
重组凝乳酶摇瓶发酵条件的初步优化研究
2009-01-15 09:03张桂芝武彬黄保华刘雪兰石天虹井庆川刘雪兰魏祥法
山东农业科学 2009年12期
张桂芝 武 彬 黄保华 刘雪兰 石天虹 井庆川 刘雪兰 魏祥法
摘 要:凝乳酶是一种酸性蛋白酶,用作动物饲料添加剂,可提高动物特别是幼龄动物对饲料中蛋白成分的消化吸收率,促进其生长。本试验研究了由毕赤酵母胞外表达多拷微小毛霉凝乳酶的摇瓶发酵条件。确定最佳表达条件为:甲醇每24 h添加1.0%,无氨基酵母氮源培养基(YNB)添加1.5%,采用pH 6.0的磷酸缓冲液,诱导120 h,在此条件下,重组凝乳酶的产量可达6 500 IU/ml。
关键词:凝乳酶;发酵条件;毕赤酵母
中图分类号:S816.79 文献标识号:A 文章编号:1001-4942(2009)12-0081-04
凝乳酶(EC3.4.23.4)是一种酸性蛋白酶,能促进蛋白质凝结,用作动物饲料添加剂,可提高动物特别是幼龄动物对饲料中蛋白成分的消化吸收率,促进其生长[1]。幼龄动物的消化系统发育还不完善,对饲料中营养的消化吸收率较低。提高饲料中蛋白类的利用效率,不仅能够节约饲料资源,降低养殖成本,而且能够降低动物粪便的氨氮排放,减轻集约化饲养对环境的污染[2]。
巴斯德毕赤酵母(Pichia pastoris)表达系统是一种比较理想的真核微生物表达系统,是用于表达重组蛋白的标准工具之一,近年发展和应用较多[3]。Pichia pastoris具有与真核生物极其相似的分泌途径和很强的真核蛋白质修饰功能,其自身分泌的蛋白质很少且易于高密度发酵,因此在表达和分离纯化异源蛋白质等方面具有很强的优势。但巴斯德毕赤酵母表达系统也会出现分泌蛋白聚合、降解等现象,从而降低蛋白的生物活性和产量[4]。此外,酵母细胞的生理状态也会严重影响其产酶活性,培养基的pH和甲醇、无氨基酵母氮源培养基(YNB)等营养物质浓度,对细胞的代谢状态会产生很大影响。本研究室成功地将微小毛霉凝乳酶在毕赤酵母中表达[5]。本试验对重组凝乳酶发酵培养基的pH值、甲醇和YNB浓度,以及接种比例等发酵条件进行了优化,提高了重组凝乳酶的产量。
1 材料与方法
1.1 材料
菌种:具有12拷贝基因的整合重组毕赤酵母菌PIC9K-mcp-03。
培养基:BMGY/BMMY培养基:取酵母提取物10 g,蛋白胨20 g,溶于700 ml去离子水中,高压灭菌20 min,冷却至室温,加入1 mol/L磷酸钾缓冲液(pH 6.0)100 ml、10×YNB 100 ml、500×B 2 ml、10×GY 100 ml(若制备BMMY,用100 ml 10×M代替10×GY),4℃保存备用(如无特殊说明,均采用上述条件,改变其中某项发酵条件时,其它项目不变)。1 000 ml三角瓶中加入100 ml培养基,于29℃ 170 r/min振荡培养5 d,每24 h补加甲醇1次。
1.2 方法
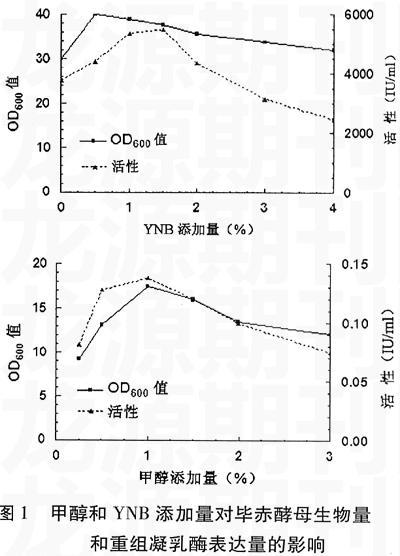
1.2.1 甲醇和YNB添加量对发酵液的影响 改变BMMY培养基中甲醇(0~3%)和YNB(0~4%)的添加量,其它条件不变,培养120 h,测定发酵液OD600 nm和发酵上清液酶活力。
1.2.2 接种量对重组凝乳酶产量的影响 改变BMMY培养基的接种量,选择不同体积的BMGY培养获得的菌体细胞,加入BMMY培养基中培养120 h,测定发酵液OD600 nm和发酵上清液酶活力。
1.2.3 BMMY培养基的初始pH值对重组凝乳酶产量的影响 改变BMMY培养基中所含磷酸缓冲液的pH值,分别调至pH 4.0~7.0,培养120 h,测定发酵液OD600 nm和发酵上清液酶活力。
1.2.4 菌体浓度的计算方法 将发酵液离心,菌体沉淀以蒸馏水重悬,经适当稀释后,测定并计算菌体悬浊液在600 nm的OD值。
1.2.5 酶活测定采用Arima方法[7] 适当稀释的上清液0.5 ml 35℃保温5 min,加入到35℃预保温10 min的5 ml 10%脱脂牛乳中,计时至管壁出现颗粒。记录凝乳时间(T)。酶活定义:在一定温度下(35℃),40 min凝乳1 ml牛乳的酶量为一个酶活单位。
凝乳酶活力(U)=[(5×2400)/(0.5×T)]×n,其中:T为凝乳时间,n为稀释倍数。
2 结果与分析
2.1 甲醇和YNB添加量对重组凝乳酶产量的影响
甲醇和YNB在培养基中起提供营养的作用,甲醇除了是发酵培养基的碳源,还是外源蛋白表达的诱导物,其浓度对重组凝乳酶的表达有一定的影响,随着甲醇浓度的增加重组凝乳酶的表达量也增加。而YNB除了作为培养基的氮源,还含有维生素和多种无机盐类。但二者在较高浓度下,又对发酵有抑制作用,且高浓度的甲醇对酵母菌也有毒害作用,甲醇经代谢生成甲醛和甲酸,甲醛可与带有羟基、巯基、氨基基团的分子发生亲核加成反应,向其提供亚甲基,使自由的分子链被甲醛交联起来,失去原有的生理功能,因此会影响重组凝乳酶的表达量。而YNB中含有的(NH4)2SO4、K3PO4、和NaCl、CaCl2等无机盐在较高浓度下,同样会影响毕赤酵母的生长代谢,降低重组凝乳酶的表达量。培养基中YNB和甲醇的添加量对重组凝乳酶的影响如图1所示。结果表明甲醇浓度在1%、YNB浓度为1.5%时,毕赤酵母的生物量与重组凝乳酶的产量达到最高值。
2.2 接种量对重组凝乳酶产量的影响毕赤酵母的接种量实际上是有可能表达重组
凝乳酶的细胞数,这些酵母是否表达目的蛋白,还受多种环境因素的影响。除了底物浓度之外,实际上培养基中的溶氧量具有重要的影响。图2为接种量影响重组凝乳酶表达量的结果。随接种量的增加,菌体生物量与酶活力的变化情况并不一致,酶活力在接种量为20%时达到最高值,25%也保持较高水平,之后略有下降;而菌体生物量在接种量为30%时达到最高,之后迅速下降。因此,溶氧量很可能是毕赤酵母表达的主要限制性因素。因为溶氧量受多种因素影响,改变摇床的转速、培养基成分等都会改变溶氧浓度,并导致表达量的改变。因此,最佳接种量的测定会随上述条件改变而略有波动,一般在16.7%~25.0%之间。

2.3 培养基的初始pH值对重组凝乳酶产量的影响
改变培养基的pH值对细胞的活性也会产生很大影响。重组凝乳酶在pH 3.2~8.0都比较稳定,因此发酵培养基可选的pH值范围较广。根据酵母的代谢特点,测定了初始培养基在pH 4.0~7.0毕赤酵母的生物量和凝乳酶的表达量(图3)。在发酵过程中酵母会产酸,因此改变了加入培养基的磷酸缓冲液的pH值。

酵母表达系统中外源蛋白的降解失活和不同pH值对酵母生理状态的影响,是导致重组凝乳酶表达量的两项主要因素。图3的结果显示,在pH 4.0~7.0范围内,凝乳酶的活力与菌体浓度变化基本一致,推测可能是由于重组凝乳酶在不同pH值下稳定性较强,且凝乳酶自身作为蛋白酶具有较强的蛋白酶水解抗性。
3 讨论与结论
发酵时间、营养和诱导物的添加量、接种量以及培养基初始pH值对重组凝乳酶的产量都有显著影响,但上述影响只是表观现象。实际上,这些因素的影响更深入地体现在对酵母的生理状态和重组凝乳酶的特定影响两个方面。
甲醇作为发酵的诱导物和碳源,而YNB作为氮源,在一定浓度下对重组凝乳酶的表达具有促进作用,但二者在较高浓度下,又对发酵有抑制作用。实际上不论是甲醇还是YNB对毕赤酵母生物量和表达量的影响并不一致。在低浓度下,甲醇主要表现诱导作用,而只表现出较弱的作为碳源的作用。因此,甲醇添加量为0.5%时,尽管OD600nm表征的菌体量较低,却获得了较高的酶活力。而在添加量为0.5%时,YNB作为氮源则主要表现出对菌体的促进生长作用,只有在满足了菌体生长需要后,在添加量为1.0%~1.5%时,才表现出促进重组蛋白分泌的作用。
接种量以及营养物添加浓度导致重组凝乳酶产量的变化,从根本上讲,体现的都是酵母生理状态对重组蛋白分泌量的影响。OD600 nm所表征的菌体浓度与蛋白分泌量的差别正说明了这一点。以甲醇为碳源,需要消耗大量的溶解氧,提高接种量,菌体浓度增加,势必降低单位细胞可利用的溶解氧含量,从而降低单位细胞重组蛋白的表达量。
酸碱度对蛋白表达量的影响主要体现在两个方面:首先,发酵液初始pH值将影响酵母细胞的生理状态;其次,发酵液的pH值还对表达蛋白的稳定性有一定的影响。因为凝乳酶具有较强的pH值稳定性,而且该酶本身为真菌胞外蛋白酶,对其它蛋白酶的水解具有一定的抗性,所以,图3显示的培养基初始pH值对毕赤酵母生物量和表达量的影响比较一致,即本试验中,发酵液初始pH值影响重组凝乳酶表达量,主要是源于对酵母细胞生理状态的影响。
上述各种因素对酵母细胞的生理状态具有不同的影响机理,各种影响作用相互交叉、十分复杂,而正交试验设计要求各个影响因素的作用彼此相互独立。因此,本试验的各项影响因素不宜简单套用正交试验进行最佳发酵条件的优化,而试验中获得的发酵条件还有进一步优化的空间。
参 考 文 献:
[1] 肖 竞,孙建议,李卫芬.酸性蛋白酶及其在畜牧业中的应用[J].饲料博览, 2003, 3:27-29.
[2] 费笛波,钱玉英,叶玲玲,等. 黑曲霉菌株6042饲用酸性蛋白酶的性质及其应用研究[J].浙江农业学报,1997, 9(6):300-304.
[3] Hao Y Y, Chu J,Wang Y H,et a1.Expression and aggregation of recombinatant human consensus interferon-α mutant by Pichia pastoris[J]. Biotechnology Letters, 2006, 28(12):905-909.
[4] Macauley-Patrick S,Fazenda M L,McNeil B,et a1.Heterologous protein production using the Pichia pastoris expression system[J].Yeast, 2005, 22: 249-270.
[5] 张桂芝,武 彬,井庆川, 等.微小毛酶凝乳酶的基因克隆及在毕赤酵母GS115中的表达[J].山东农业科学,2009,11:1-4.
