
干旱胁迫对4种观赏草枯叶率及生理指标的影响
2010-03-31 06:11:26李秀玲杨志民
草业科学 2010年11期
李秀玲,刘 君,杨志民
(1.南京农业大学草坪研究与开发工程技术中心,江苏南京 210095;2.广西药用植物园,广西南宁 530023)
观赏草是我国园林界新兴的新型景观植物,因其自然朴实、观赏价值高且耐旱、耐贫瘠、抗病虫害、养护成本低等优点而倍受推崇[1-2]。近年来,观赏草在我国各地的引种、推广应用研究越来越受到重视,先后开展了国外优良观赏草种的引种、评价、筛选及推广和本地草种选育工作[2-5]。当前城市水资源严重匮乏,城市园林绿地用水与居民用水矛盾日益加重,因而专家学者提出了建设城市可持续旱景园林的构想[6-7],以缓解城市绿地用水困难等问题,同时,具有美化绿化城市环境作用,为抗旱观赏草提供了广阔的应用前景。然而,到目前为止,大部分文献只是笼统地说明观赏草具有较强的耐旱性,而定量的实验研究则较少[8-10]。此外,观赏草种类繁多,待研究范围较广。因此,本研究对4种常见观赏草在干旱胁迫下的形态和生理变化规律进行了探讨,并对其抗旱性进行评价,以建立一种快速、有效的观赏草耐旱性评价体系,为观赏草耐旱资源的筛选和园林应用提供理论依据。
1 材料与方法
1.1 试验材料供试的4种多年生观赏草均由南京农业大学芳华园艺中心提供,为分根容器苗,苗龄3个月。分别为禾本科的红叶白茅(Imperata cylindrica‘Red Baron')、花叶芒(Miscanthussinensis‘Variegatus')、细茎针茅(Stipa tenuissima)和莎草科的金叶苔草(Carex oshimensis‘Evergold')。
1.2 试验设计试验于2008年5月在南京农业大学牌楼试验地进行,采用单因素试验设计,盆栽控水处理。将长势一致的观赏草移栽至高20 cm、口径24 cm的白色聚乙烯塑料花盆中,盆土按泥炭和园土体积比为1∶1进行配置,土壤含有机质1.610%,pH值为6.01。每盆1株,每种草各20盆,缓苗后待测。在南京自然气候条件下遮雨栽培,保持与外界通风,8月开始进行干旱胁迫试验。每个草种各设对照组10盆,处理组10盆。处理组以正常浇水后1 d作为干旱胁迫基点,进行干旱胁迫处理;对照组正常浇水管理。每4 d观察记录植株失水形态,同时,取功能叶测定各项生理指标,3次重复,取平均值。叶片取样时间统一在8:00-8:30进行,试验期间试验地温度见图1。

图1 试验期间试验地温度
1.3 测定指标和方法
枯叶率:50%面积出现干枯症状的叶片数占总叶片数的百分比[8]。
叶片相对含水量:采用组织烘干法[7],即称取叶片鲜质量(Wf),然后将叶片放入蒸馏水中浸泡24 h,取出擦干称取饱和质量(Wt),置烘箱中105℃下杀青15 min,然后于80℃下烘 48 h,称干质量(Wd)。

质膜透性:采用电导率仪法[11]。称取0.1 g叶片加20 mL去离子水,用抽气泵抽气直至叶片完全下沉,用DDS-12A型电导率仪测定其电导率,然后再放入沸水中水浴20 min以杀死组织,冷却至室温后再测定溶液的电导率,用相对电导率表示质膜透性。

丙二醛(MDA)含量:采用巴比妥酸比色法测定[12]。
超氧化物歧化酶(SOD)活性:采用氮蓝四唑(NBT)比色法测定[12]。
可溶性蛋白含量:采用考马斯亮蓝G-250染色法测定[12]。
叶绿素含量:采用李合生方法测定[12]。
1.4 数据分析数据采用 Excel 2003软件处理,差异比较采用SPSS 16.0邓肯新复极差法(SSR法)。
2 结果与分析
2.1 干旱胁迫对枯叶率的影响比较干旱胁迫后枯叶率的变化,可以明显看出处理组4种观赏草的枯叶率均显著上升(图2),而对照组草种的枯叶率变化幅度均较小。从进程来看,处理组红叶白茅在干旱胁迫的第9天枯叶率急剧上升,至胁迫第13天枯叶率高达83.01%,增幅最大;其次为花叶芒和细茎针茅,胁迫末期枯叶率分别为63.00%和42.00%;金叶苔草枯叶率的上升幅度最小,胁迫末期枯叶率仅为10.00%。干旱胁迫期间枯叶率上升幅度由高到低依次为:红叶白茅、花叶芒、细茎针茅和金叶苔草。结果表明,同一草种随着干旱胁迫时间的推进枯叶率呈极显著差异(P<0.01),不同草种在干旱胁迫的第5、9、13天枯叶率均呈显著差异(P<0.05)。

图2 干旱胁迫下4种观赏草枯叶率的变化
2.2 干旱胁迫下植株的生理响应
2.2.1叶片相对含水量(RWC) 在干旱胁迫下,植物首先表现出来的是体内含水量的下降[13-14],植物组织在干旱条件下RWC下降的主要原因可能是土壤含水量下降及植物对水分吸收显著降低。由图3可知,对照组草种的叶片RWC变化较小。处理组不同草种的叶片RWC呈现不同程度的下降趋势,以胁迫第5-9天下降最快,之后趋于平缓。干旱胁迫第9天时,红叶白茅、花叶芒、细茎针茅和金叶苔草的 RWC分别为33.39%、44.34%、45.46%和69.24%,与第 1天相比,下降幅度依次为 63.30%、50.59%、45.46%、21.91%。结果表明,同一草种第9天时的叶片RWC与第5天时比较呈极显著差异(P<0.01)。胁迫末期(第 13天),红叶白茅、花叶芒、细茎针茅和金叶苔草的叶片RWC依次为26.50%、30.21%、38.78%和45.35%,较第 1天的下降幅度为红叶白茅>花叶芒>细茎针茅>金叶苔草。综上分析,金叶苔草的RWC下降最为缓慢,组织保水能力较强,显示出较强的抗旱能力;红叶白茅叶片RWC降幅最明显,抗旱性最弱;花叶芒和细茎针茅居于两者之间。
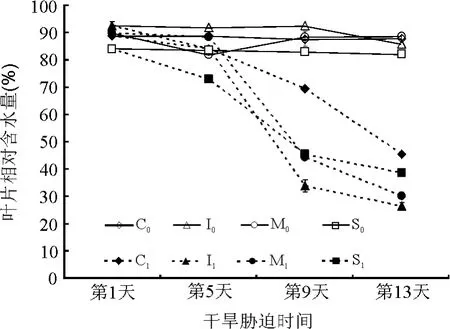
图3 干旱胁迫下4种观赏草叶片相对含水量的变化
2.2.2叶片细胞相对电导率(REC) 植物受到逆境伤害时细胞膜发生膜脂过氧化作用,膜脂由液晶态转变为凝胶态,从而导致膜流动性下降,透性增加,细胞内物质外渗,REC增大,细胞功能下降。干旱胁迫条件下,REC升高,说明叶片的质膜受到了伤害,引起细胞内含物质的外渗,而且REC随干旱胁迫程度的加深不断升高[15-16]。干旱胁迫下不同草种细胞内溶质外渗量均呈上升趋势(图4),随着干旱胁迫的加重,细胞内溶质外渗量增加,膜的受害程度加重。4种观赏草的REC均于胁迫5 d后变化最为明显,干旱胁迫末期,红叶白茅和金叶苔草的REC分别为89.67%和48.76%,分别为溶质外渗率最高草种和最低草种。红叶白茅、花叶芒、细茎针茅和金叶苔草相对电导率的增幅依次为76.98%、55.92%、42.37%和36.02%。
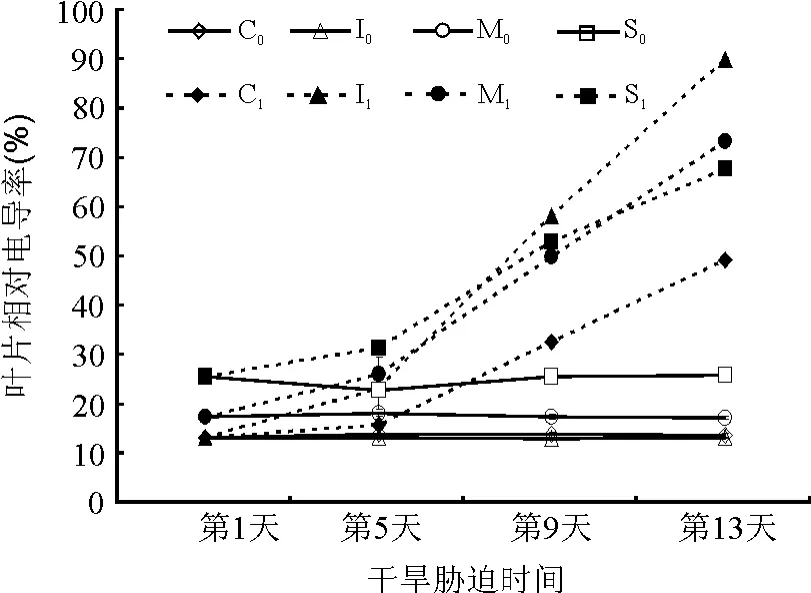
图4 干旱胁迫下4种观赏草叶片相对电导率的变化
2.2.3叶片丙二醛(MDA)含量 MDA是膜质过氧化作用的最终产物,是膜系统受伤害的重要标志之一[17]。MDA积累越多,表明组织的保护能力越弱。从图5可以看出,对照组MDA含量变化较小;处理组草种MDA含量随着胁迫程度的加重而增高,质膜受到不同程度的破坏。胁迫前5 d,MDA含量变化平缓,随后各草种均呈急剧增加趋势,其中以红叶白茅增幅最大,MDA由初期的34.26上升至胁迫末期的87.96 mmol/g,且其MDA含量的基点较高,这可能是红叶白茅特殊的颜色(胁迫及衰老时呈血红色)及其变化所致。花叶芒和细茎针茅MDA含量变化趋势基本一致,干旱胁迫末期MDA含量分别为36.91和35.86 mmol/g,分别上升了 27.58和 28.34 μ mol/g。金叶苔草MDA含量积累最小,变化最为缓慢。结果表明,同一草种随着干旱胁迫程度的加重MDA含量差异显著(P<0.05)。MDA含量增幅由高到低依次为红叶白茅、花叶芒、细茎针茅、金叶苔草。
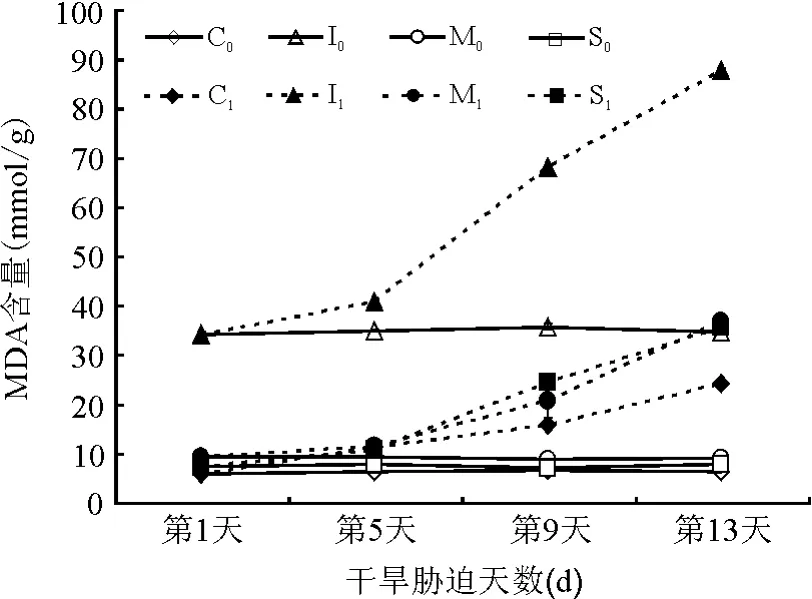
图5 干旱胁迫下4种观赏草叶片MDA含量的变化
2.2.4叶片超氧化物岐化酶(SOD)活性 SOD活性的高低是植物抗逆性的重要标志之一,它可以将超氧阴离子自由基歧化成H2O2,在保护系统中处于核心地位[18]。图6表明,对照组草种SOD活性变化不大,处理组草种SOD活性呈现不同的变化趋势。金叶苔草和细茎针茅SOD活性一直升高;红叶白茅SOD活性呈先上升后下降趋势,这可能是胁迫至第5天时,SOD活性达到最高限度,清除超氧阴离子自由基的能力达到顶峰,随后开始下降,胁迫末期SOD活性与初期SOD活性呈极显著差异(P<0.01);花叶芒SOD活性在胁迫第5天达峰值,随后开始下降,但是花叶芒下降的幅度较红叶白茅缓和,且末期SOD活性与初期相比差异不大。这表明干旱胁迫下红叶白茅和花叶芒叶片的SOD保护酶系统被破坏,其活性明显受到抑制,且受抑制的程度为红叶白茅>花叶芒。

图6 干旱胁迫下4种观赏草S OD活性的变化
2.2.5叶片可溶性蛋白含量 植物在干旱逆境下体内会主动积累一些渗透调节物质,如可溶性糖、可溶性蛋白等。由图7可知,随着干旱时间的推进,4种观赏草可溶性蛋白含量均呈现上升趋势,不同草种上升幅度不同。细茎针茅上升幅度最大,其次为红叶白茅、花叶芒、细茎针茅,金叶苔草的上升幅度最小。结果表明,同一草种随着干旱胁迫时间的延长,可溶性蛋白含量均呈显著差异(P<0.05)。说明4种观赏草受到干旱胁迫后,大量胁迫蛋白被诱导,从而降低了细胞的渗透势,以利于观赏草在干旱逆境下维持正常生长所需水分。
2.2.6叶绿素含量 由图8可知,与对照组相比,处理组4种观赏草叶绿素的含量出现了不同的变化趋势,红叶白茅和花叶芒的叶绿素含量均呈下降趋势,在胁迫的第9天降幅最大;而细茎针茅和金叶苔草都呈现先上升后下降的变化趋势,均于胁迫第5天出现上升趋势。红叶白茅在干旱胁迫末期叶绿素含量降幅最大,其次为花叶芒和细茎针茅,金叶苔草降幅最小。可见,红叶白茅和花叶芒对干旱胁迫较为敏感,在胁迫第5天叶绿素呈现下降趋势;而细茎针茅和金叶苔草在干旱胁迫第5天时叶绿素呈上升趋势,干旱使叶绿素迅速积累,抗逆境能力增强,随着干旱胁迫时间的延长,叶绿素含量随之下降。

图7 干旱胁迫下4种观赏草叶片可溶性蛋白含量的变化

图8 干旱胁迫下4种观赏草叶片叶绿素含量的变化
2.3 枯叶率与生理指标的相关性分析对4种观赏草在干旱胁迫下的枯叶率和生理指标进行相关性分析(表 1)。叶片 REC、MDA含量、可溶性蛋白含量与枯叶率之间均具有极显著的正相关性;叶片RWC、叶绿素含量与枯叶率之间具有极显著的负相关性;各草种SOD活性与枯叶率的相关性差异较大,其中红叶白茅和花叶芒的SOD活性分别与其枯叶率呈显著负相关,而细茎针茅和金叶苔草的枯叶率与其SOD活性呈显著正相关。其余生理指标均与枯叶率呈现出较强的一致性。说明枯叶率可作为观赏草耐旱性的形态判断指标之一。

表1 自然失水胁迫下4种观赏草的枯叶率与各生理指标的相关系数
3 讨论与结论
耐旱性形态指标具有简单易测的优点,反应植物在遭受干旱胁迫后植株的整体表现[8]。本研究主要采用枯叶率作为衡量观赏草在自然失水胁迫下的形态变化。研究结果表明,红叶白茅对干旱胁迫最为敏感,其次为花叶芒和细茎针茅,金叶苔草的枯叶率增加量最少,对干旱胁迫最不敏感。可见,枯叶率能够从表观上较好的反应植物的抗旱性,可作为观赏草耐旱性的形态判断指标,这与张智等[8]研究结果一致。
一般认为,在干旱胁迫下,抗旱能力强的草种保护酶活性较高,且抗性强的草种SOD活性降低比抗性弱的品种缓慢[19]。由以上分析可以看出,虽然金叶苔草SOD活性较低,但是在干旱胁迫期间其活性一直升高,说明其抗旱能力较高,其次为细茎针茅。花叶芒和红叶白茅均呈现先上升后下降的趋势,且两草种干旱末期SOD活性明显低于对照,红叶白茅SOD活性下降幅度最大。
干旱胁迫可导致植物叶片中叶绿素含量降低。叶片缺水不仅影响叶绿素的生物合成,还会促进已形成的叶绿素加速分解,导致叶片发黄,而在一定范围内叶绿素含量的高低则直接影响叶片的光合能力[20],通过对叶绿素含量的测定可以反映出植株对干旱的耐受能力[21-22]。本试验研究结果表明,红叶白茅和花叶芒叶绿素含量先升高后下降,而细茎针茅和金叶苔草叶绿素含量呈一直升高趋势,可见适度的干旱胁迫(1~5 d)可以激活植株的光合性能,促进观赏草的生长,表现为叶绿素含量的增加。而观赏草在过度干旱胁迫(5~9 d)下表现不尽相同,其中红叶白茅和花叶芒叶片过度干旱胁迫下容易失水,叶绿素分解加速,抑制光合作用;而5~9 d的过度干旱胁迫仍然可以提高细茎针茅和金叶苔草的光合性能,叶片叶绿素含量反而增加,表现出较强的抗旱性。这可能与细茎针茅和金叶苔草的根系分布广、扎根深的生长特点有关,干旱胁迫促进了根系的生长和活性的增加,间接地提高了植株对深层养分和水分的吸收,表现出较强的生长势。
综合比较形态和生理指标后,得到4种观赏草的抗旱性大小依次为金叶苔草>细茎针茅>花叶芒>红叶白茅。
[1]Darke R.The Color Encyclopedia of Ornamental Grass[M].Portland,Oregon:Timber Press,1999.
[2]高鹤,刘建秀,郭爱桂.南京地区观赏草的适应性和利用价值初步评价[J].草业科学,2008,25(8):131-138.
[3]武菊英,藤文军,王庆海.耐旱多年生观赏草春季观赏性评价[J].华北农学报,2006,21(1):129-132.
[4]武菊英,藤文军,王庆海.狼尾草的生物学特性及在园林中的应用[J].中国园林,2005,21(12):57-59.
[5]高鹤,刘建秀.南京地区观赏草的种类,观赏价值及其造景配置[J].草原与草坪,2005(3):13-16.
[6]Llewellyn M,Jerry C.Xeriscape ornamental perennial grass trial for low water use landscaping[A].Larson.2002 Annual Report[R].Dickinson:Dickinson Research Extension Center,2002.
[7]武菊英,王国进.可持续旱景园林与观赏草[J].科技潮,2003(10):42-43.
[8]张智,夏宜平,常乐,等.3种观赏草在自然失水胁迫下的生理变化与耐旱性关系[J].东北林业大学学报,2007,35(12):17-20.
[9]张智,夏宜平,徐伟韦.两种观赏草的自然失水胁迫初步研究[J].园艺学报,2007,34(4):1029-1032.
[10]孔兰静,李红双,张志国.三种观赏草对土壤干旱胁迫的生理响应[J].中国草地学报,2008,30(4):40-44.
[11]邹琦.植物生理生化实验指导[M].北京:中国农业出版社,1995.
[12]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[13]马祎,王彩云.几种引进冷季型草坪草的生长及抗旱性生理指标[J].草业科学,2001,18(2):57-61.
[14]孙启忠.四种冰草幼苗抗旱性的研究[J].中国草地,1991(3):29-32.
[15]孟林,毛培春,张国芳,等.17个苜蓿品种苗期抗旱性鉴定[J].草业科学,2008,25(1):21-25.
[16]余书文,汤章城.植物生理与分子生物学[M].北京:科学出版社,1998:366-389.
[17]陈少裕.膜脂过氧化对植物细胞的伤害[J].植物生理学通讯,1991,27(2):84-90.
[18]韩蕊莲,李丽霞,梁宗锁,等.干旱胁迫下沙棘膜脂过氧化保护体系研究[J].西北林学院学报,2002,17(4):1-5.
[19]杜建雄,孙吉雄,刘金荣,等.5个草地早熟禾品种在干热胁迫下的生理响应[J].草原与草坪,2007(3):41-44.
[20]张永强,毛学森,孙宏勇.干旱胁迫对冬小麦叶绿素荧光的影响[J].中国生态农业学报,2002,10(4):13-15.
[21]沈艳,谢应忠.干旱对紫花苜蓿叶绿素含量与水分饱和亏缺的影响[J].宁夏农学院学报,2004,25(2):25-28.
[22]夏阳.水分胁迫对果树脯氨酸和叶绿素含量变化的影响[J].甘肃农业大学学报,1993,28(1):26-31.
猜你喜欢
散文选刊·下半月(2023年5期)2023-05-23 05:09:41
今日农业(2021年16期)2021-10-14 07:43:24
阅读(低年级)(2020年11期)2020-12-28 02:26:35
智慧少年·故事叮当(2020年9期)2020-10-09 10:21:44
读者·校园版(2019年20期)2019-10-18 01:24:32
少年漫画(艺术创想)(2018年7期)2018-11-19 03:00:26
读者·校园版(2018年22期)2018-11-01 03:29:30
现代园艺(2018年3期)2018-02-10 05:18:20
快乐语文(2017年9期)2017-06-15 20:28:36
读者·校园版(2016年17期)2016-08-25 16:39:40
