
小麦新品系配合力和遗传力分析
2009-01-15 09:03于海涛闫美徐磊李安飞
山东农业科学 2009年12期
于海涛 闫 美 徐 磊 李安飞
摘 要:采用8×6不完全双列杂交,对8个小麦新品系的9个性状进行配合力和遗传力分析。结果表明,株高、穗长和穗下节间长以加性效应为主,而结实小穗数、总小穗数、穗粒数、单株穗数、单株粒重和百粒重以非加性效应为主。遗传力分析表明,株高、穗长和穗下节间长广义遗传力和狭义遗传力均较大,可在早代进行选择,其它性状早代选择效果不佳。
关键词:小麦; 品种(系); 配合力; 遗传力
中图分类号:S512.103.2 文献标识号:A 文章编号:1001-4942(2009)12-0024-05Analysis
小麦是我国的第二大粮食作物,选育新品种(系)提高小麦产量对我国农业的发展具有重大的作用[1]。配合力和遗传力是杂交育种和杂种优势利用中亲本选配的重要指标,对于评价亲本、正确选择和提高育种效率都具有重要的指导意义。前人在小麦配合力研究中已经做了大量的工作,对早熟、粒重[2,3]、沉淀值[4]、冠层温度[5]、穗部性状[6]和产量性状[7~10]等均进行了分析,为深入了解各个性状的遗传规律和指导育种工作积累了丰富的资料。本试验选用了山东农业大学农学院新选育的8个新品系进行配合力和遗传力分析,以了解其配合力和遗传力,明确其作为亲本在育种中的价值。
1 材料与方法
1.1 试验材料与设计
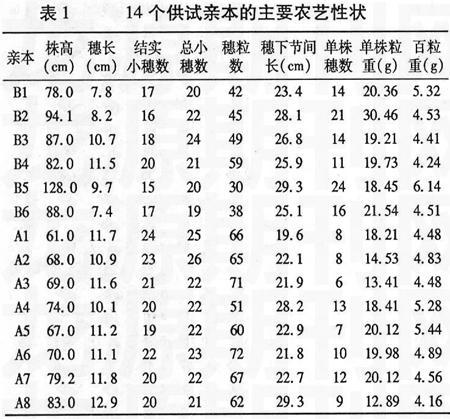
试验于2006~2007年在山东农业大学农学实验站进行。以山东农业大学选育的530001(A1)、530065(A2)、530170(A3)、530311(A4)、530319(A5)、530351(A6)、530385(A7)、530639(A8)共8个矮秆、抗病、大穗、早熟种质系为母本组,以济麦20(B1)、藁城8901(B2)、山农919(B3)、山农5536(B4)、辉县红(B5)、山农990525(B6)6个品种(系)为父本组,组配8×6不完全双列杂交。供试材料的农艺性状情况见表1。试验采用随机区组设计,重复2次,每组合F1和亲本种植2行,行长1.55 m,行距25 cm,粒距6.7 cm,人工点播。栽培管理措施按常规进行。
1.2 性状调查
于成熟前各组合随机取20个单茎,调查株高、穗长、结实小穗数、穗粒数、总小穗数、穗下节间长。完熟时每组合收获10个单株考察单株穗数、单株粒重并计算百粒重。
1.3 统计分析
采用刘来福的不完全双列杂交分析方法进行配合力模型分析[11],使用Excel和DPS软件进行方差分析[12]。
2 结果与分析
2.1 各性状配合力的方差分析
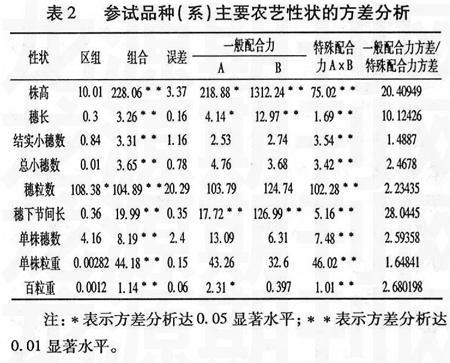
对不完全双列杂交组合的9个性状进行方差分析,结果表明:供试材料株高、穗长、结实小穗数、总小穗数、穗粒数、穗下节间长、单株穗数、单株粒重、百粒重9个性状在F1组合间存在极显著差异(表2)。因此可以对这9个性状进行配合力分析。
由表2可知,株高、穗长、穗下节间长和百粒重4个性状中除父本百粒重外其它的一般配合力差异达到极显著或显著水平,同时它们的特殊配合力差异均达到极显著水平,表明这4个农艺性状的遗传同时受基因加性和非加性效应的共同作用。结实小穗数、总小穗数、穗粒数、单株穗数和单株粒重的一般配合力差异并不显著,而特殊配合力差异达到极显著,表明这5个性状主要由非加性效应作用。由一般配合力和特殊配合力的方差比可以看出,株高、穗长、穗下节间长的一般配合力方差大幅度超过特殊配合力方差,而其余6个性状的一般配合力方差略大于特殊配合力方差。表明株高、穗长和穗下节间长3个性状的遗传方差中,加性效应占主导地位;其余6个性状的的遗传方差中加性和非加性效应同等重要。
2.2 一般配合力效应
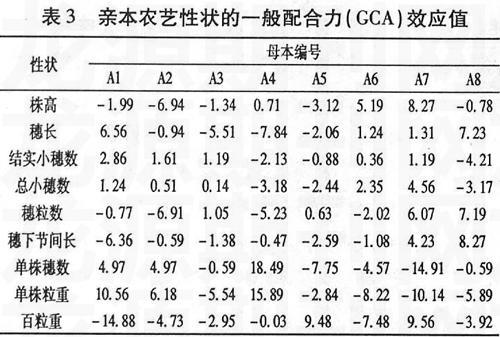
一般配合力是对基因加性效应的度量,某个亲本某个性状一般配合力效应值大,表明亲本的加性基因效应高,向后代传递能力强,易于稳定遗传和固定选取一般配合力高的亲本,可望该亲本在所配制的杂交组合中对该性状产生较大的影响。从表3中可以看出,同一亲本的不同性状或同一性状的不同亲本的一般配合力效应值差别很大。就株高而言,母本组8个亲本中A1、A2、A3、A5、A8都有降秆作用,A2的负效应值最大,降秆作用明显,利于选育矮秆高产组合。就穗长而言,A8的配合力最大,其次是A1,利于选育大穗型组合。结实小穗数的一般配合力A1最高,A8的负效应值最大。总小穗数的一般配合力A7最大。穗粒数的一般配合力A8最大,其次为A7。单株穗数和单株粒重的一般配合力效应值A4均最高,A1、A2次之。百粒重的一般配合力A7最高,A5次之。
2.3 特殊配合力效应
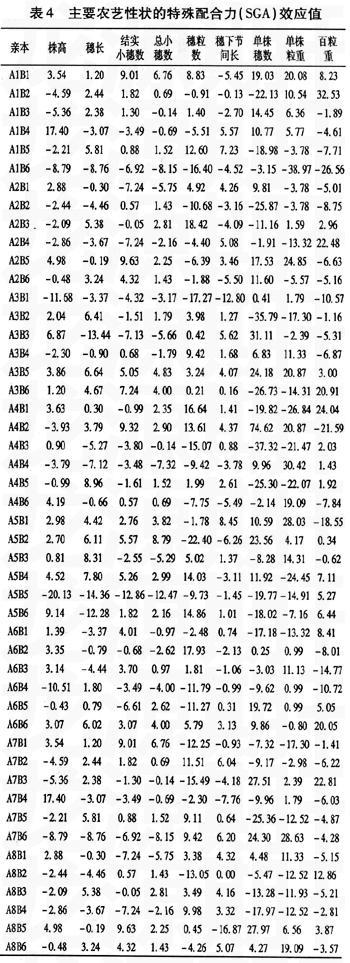
特殊配合力主要是受基因非加性效应控制,即基因的显性和上位性效应。从表4中可以看出,同一性状不同组合间差异较大,且各个性状的变异范围较大。就株高而言其变异范围为-20.13~17.40,A5B5组合株高SGA最低,降秆效果明显;穗长的变异范围-14.36~8.96,A4B5组合穗长SGA最高;结实小穗数的变异范围-12.86~9.63,A2B5和A8B5组合SGA最高;总小穗数的变异范围-12.47~8.79,A5B2组合的SGA最高;穗粒数的变异范围-22.40~18.42,A2B3组合的SGA最高;穗下节间长的变异范围-16.87~8.45,组合A5B1的SGA最高;单株穗数的变异范围-37.32~74.62,其变异范围最大,组合A4B2的SGA最高,并且该组合株高SGA为负效应,其它性状除百粒重外均有较大的正效应;单株粒重的变异范围-38.97~30.42,组合A4B4的SGA最高;百粒重的变异范围-26.56~32.53,组合A1B2的SGA最高。 从表4还可看出,A4B4组合各个产量性状的特殊配合力虽然都不是最高,但是单株粒重、单株穗数和百粒重的特殊配合力均为正效应,并且存在一定的降秆作用;该组合实际单株产量虽然不是最高,但是相对来说也较优(25.54 g)。其余组合某一性状也有很好的表现,在选配组合时可以考虑它们某一性状的特殊配合力。
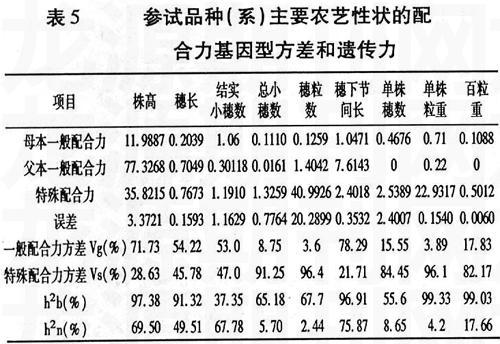
2.4 配合力基因型方差和遗传力的估计
遗传力是表示数量性状遗传过程中遗传决定与环境影响相对重要性的数量指标,是重要的遗传和育种参数。进一步分析各性状的配合力基因型方差和遗传力(表5)可以看出,株高、穗长、结实小穗数和穗下节间长的Vg(%)大于Vs(%),
说明这5个性状的加性作用相对于非加性作用更重要,而其它5个性状的Vs(%)远远大于Vg(%),说明在这5个性状中,非加性成分(显性和上位性)更为重要。广义遗传率(h2b):单株粒重>百粒重>株高>穗下节间长>穗长>穗粒数>总小穗数>单株穗数>结实小穗数;狭义遗传率(h2n):穗下节间长>株高>结实小穗数>穗长>百粒重>单株穗数>总小穗数>单株粒重>穗粒数。9个性状中有8个的广义遗传率大于50%,其中株高、穗长、穗下节间长、单株粒重和百粒重皆在90%以上,表明株高、穗长、穗下节间长、单株粒重和百粒重这5个性状受环境的影响较小。株高、穗下节间长的狭义遗传率为69.5%和75.87%,可在早期进行选择。穗长的狭义遗传率中等,宜在F4代进行选择。结实小穗数、总小穗数、穗粒数、单株穗数、单株粒重和百粒重的广义遗传率较大,但狭义遗传率较低,说明加性遗传效应较小,显性效应和上位性效应较大,应该适当提高选择的代数。
3 讨论与结论
对新育成的优良品系进行配合力和遗传力分析可以帮助我们深入了解新品系各个性状的遗传规律,同时为以后利用这些品系提供理论依据。前人在不同地区对不同的优良品系进行了大量的配合力和遗传力分析,因为所选材料的不同导致其结果存在较大差异。欧俊梅等(2000)[13]认为,株高、穗长、穗粒数和千粒重为加性和非加性效应共同作用,主要受加性基因控制。何良荣等(2001)[8]研究表明,株高、每穗小穗数以加性效应为主。庞红喜等(2000)[9]则发现株高、穗长、每穗小穗数、穗粒数和千粒重的加性、显性效应均重要。曹亚萍等(2000)[10]指出,株高受基因加性、非加性和母体效应的共同作用,以加性效应占优势;千粒重取决于加性和非加性效应,穗粒数由加性效应控制,而穗粒重存在明显的核质互作。姚金保等(2003)[3]认为千粒重符合加性—显性模型,以加性效应为主。
本试验选用山东农业大学新选育的抗病、高产新品系作为母本,通过配合力和遗传力分析评价其在育种中的应用价值,为以后的育种工作奠定基础。研究表明,在产量性状方面母本的一般配合力要大于父本即母本对于后代产量的影响要大于父本;在株高方面,A1、A2、A3、A5和A8均具有较好的降秆作用,且A1、A2、A3和A5亲本本身株高均低于70 cm,利于选育矮秆、高产品种。A1(530001)亲本的穗长GCA较大,结实小穗数的GCA最大,可以作为大穗、多小穗型亲本。A2株高的GCA最低,因此可以作为矮源亲本。A4单株穗数和单株粒重的GCA均最高,可以作为多穗、高产亲本。A5、 A6和A7其它性状的GCA表现一般,但它们的百粒重配合力很高,可以作为大粒型亲本。A8在穗粒数上GCA最大,可以作为多穗材料。
通过试验发现,株高、穗长、结实小穗数和穗下节间长这4个性状由加性效应和非加性效应共同作用,其中以加性效应为主,这和前人的研究基本一致。同时发现总小穗数、穗粒数、单株粒重、单株穗数和百粒重5个性状主要受非加性效应影响,其中总小穗数、穗粒数和单株粒重3个性状非加性效应占遗传方差的90%以上。非加性效应包括显性效应和上位性效应,其中显性效应是不能够稳定遗传到后代的,而研究表明上位性效应在遗传效应中具有重要地位并且在自花授粉作物中可以稳定遗传。如果上位性存在,将会给育种选择带来额外收益[14]。因此为了进一步明确本试验新品系的利用价值,需要研究上位性在这些性状中的作用。
参 考 文 献:
[1] 田纪春. 超级小麦的概念、育种目标和任务[J].山东农业科学,2004,5 :18 - 21.
[2] 曹亚萍,范绍强,张 娟,等.冬小麦BYDV材料粒重的配合力与遗传模型分析[J].山西农业科学,2002,30(1):17-20.
[3] 姚金保,王书文,姚国才,等.普通小麦千粒重的配合力与遗传模型分析[J].南京农学专报,2003,19(1):52-54.
[4] 张彩英,谢令琴,常文锁,等.冬小麦品种资源沉降值配合力及其它性状相关性研究[J].中国农学通报,2001,17(4):15-17.
[5] 隋新霞,李根英,吴祥云,等.不同穗型冬小麦主要农艺性状配合力分析[J].山东农业科学,2004,2:20-22.
[6] 王 瑞,宁 锟,王 怡,等.普通小麦穗部性状的配合力与遗传模型分析[J].西北农业学报,1996,5(1):1-5.
[7] 陈集贤,郜和臣,黄相国.十一个春小麦品种产量性状的配合力与遗传力分析[J].作物学报,1981,7(3):201-207.
[8] 何良荣,杜 娟,邹春梅,等.大穗小麦产量性状的配合力与遗传力分析[J].新疆农业科学,2001,38(1):16-17.
[9] 庞红喜,亢福仁,王成社,等.大穗型小麦主要农艺性状配合力分析[J].西北农业学报,2000,9(2):78-81.
[10]曹亚萍,张明仪,乔合心,等.冬小麦产量性状的配合力及遗传规律研究[J].山西农业科学,2000,28(2):7-11.
[11]刘来福编著.作物数量遗传[M].北京:农业出版社,1982,211-281.
[12]唐启义,冯明光.实用统计分析及其DPS数据处理系统[M].北京:科学出版社,2002.
[13]欧俊梅,李生容,杜小英,等.5个矮秆小麦品种(系)主要农艺性状的配合力和遗传力分析[J].绵阳经济技术高等专科学校学报,2000,17(2):1-4.
[14]张文英,程君奇,朱 军,等.上位性及其在遗传育种中的应用[J].生物信息学,2004,2(2):39-43.
[15]Cheverud J M, Routman E J. Epistasis and its contribution to genetic variance components [J]. Genetics, 1995, 139: 1455-1461.
猜你喜欢
新农业(2021年21期)2021-11-20
农民致富之友(2020年8期)2020-05-11
农民致富之友(2019年32期)2019-11-23
清明(2019年6期)2019-11-21
新农村(2017年17期)2017-08-23
科学种养(2017年5期)2017-05-17
股市动态分析(2016年6期)2016-09-29
绿色中国(2015年6期)2015-07-03
小小说月刊·下半月(2014年8期)2014-05-14
少年文艺·我爱写作文(2009年5期)2009-06-08
