
小麦抗条锈新品系89144抗锈机理研究
2006-04-29 04:11倪建福欧巧明令利军叶春雷王亚馥邢更妹李杉崔凯荣武禄光
植物保护 2006年2期
倪建福 欧巧明 令利军 叶春雷 王亚馥 邢更妹 李 杉 崔凯荣 武禄光
摘要将高粱总DNA通过花粉管通道导入小麦感病品种甘麦8号,D2代出现2株对条锈病免疫的变异株,D5代有9个株系抗性已经稳定;用混合菌和分小种鉴定,对条中29、30、洛13Ⅱ、水14、水14中梁17-s、HY3、条中31号等小种表现免疫。结果分析表明,新品系89144接种后组织中SOD活性升高,原受体甘麦8号接种锈菌后SA含量也有升高;但并不伴随有CAT活性下降,SOD活性和H2O2含量的升高;推测5A为CAT过氧化活性提供一个电子的过程中SA含量必须达到一定的阈值,并且与CAT的时序调节相配合;据SA结合态和游离态含量的变化动态,表明89144具有SA信号传导途径,推测SA信号传导途径的上游应该还有一些机制在起作用。
关键词植物病理学:小麦条锈病;抗锈新品系
中图分类号S435.121.42
小麦条锈病(Puccinia striiformis f.sp.triti-ci)是甘肃省小麦主要病害之一,严重影响小麦高产稳产。国内外研究和生产实践证明,选育抗锈病品种是防治该病最经济有效的途径。为了培育新的抗锈品种,从1988年起,在总结以往工作的基础上开展了应用花粉管通道法将外源C4作物高粱DNA向小麦转移的探索,1989年在大田和温室共做了80个外源DNA导人组合,获得抗条锈病新品系89144,并对其后代材料作了较系统的观察和研究。本文报道小麦抗条锈新品系主要特征和抗性生理生化变化进行比较测定和分析。
1材料和方法
1.1材料
受体为普通小麦(Triticum sestrvum),品种甘麦8号,株高107cm左右,穗型棍棒状、顶芒、生育期104d,适应性强,曾大面积种植,现重感条锈病;供体为高粱[Sorghumbicolor(L.)]品种米高粱,抗旱、耐涝、耐盐碱、适应性强。
1.2方法
1.2.1DNA提取和导入
供体高梁DNA提取和导人参照倪建福等方法。
1.2.2田间资料统计与观察
经处理的当代受体记作DO,其后代依次记为D1、D2和D3等。DO~D1按组合收种子,以后按系谱法选择,等抗性稳定后再按组合收种子,种植按行长1.20m、行距0.20m、粒距0.04 m点播,田间观察和室内外考种项目按常规育种要求进行,各种材料按种植小区和株系收获。
1.2.3抗锈性鉴定
除本中心每年用心叶涂抹法接种流行混合菌选择外,对获得的抗锈新品系再送甘肃省农业科学院植保所进行专门鉴定,反应型按常规标准汇载。
1.2.4种子粗蛋白和淀粉含量的测定
抗锈新品系89144和供、受体一并送测试中心,按照国家规定的标准方法测定。粗蛋门是将材料先采用凯氏法处理后,用瑞典Tecalor公司的Kielteasystem Ⅱ氮/蛋白质分析仪测定;赖氨酸采用国际染料结合法(PBL法)染色,再用北京环保仪器厂的GXDL。-202型蛋白质赖氨酸分析仪测定;淀粉采用国际旋光法测定”。
1.2.5水杨酸(psalicylicacid,SA)含量的测定
将小麦种子浸种后,播种于塑料钵蛭石内,在小麦一叶展叶时利用心叶涂抹法接人小麦条锈菌混合菌种,培养温度13~19℃.光照12h/d,湿度100%,接种后24、32、40、48、56 h取材。参考李兆亮等方法用硅胶板纯化粗提液,用HPLC(仪器型号WatersTM600EC-18柱)检测样品中SA含量,检测温度40℃,流速为0.6 mL/min,检测波长310 nm,保留时间4.05 min。
1.2.6过氧化氢酶(CAT)活性的测定
取0.5g叶片加1.8mL预冷的提取缓冲液(10 mmoI/L Tris-HCl pH7.5,0.25 mol/L蔗糖,1 mmol/L EDTA,10.5 mmol/L DTT,0.1 mmol/LPMSF),研磨成匀浆后在4℃条件下27000g离心15min,上清液按崔凯荣方法测定酶活性。
1.2.7过氧化氢(H2O2)含量的测定
取1g叶片参考Patterson等方法加3mL冷丙酮磨成匀浆,16000g离心10 min,上清液定容,反应液中含0.1mL体积分数20%TiCL4的浓盐酸,0.2 mL浓氨水和1mL。上清液。生成的过氧化物-Ti复合物用丙酮洗5次.丙酮挥发后溶于3mL。硫酸(1mol/L)中,测410nm光吸收值,制作H2O2标准曲线。
1.2.8超氧化物岐化酶(SOD)活性的测定
取0.5g叶片加1,8 mI。预冷的提取缓冲液(50mmol/LpH7.8磷酸缓冲液,0.1 mmol/LED-TA,体积质量0.3%TritonX-100;体积质量4%聚乙烯吡咯烷酮(PVPP)研磨后用纱布过滤,10500g离心20 min,上清液为粗酶液。按照EL-moshaty等方法测定SOD活性,根据SOD抑制NBT光化学还原的量计算酶活性。
1.2.9过氧化物酶(POD)活性的测定
取0.5g叶片参考崔凯荣等方法加1.8 mL预冷的提取缓冲液(100mmol/LPBS,pH6.0.内含体积分数0.1%TritonX-100和体积质量0.04%偏重亚硫酸钠。冰浴研磨,14 000 S离心15 mln。以愈创木酚作底物,在470 nm处测OD值。以△OD/min·mg蛋山为酶活单位。
2结果与分析
2.1生物学特性
1989年将高梁DNA通过花粉管通道导入小麦感病品种麦8号,共处理小花或子房134个,获得Do代种子28粒,结实率为20.90%。1990年点播成苗27株;1991年种植900株,从中选出2株变异株;1992年从中选1株种子点播;1993年种植l800株.获得变异株121株;1994年种植1140株,入选41株;1995年种植1800株,从中选出稳定遗传的抗锈新品系89144系列,其生物学特性,见表1。
两个抗锈新品系的大多数生物学性状是介于受体和供休之间,如叶功能期延长,株高降低,穗长和粗蛋白增加以及千粒重、赖氨酸和淀粉含量等均介于供体和受体之间,但籽粒饱满度增加,小穗排列疏松,叶片变窄变长,颖壳变硬(表2)。

抗锈性鉴定结果表明,两个抗锈新品系89144
对目前流行的所有小种免疫,特别是在近年甘肃生产上应用的所有小麦品系几乎对条中3l和32号丧失抗性的情况下,89144仍然抗锈,与其杂交的组合后代也非常抗锈,见表2。
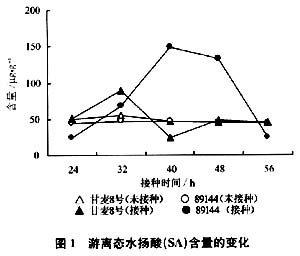
2.2水杨酸含量变化
在接种条锈菌后,通过HPLC测定结合态与游离态SA含量变化结果如图l、2。抗锈新品系89144游离态水杨酸从32 h开始逐渐升高,40 h达到峰值,然后丌绐缓慢下降,到48h急剧下降。甘麦8号从24h开始缓慢上升,上升幅度比89144小2倍,到32 h快速下降,而在未接种锈菌的对照中,89144和甘麦8号内源游离SA含量变化都不大。图2表明,接种后89144、甘麦8号的结合态SA含量都有上升,并且甘麦8号比89144上升幅度大而且时间早。在对照中,89144、甘麦8号的结合态SA含量变化不大。
2.3过氧化氢酶活性变化
由图3可以看出,抗锈新品系89144的CAT活性在接种条锈菌40h处开始大幅度下降,48h后又略有回升,而甘麦8号变化不大。对照未接种条锈菌的89144、甘麦8号,CAT活性变化不大。

2.4过氧化氢含量变化
图4表明,在接种锈菌48 h后,抗锈新品系89144中H2O2含量达到峰值,甘麦8号含量也略有升高,而对照89144、甘麦8号H2O2含量变化不大。
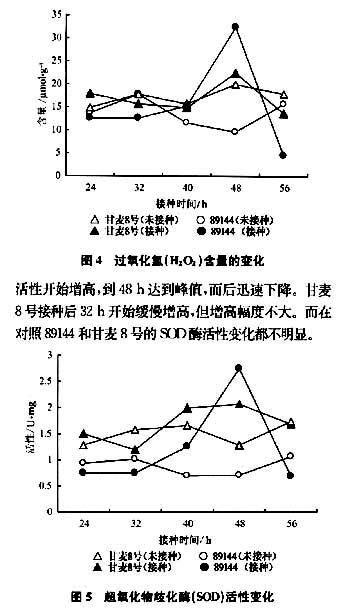
2.5超氧化物歧化酶活性变化
图5表明,抗锈新品系89144在接种后40h,SOD活性开始增高,到48h达到峰值,而后迅速下降。甘麦8号接种后32h开始缓慢增高,但增高幅度不大。而在对照89144和甘麦8号的SOD酶活性变化都不明显。
2.6过氧化物酶活性变化
图6中抗锈新品系POD酶活性无论在接种条件下还是在未接种条件下变化都不大,只是在接种56 h后抗锈新品系89144的POD活性稍有升高。

3讨论
经多年实践表明,利用花粉管通道将外源DNA导入小麦胚囊,转化上不具备正常细胞壁即合子或早期细胞是完全可行的,因为DNA片断与受体基因组部分基因间有可能存在同源性而发生杂交重组。该技术之所以提高选育出抗锈品系的成功率,是因为供体一方的染色体经提取后片断变小,从而减少了整条染色体之间的排斥性,增加了部分染色体重组几率。
实验研究显示,外源DNA导入后代抗锈新品系89144在接种锈菌40h后,内源游离SA含量达到峰值,从40 h至48 h有一个缓慢下降阶段,同时伴随着CAT活性降低,H2O2含量增加,SOD活性增高。而不抗锈品种甘麦8号在接种锈菌后,也有内源游离SA含量升高,但并不伴随有CAT活性下降和SOD、POD活性升高,也没有H2O2含量的上升。Chen等曾研究证明烟草中的SA结合蛋白(SA—bindingprotem,SABP)具有CAT活性,可以降解H2O2产生O2和H2O,SA阻遏SABP的CAT活性,SA作为电子供体为CAT的过氧化活性提供一个电子,同时抑制并使这些酶处于不活跃状态,从而导致H2O2含量升高。从本实验结果可以看出虽然抗锈新品系89144和甘麦8号接种锈菌后,SA含量都有升高,但甘麦8号H2O2含量达到峰值的时间早,峰值低,这可能是因为SA为CAT过氧化活性提供一个电子的过程中SA的量必须达到一定的阈值,并且与CAT的时序调节相配合。
实验中还发现,甘麦8号、抗锈新品系89144接种锈菌后游离SA含量增高,结合态SA含量也升高,所以在诱导条件下游离SA含量的升高不一定来自于结合态SA的释放,还可能是植物体内与SA合成的相关基因被启动。这样,在SA信号传导途径的上游就还有某些机制在起作用。抗锈新品系89144接种锈菌后SOD活性也有升高,SOD可使O2通过歧化反应产生H2O2,在正常条件下,植物体中的SOD活性足以清除体内的O2,使O2保持在正常生理水平。但在逆境条件下,O2水平的提高刺激了SOD活性的增加,这可能与病原物诱导的氧化喷发有关。但在实验中并没有发现POD活性的大幅度变化,也许在检测时POD活性已恢复常态,这也符合氧化喷发的时相。由此推测在植物体内H2O2含量升高可以通过多种途径造成,Chen等的实验也证明H2O2或它的衍生物活性氧可激活抗病反应途径中相关基因表达。
