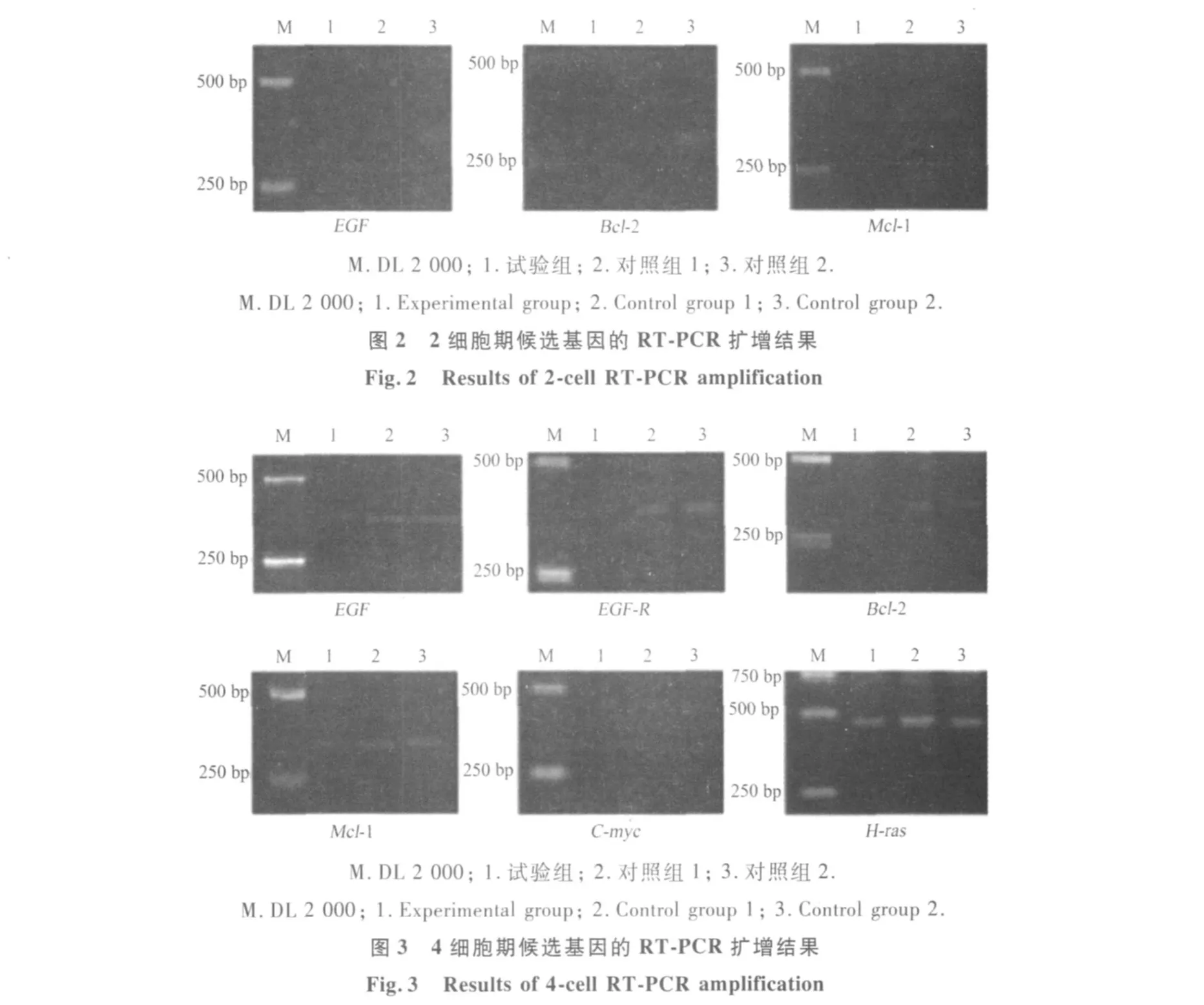
2.2.2 2细胞期候选基因的 RT-PCR扩增 候选基因在 2细胞期胚胎中的扩增产物,利用质量分数2%琼脂糖凝胶进行电泳(图 2).结果表明,在 3种交配组合中只检测到 EGF,Bcl-2,Mcl-1的表达,而其他 5个候选基因都未检测到表达.由于 2细胞期是小鼠胚胎发育过程中,母型调控向合子型调控的过渡阶段,胚胎合子基因组未完全激活.因此,检测到的候选基因表达量较低,电泳条带较弱.
2.2.3 4细胞期候选基因的 RT-PCR扩增 候选基因在 4细胞胚胎中的扩增产物,利用质量分数2%琼脂糖凝胶进行电泳(图 3).结果表明在 3种交配组合中共检测 6个候选基因表达,且表达量较高.由于 4细胞期小鼠胚胎中合子基因组已经激活,因此扩增条带明显.另外,检测到的候选基因中H-ras,C-myc在 3种交配组合胚胎细胞中扩增条带亮度相同;而 EGF,EGF-R,Bcl-2,Mcl-1在试验组组合中扩增条带亮度弱于对照组,说明这些候选基因在试验组胚胎中表达被抑制.
2.2.4 8细胞期候选基因的 RT-PCR扩增电泳结果 候选基因在 8细胞胚胎中的扩增产物,利用质量分数 2%琼脂糖凝胶进行电泳(图 4).其结果与4细胞期相似,在 3种交配组合中仍未检测到 LIF,但是检测到 TGF-α的表达,候选基因扩增条带明显,并且个别候选基因表达量上较 4细胞期有明显提高.其中 TGF-α,C-myc,H-ras在 3种交配组合胚胎细胞中扩增条带亮度相同;而 EGF,EGF-R,Bcl-2,Mcl-1在试验组组合中扩增条带亮度弱于对照组,说明这些候选基因在试验组胚胎中表达量较低.
2.2.5 扩增片段测序结果 将扩增条带回收后送到 TIANGEN公司测序.对测序差异表达片段信息进行统计分析(略).结果表明,扩增片段序列与GenBank公布的候选基因 cDNA序列相比,各个基因扩增结果较好,同源性达 97% ~99%,证明所扩增的片段是候选基因片段.
2.2.6 候选基因的半定量分析 将凝胶成像图片用 BandScan 5.0图像分析系统对候选基因与 βactin电泳结果进行灰度对比分析(每个样本重复 3次),从而显示不同时期候选基因在小鼠胚胎中的mRNA丰度,2细胞期、4细胞期、8细胞期结果分别见表 2~表 4.
从表 2可以看出,候选基因小鼠的 2细胞期胚胎中的表达存在差异.EGF,Bcl-2,Mcl-1 3种候选基因在试验组 2细胞期胚胎中表达量分别与对照组 2细胞期胚胎中表达量差异显著(P<0.05),并且在试验组中表达量明显低于 2个对照组.
从表 3可以看出,在 3种交配组合小鼠 4细胞期胚胎中检测到 6个候选基因,其中部分候选基因的表达存在差异.C-myc,H-ras在 4细胞期小鼠胚胎细胞中表达差异不显著(P>0.05);而 EGF,EGF-R,Bcl-2,Mcl-1在 3种交配组合 4细胞期胚胎中表达差异显著(P<0.05),并且在试验组中表达量明显低于 2组对照组.

图4 8细胞期候选基因的RT-PCR扩增结果Fig.4 Results of 8-cell RT-PCR amplification

表2 2细胞期候选基因在 3种交配组合小鼠胚胎中的 mRNA丰度Table2 Relative contents of candidate gene in 2-cell embryo

表3 4细胞期候选基因在 3种交配组合小鼠胚胎中的 mRNA丰度Table3 Relative contents of candidate gene in 4-cell embryo
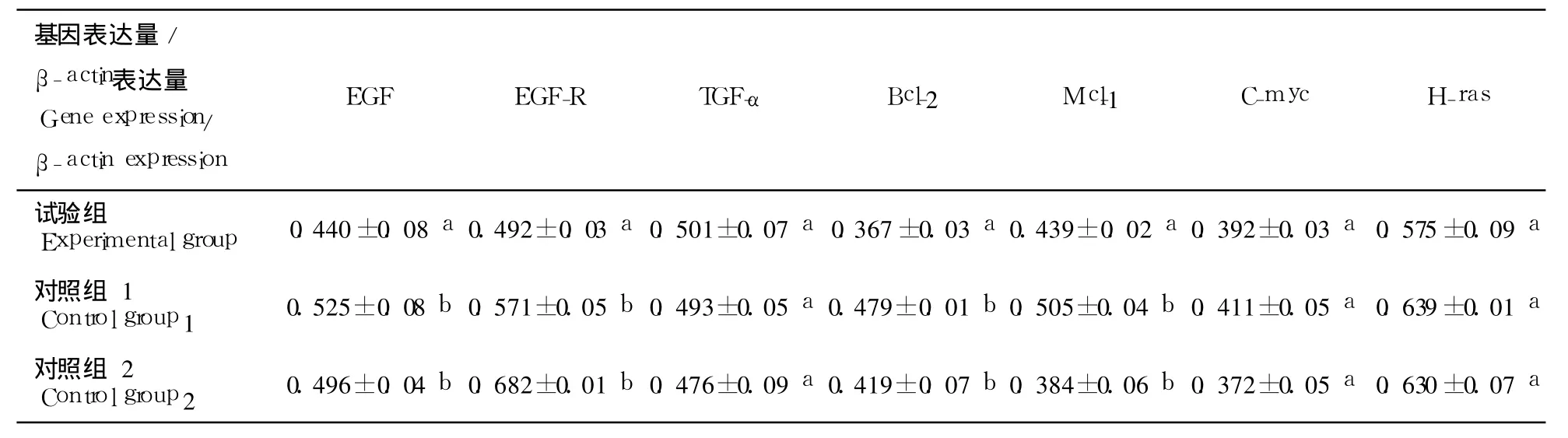
表4 8细胞期候选基因在 3种交配组合小鼠胚胎中的 mRNA丰度Table4 Relative contents of candidate gene in 8-cell embryo
从表 4可以看出,在 3种交配组合小鼠 8细胞期胚胎中检测到 7个候选基因,其中与 4细胞期检测相同的 6个候选基因,在表达上的差异也相同.TGF-α,C-myc,H-ras在 8细胞期小鼠胚胎细胞中表达差异不显著(P>0.05);而 EGF,EGF-R,Bcl-2,Mcl-1在 3种交配组合 8细胞期胚胎中表达量差异显著(P<0.05),并且在试验组中表达量明显低于 2个对照组.
3 小结与讨论
1)基于对 Om基因研究的相关结论分析,Om基因可能是通过母源 RNA分子影响其它早期胚胎发育的功能基因,通过对 DNA(2细胞期精卵基因组发生融合)水平进行的直接修饰或对功能基因RNA水平(成熟 mRNA形成过程中)的间接修饰,使其不能正常表达或者不能正常作用于胚胎发育,进而导致胚胎致死[7].小鼠作为高等哺乳动物,其基因调控具有复杂性,致死性状的产生极有可能是由多基因调控的.本试验通过对小鼠胚胎发育过程中功能基因的筛选分析确定一系列候选基因,这些功能基因均在胚胎发育早期表达,并且其表达情况直接影响胚胎的正常发育.通过对胚胎学及形态学的分析,这些候选基因的异常表达能够导致胚胎致死,并且与 Om基因介导的胚胎致死现象极为相似.
2)EGF和 EGF-R基因属于生长因子,体细胞的生长分化由生长因子调节是早已明确证实的[10],而在早期胚胎发育方面,生长因子可能的作用尚不清楚.小鼠胚胎中,从 4细胞期开始就可测到 EGF-RmRNA,EGF主要通过自分泌或旁分泌的方式作用于胚胎中的EGF-R,调节胚胎的生长发育[11].不同发育时期的小鼠植前胚胎受 EGF的作用也有差异,在 4细胞期前促进卵裂,桑葚胚期以后表现为调节分化.在含有 EGF的培养基中能显著提高囊胚孵化率,且特定浓度的 EGF对胚胎早期发育、着床均起到一定的作用[9].试验证明,EGF,EGF-R在早期胚胎发育、胚胎的分化与增生、胚泡附植等过程中起着重要的调节作用[11~15].TGF-α通过与 EGF共同的受体即 EGF-R,对哺乳动物早期胚胎发育也具有重要的作用.在体外培养液中添加 TGF-α,则小鼠胚胎 mRNA转录加快,蛋白质合成增加,细胞数量增多,胚胎发育速度加快.严云勤等[12]通过 RT-PCR技术检测小鼠早期胚胎中 TGF-α的 mRNA表达,发现 TGF-α在囊胚期作用较大,可能与囊胚腔的形成和扩展有关.
3)Bcl-2和 Mcl-1同属于凋亡基因[9].HARDY[11]在形态正常的人类囊胚中检测到了 Bcl-2的高表达,而在同期碎裂胚胎当中则没有发现.李威等[14]利用巢式 PCR方法检测小鼠早期胚胎发育过程中 Bcl-2家族基因的表达情况,及其与胚胎细胞凋亡的关系.Bcl-2的高表达通常被认为能够抑制细胞的凋亡,它的表达量的升高导致胚胎细胞凋亡比率的降低.随着胚胎发育进程,胚胎细胞数量不断增多,胚胎总体处于快速增殖时期,Bcl-2的表达不断增加有利于胚胎生长和发育.
4)在对候选基因的检测过程中,发现 3种交配方式小鼠早期胚胎发育过程中都没有检测到LIF的表达,在胚胎发育过程中 LIF可能是通过母体子宫或者卵巢分泌机制产生进一步作用于胚胎,而早期胚胎细胞本身不产生 LIF.
5)本试验利用半定量 RT-PCR技术,通过对(DDK♀ ×BALB/c♂),(DDK♀ ×DDK♂),(BALB/c♀×BALB/c♂)3组交配组合小鼠早期胚胎细胞中候选基因表达量差异的检测,证实了Om基因控制的卵子细胞质物质以某种机制作用于 Mcl-1,Bcl-2,EGF和 EGF-R,并通过这些基因表达水平的降低间接地引发早期胚胎致死现象.
[1] WAKASUGI N,TOMITA T,KONDO K.Differences of fertility in reciprocal crosses between inbred strains of mice:DDK,KK and NC[J].Reprod Fertil,1967,13:41-50.
[2] WAKASUGI N.Studies on fertility of DDK mice:reciprocal crosses between DDK and C57BL/6J strains and experimental transplantation of the ovary[J].Reprod Fert,1973,33:283-291.
[3] WAKASUGI N.Embryologic,cytobiologic and genetic interpretations of DDK syndrome in mice[J].Growth Differ,2007,49:555-559.
[4] BALDACCI P A,COHEN-TANNOUDJI M,KRESS C,et al.Highresolution map around the locus Om on mouse Chromosome 11[J].Mamm Genome,1996,7:114-116.
[5] PARDO-MANUEL D E,VILLENA F,NAUMOVA A K,et al.Confirmation of maternal transmission ratio distortion at Om and direct evidence that the maternal and paternal“DDK syndrome” genes are linked[J].Mamm Genome,1997,8:642-646.
[6] ADAMSON E D.Oncogenes in development[J].Molecular Reproduction and Development,1987,99:449-471.
[7] KIM C H,CHAE H D,CHENO Y P,et al.The efect of epidermal growth factor on the preimplantation development,implantation and its receptor expression in mouse embryos[J].JObstet Gynaecol,1999,25(2):87-93.
[8] SHI C Z,DHIR R N,KESAVAN P,et al.Mouse embryonic stem cells express receptors of the insulin family of growth factos[J].Mol Reprod Dev,1995,42(2):173-179.
[9] YOKO H,HIDEO T,JUN F,et al.Experssion of smac/diablo in mouse preimplantation embryos and its corelation to apoptosis and fragmentation[J].Molecular Human Reproduction,2005,11(3):183-188.
[10]HILTOND J,GOUGHN M.Leukaemia inhibitory factor:a biological perspective[J].J Cellular Biochem,1991,46(1):21-26.
[11]HARDY K.Cell death in the mammalian blastocyst[J].Mol Human Reprod,1999,3:919-925.
[12]严云勤,谭景和,张黎霞.小鼠早期胚胎发育过程中TGF-α和 EGF-R mRNA的表达[J].动物学报,2002,48(4):567-569.
[13]李 莉,陈秋阁,王红梅,等.水红花子对小鼠免疫功能及迟发型超敏反应的抑制作用[J].河南师范大学学报:自然科学版,2009,37(4):125-129.
[14]李 威,王连卿,苏静艳,等.小鼠早期胚胎发育过程中细胞凋亡及凋亡基因表达的检测[J].细胞生物学杂志,2006,28(4):596-602.
[15]金 铖,刘慧敏,耿 娟,等.粗提牛白细胞抗菌肽对小鼠免疫功能的影响[J].河南农业大学学报,2009,43(6):630-634.
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
四川蚕业(2021年1期)2021-02-12
中国生殖健康(2020年5期)2021-01-18
昆虫学报(2020年6期)2020-08-06
昆虫学报(2020年1期)2020-03-03
中国生殖健康(2018年5期)2018-11-06
罕少疾病杂志(2017年2期)2017-02-23
广西林业科学(2016年3期)2016-03-16
新疆医科大学学报(2015年10期)2015-12-26
