
不同放牧强度下陇东天然草地土壤微生物三大类群的动态特征
2010-03-31 06:11:54张成霞南志标
草业科学 2010年11期
张成霞,南志标
(1.西南林业大学园林学院,云南昆明 650224;2.兰州大学草地农业科技学院,甘肃兰州 730020)
土壤微生物是土壤物质转化的重要参与者,在维持生态系统平衡,评价土壤健康方面具有不可替代的作用[1]。土壤微生物是草地生态系统的重要组分,其将植物不能直接吸收的难溶性无机物转变为可溶性的,增加土壤肥力,在有机质分解转化、养分的释放和循环及改善土壤理化性质中起着主导作用,能动地影响着草原生态系统的能量流动和物质转化。微生物在土壤中的分布与活动,既直接反映了各环境因素及草地管理利用对微生物的影响和作用,也间接反映了草地的状况[2-5]。放牧对东北羊草(Leymus chinensis)草原[3]、内蒙古冷蒿(Artemisia f rigida)-小禾草草原[6]、荒漠草原[7]、东祁连山高寒草地[8]土壤微生物影响已有文献报道,但有关放牧对黄土高原北部农牧交错带天然草地土壤微生物的影响鲜见报道。本研究通过分析不同放牧强度、不同取样期、不同土层下土壤微生物数量的变化,探索放牧对草地土壤微生物的影响,为黄土高原天然草地的可持续利用提供理论依据。
1 材料与方法
1.1 样地自然概况研究样地位于甘肃省环县甜水镇大梁洼村——兰州大学草地农业科技学院野外试验点。具体简况见已发表的文章[9]。
1.2 放牧试验设计试验区于2001年在天然草地上进行划区围栏,设4个放牧处理,分别为0(对照)、2.7、5.3和8.7只羊/hm2,每放牧处理3个重复,随机排列,每轮牧分区为1个重复,面积为50 m×100 m,根据轮牧分区面积确定每区实际放牧羊数。每年夏季开始放牧,放牧时间为每年6月中旬至9月中旬,每分区每次放牧10 d,共轮牧3次。试验羊为滩羊。至本试验开始取样时,已连续放牧3年。
1.3 土样采集采样时间在牧草返青后,于2005年5月中旬、6月中旬、9月中旬、11月中旬和2006年4月中旬、6月中旬分别采集土样。每轮牧分区均按 0~10、10~20和20~30 cm 3层取样,每样地采用土壤剖面和混合多点取样法,同层混合为一个土样,人工捡除碎石、植物残体等可见物,装入塑料自封袋,4℃低温下运回实验室,未过筛的新鲜土样立刻进行土壤微生物的分离。菌落数计算以干土为基础。
1.4 土壤三大类群微生物的分离及数量测定土壤微生物测定分析采用平板分析法[10]:真菌采用马丁氏-孟加拉红培养基,细菌采用牛肉膏蛋白胨培养基,放线菌采用高氏1号培养基。真菌、细菌和放线菌计数的土壤悬浮液稀释浓度分别为10-2、10-3和10-3,每个土样共涂抹 9个平板,其中每类微生物3个平板,每个平板为1个重复,共3个重复。涂抹后真菌置22℃黑暗培养箱内培养,3~7 d内统计菌落数;细菌置28℃黑暗温箱内培养,逐日观察菌落生长情况,5 d后没有新的菌落长出时计数工作结束;放线菌在30℃黑暗条件下培养,5 d后开始统计菌落数,10 d后没有新的菌落长出时计数工作结束。
1.5 数据统计与分析试验数据用Statistical(1993)软件进行显著性统计分析,并用Excel软件进行图表制作。
2 结果
2.1 土壤微生物总数量2年6次测定平均土壤微生物总数在放牧强度为8.7只羊/hm2的轮牧分区最高,在5.3只羊/hm2的轮牧分区最低(表1)。但逐一比较同一测定日期内,不同放牧率下各轮牧分区土壤微生物总数量发现,仅2005年5月、6月0只羊/hm2显著(P<0.05)低于8.7只羊/hm2,2006年6月5.3只羊/hm2轮牧分区显著(P<0.05)低于 2.7只羊/hm2和 8.7只羊/hm2的处理,其他各测定日期内,不同放牧率处理间差异并不显著(P>0.05)(表1)。

表1 不同放牧强度、不同取样期土壤微生物三大类群总数量 (×104CFU/g)
2.2 各层土壤微生物的动态特征
2.2.1真菌的动态特征 不同放牧强度下,0~10、10~20和20~30 cm土层,真菌数量的变化趋势基本一致。即真菌数量的最高值均出现在9月,最低值在4或11月(图1)。20~30 cm土层,2005年5月,2.7只羊/hm2轮牧分区的真菌数量显著(P<0.05)高于5.3只羊/hm2轮牧分区,高幅达83.50%(图1)。
2.2.2细菌的动态特征 不同放牧强度下,0~10、10~20和20~30 cm土层,细菌的变化趋势一致。3个层次的细菌数量最高值均在9月,最低值均在4月。20~30 cm土层,2005年6月,8.7只羊/hm2轮牧分区的细菌数量显著(P<0.05)高于未放牧的轮牧分区,高幅达81.50%(图2)。
2.2.3放线菌的动态特征 不同放牧强度下,0~10、10~20和20~30 cm 土层,放线菌的变化趋势基本一致。3个层次的放线菌数量最高值均在11月,而最低值在4或6月。2005年5月和2006年6月在 0~10 cm土层,放线菌数量在2.7只羊/hm2轮牧分区显著(P<0.05)高于0和5.3只羊/hm2,高幅达43.69%~180.36%。10~20 cm土层放线菌数量,2005年 5月,在 2.7只羊/hm2的轮牧分区显著(P<0.05)高于 5.3只羊/hm2,高幅达 77.59%。20~30 cm土层放线菌数量,2006年4月,在2.7只羊/hm2轮牧分区显著(P<0.05)高于 0只羊/hm2,幅度达107.04%(图3)。
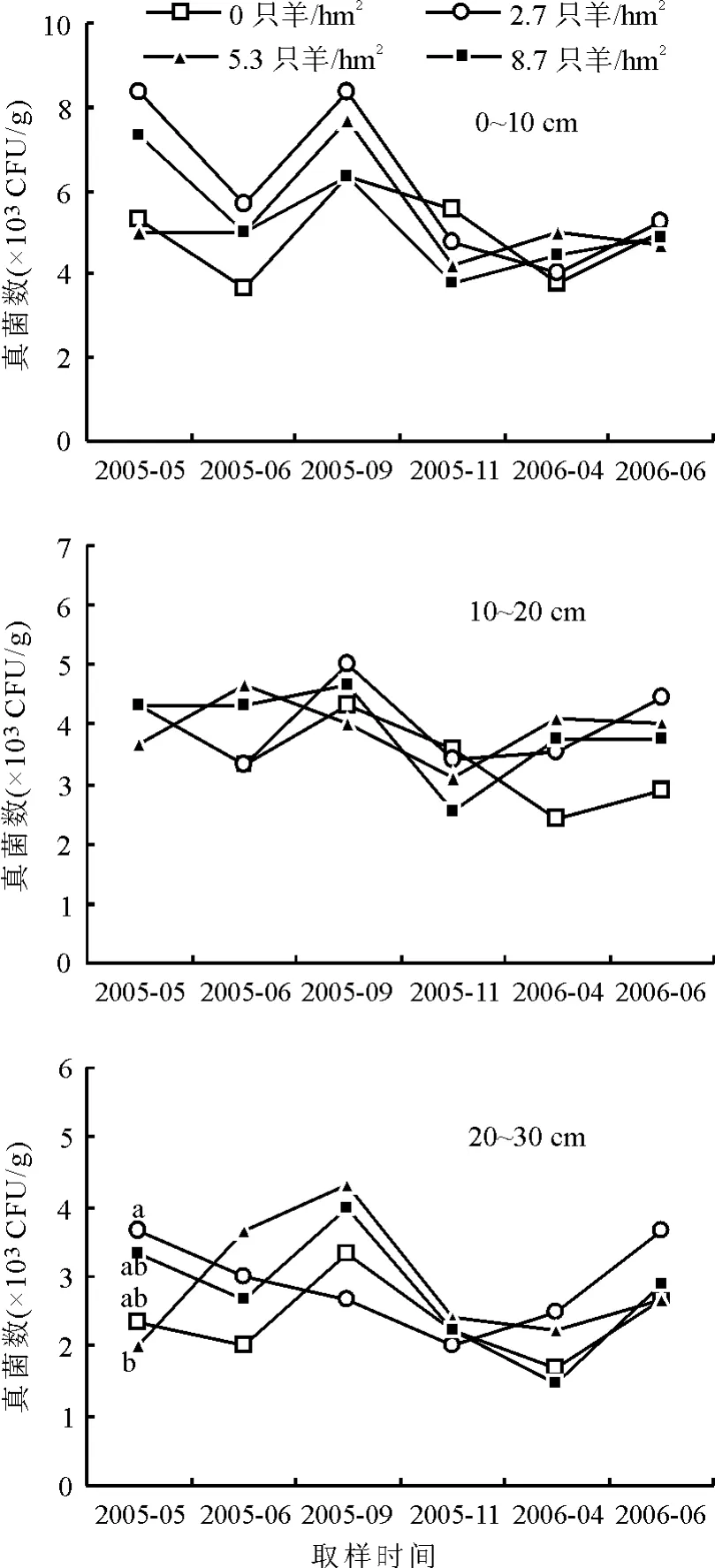
图1 不同放牧强度下土壤真菌的动态
2.3 土壤微生物的空间动态分布2005年,土壤真菌、细菌和放线菌数量均表现为随土层加深呈极显著(P<0.01)的降低。2006年,真菌和放线菌数量亦表现为随土层加深逐渐降低,但细菌数量表现10~20 cm高于0~10 cm。从2年试验结果看出,土壤微生物数量总体表现为表层最高,随土层加深逐渐降低(图4)。
3 讨论
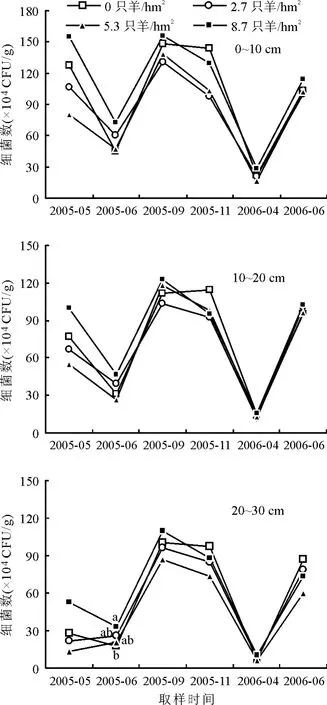
图2 不同放牧强度下土壤细菌的动态
3.1 土壤微生物的数量和比例真菌、细菌和放线菌由于其生态属性不同,它们的数量及其在微生物总数中所占的比例对栖息的生态条件有一定的指示意义,同时,受土壤成分、土壤酸碱度和植被种类的影响,是土壤的重要生物学指标。本研究结果表明,未放牧的对照区和轮牧分区,三大类群微生物数量上,细菌占绝对优势,放线菌次之,真菌最少,说明细菌对该土壤的适应性强,而真菌和放线菌因受土壤中有机质、pH值等环境因子的影响,生长发育受到极大限制。这与以往研究报道的草地土壤微生物数量变化一致[8,11-12]。

图3 不同放牧强度下土壤放线菌的动态
3.2 土壤微生物数量对放牧的响应放牧对土壤微生物数量的影响因草地类型、管理措施等而异,因为各类草原的土壤水热条件、肥力状况及生产力不同。本研究结果表明,同一取样期(6月)不同的年份(2005和2006年)土壤微生物的数量变化差异较大,导致这种现象的原因与2年不同的气候条件有关。2005和2006年1-5月的降水量分别是36.0和34.7 mm,1-5月的平均气温分别为0.62和2.58℃。土壤水分和气温均会影响土壤微生物的活动和数量,加之随着放牧时间的增加,牲畜的踩踏和采食导致土壤板结和植被结构改变;另一方面,牲畜排泄物造成土壤外来微生物的增加和土壤养分的改变,而这些变化都将对土壤微生物造成影响,故2006年6月土壤微生物数量明显高于2005年6月。

图4 2005和2006年土壤微生物的空间分布
不同微生物类群对放牧的响应不一致。本研究结果表明,真菌和细菌对放牧的响应比放线菌更为敏感。土壤真菌和放线菌的数量在2.7只羊/hm2轮牧分区上最高,但随着牧压的增加其值下降,这与赵吉[6]、马爽等[13]的研究结果一致。说明围栏封育和适度放牧均有助于土壤微生物的恢复,提高草地土壤微生物的数量,但过度放牧使土壤微生物活性降低,从而使微生物数量下降。细菌数量在牧压最重的8.7只羊/hm2的轮牧分区最高,这说明牧畜的践踏作用使土壤变得紧实,通气性下降,从而使土壤中好气性细菌随着放牧率的提高逐渐减少,嫌气性细菌数量在一定的放牧率下略有增加[6]。同时,细菌适应不良环境的能力强[14]。因而,细菌数量在牧压最重的处理区最高。
3.3 土壤微生物的季节动态土壤微生物的变化是非常复杂的。土壤微生物的变化具有明显的季节性,这与有机物的供应、植物生长状况以及温湿等环境因素有关[15]。生态环境中的土壤温度、水分等因素伴随着季节变化而变化,是影响土壤微生物数量的主要因素。同一种草地类型的同一块样地,季节变化是导致土壤微生物类群数量分布特征存在差异的原因之一。本试验结果表明,陇东黄土高原天然草地土壤的真菌、细菌和放线菌数量具有明显的季节性变化,土壤真菌和细菌数量均呈单峰曲线,5月到9月上升,9月到11月下降,高峰出现在9月。产生这种变化的原因是:早春土壤温度和水分较低,导致微生物不能进行旺盛地生长繁殖,数量较低。5月随着气温回升,牧草开始返青,微生物生长繁殖亦加快,数量逐渐上升,9月土壤温度高、水分较充足,有利于微生物的生长和繁殖,数量急剧上升,达到全年最高。该时期也是枯枝落叶分解最旺盛的季节,说明微生物对枯枝落叶的分解作用非常强烈。9月以后进入秋季,温度和水分也逐渐下降,随着气温的降低土壤温度下降,微生物的活性也降低,导致其数量也下降。土壤放线菌数量的变化不同于真菌和细菌的变化,其值从5月到11月逐渐升高,最高值在11月,这与马爽等[13]的结果一致。产生这种变化的原因可能与放线菌的生理特征有关,如较耐低温,参与一些难分解的物质[14],也是放线菌对干旱及其他不良环境的适应能力较强导致的。但也有研究表明,草地土壤微生物量的高峰在7或8月,而9月呈下降趋势[15-17],与本试验的结果略有不同,这可能是不同的草地类型、降水量等因素造成的。
3.4 土壤微生物数量的垂直分布土壤微生物数量从上而下呈急剧减少趋势,这与以往研究报道的结果一致[7-8,11-12,16,18]。这是因为地表聚集大量枯枝落叶,土壤结构疏松,给微生物活动提供良好的通气条件,另一方面,表层土壤的水热和通气状况好,利于微生物生长繁殖。土壤微生物的数量垂直分布也反映了草原土壤层次的营养状况。
[1]Smith J L,Paul E A.The significance of soil microbial biomass estimations[M].New York:Marcel Dekker,1990:357-398.
[2]Klemedtson L,Berg P,Clarholm M.Microbial nitrogen transformation in the root environment of barley[J].Soil Biology&Biochemistry,1987,19:551-558.
[3]杨靖春,刘义,郭玲.放牧对羊草草原微生物区系的影响[J].中国草地,1984(2):35-40.
[4]刘世贵,葛绍荣.川西北退化草地土壤微生物数量与区系研究[J].草业学报,1994,3(4):70-76.
[5]张成霞,南志标.放牧对草地土壤微生物影响的研究述评[J].草业科学,2010,27(1):65-70.
[6]赵吉.不同放牧率对冷蒿小禾草草原土壤微生物数量和生物量的影响[J].草地学报,1999,17(3):223-227.
[7]高雪峰,韩国栋,张功,等.放牧对荒漠草原土壤微生物的影响及其季节动态研究[J].土壤通报,2007,38(1):145-148.
[8]张俊忠,陈秀蓉,杨成德,等.东祁连山高寒草地土壤微生物三大类群的时空动态特征[J].水土保持学报,2008,22(5):167-171.
[9]于应文,南志标.畜尿排泄特征及其对草地植被和家畜选择采食的作用[J].生态学报,2008,28(2):0777-0785.
[10]许光辉,郑洪元.土壤微生物分析方法手册[M].北京:农业出版社,1986.
[11]龙章富,刘世贵.退化草地土壤农化性状与微生物区系研究[J].土壤学报,1996,33(2):192-200.
[12]邵玉琴,赵吉,杨劼.恢复草地和退化草地土壤微生物类群数量的分布特征[J].中国沙漠,2004,24(2):223-226.
[13]马爽,杨成德,薛莉,等.陇东黄土高原不同载畜量土壤的微生物季节变化[J].草原与草坪,2004(2):27-30.
[14]Srivastava S C.Measurement of soil microbial biomass provides an early indication of changes in total soil organic matter due to straw incorporation[J].Soil Biology&Biochemistry,1992,24:711-714.
[15]郭继勋,祝廷成.羊草草原土壤微生物的数量和生物量[J].生态学报,1997,17(1):78-82.
[16]陈珊,张常钟,刘东波,等.东北羊草草原土壤微生物生物量的季节变化及其与土壤生境的关系[J].生态学报,1995,15(1):91-94.
[17]邵玉琴,赵吉.内蒙古库布齐油蒿固定沙丘土壤微生物数量的季节动态分布研究[J].中国草地,2000(2):42-45.
[18]丁玲玲,祁彪,尚占环,等.东祁连山亚高山草地土壤微生物功能数量动态及其与土壤环境关系[J].草业学报,2007,16(2):9-18.
猜你喜欢
环球时报(2022-03-29)2022-03-29 17:14:11
今日农业(2020年19期)2020-12-14 14:16:52
国际呼吸杂志(2019年22期)2019-12-09 09:20:36
农民致富之友(2018年20期)2018-11-25 15:35:58
知识经济·中国直销(2018年7期)2018-07-27 02:49:52
中国农业科学(2017年8期)2017-05-03 09:02:42
甘肃畜牧兽医(2016年12期)2016-09-21 01:37:12
电测与仪表(2015年8期)2015-04-09 11:50:16
电测与仪表(2015年7期)2015-04-09 11:40:16
应用海洋学学报(2014年4期)2014-11-22 07:43:54
