
苎麻疫霉抗甲霜灵突变株对棉苗的致病力及其遗传
2006-04-29 00:44陈方新齐永霞高智谋吴红星汪爱娥
植物保护 2006年2期
陈方新 齐永霞 高智谋 吴红星 汪爱娥
摘要研究了苎麻疫霉抗甲霜灵(Mtr)突变株对棉苗的致病力及其遗传。结果显示,抗性突变株对棉苗的致病力与其野生型亲本间无显著差异。而Mtr突变株XC-6-2对棉苗的致病力在其无性第1代(ZG1)单孢株间及其与亲本问均存在极显著差异,且XC-6-2对棉苗的致病力性状在单游动孢子第2代(ZG2)持续发生分离;XC-6-2单卵孢第1代(OG1)对棉苗的致病力间亦存在极显著差异。上述结果提示,抗甲霜灵突变株对棉苗的致病力在无性单孢和单卵孢子后代间均不能稳定遗传。对比研究发现,野生型亲本菌株XC-6对棉苗的致病力在单游动孢子卸单卵孢后代有相似的遗传规律。且单游动孢子群体对棉苗致病力的变异系数较其单卵孢株间的变异系数大。
关键词植物病理学;苎麻疫霉;抗甲霜灵突变株
中图分类号S435.621.24
甲霜灵属特异性位点抑制剂,病菌容易对其产生抗药性突变。关于甲霜灵处理可导致疫霉菌致病力发生改变已有一些报道。研究指出致病疫霉(Phytophthora infestans)抗甲霜灵(Mtr)突变株的致病力强于对甲霜灵敏感(Mts菌株,而Dowley报道,田间获得的致病疫霉(P.infestans)Mtr菌株的孢子萌发率和致病力与Mts菌株相似。Stack等亦报道,大雄疫霉大豆专化型(P.megasperma f.sp.sojae)的Mtr与Mts菌株的致病力基本相似。而关于苎麻疫霉抗甲霜灵突变株对棉苗致病力的遗传与变异未见报道。
本文以苎麻疫霉为材料,研究抗甲霜灵突变株对棉苗致病力在无性和有性后代的遗传,旨在明确苎麻疫霉抗甲霜灵突变株对棉苗的致病力及其遗传背景。
1材料与方法
1.1供试菌株
供试菌株XC-6、WH-2、wJ-5、SX-9、WW-13为苎麻疫霉(Phytophthoraboehmeriae Sawada)野生型敏感菌株‘,XL'6—2、XC 6—3、WH—2—2、WJ—5—2、SX 9—2、SX~9-4、WW-13—1、WW-13—3均为上述野生型菌株经甲霜灵盲接诱变筛选获得的杭甲霜灵(Mt)突变株。
1.2游动孢子囊的诱导和单游动孢子无性系的建立参照高智谋等的方法。
1.3单卵孢株群体的建立参照高智谋等的方法。
1.4对棉苗致病力的测定
1.4.1供试棉苗
供试棉苗品种为泗棉3号,由安徽农业大学农场提供。棉籽经浓硫酸脱绒后,在55~60℃温汤浸种20 min,30℃下催芽24 h.露白后播种于温室铁皮盘内的表层土壤(稻田土)中,培育10 d左右待子n十充分展开后,拔取大小相当的健康棉苗插于装有灭菌水的小试管(15 mmXl50 mm)中,供接种用。
1.4.2接种方法
供试菌株在IBA平板上,25℃、黑暗条件下培养3 d后,用打孔器((d=5mm)沿菌落边缘取菌碟。在棉苗叶片中央用6根小号昆虫针束刺l下,将打好的菌碟置于创伤处,用脱脂棉蘸灭菌水覆盖保湿,置25℃恒温光照下培育。24h后去掉棉球和菌丝块,48h后测量病斑直径。每菌株接种2片子叶,设3次重复。以6片子叶的平均病斑直径大小评价菌株致病力的强弱,以空白培养基块接种作为对照。
2结果与分析
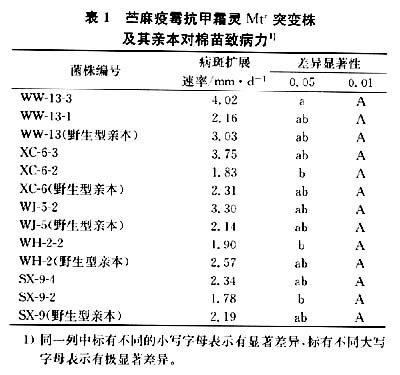
2.1抗甲霜灵(Mtr)突变株对棉苗的致病力
测定结果(表1)表明,Mtr突变株XC—6—2、XC—6—3、WJ—5—2、WW 13—1、WW—1 3—3、SX—9—2、SX—9—4对棉苗的致病力与其各自的野生型亲本菌株对棉苗的致病力间无显著差异,苎麻疫霉菌株抗中霜灵突变对其致病力性状的表达影响不显著。
2.2抗甲霜灵突变株对棉苗致病力的遗传
2.2.1突变株单游动孢子后代对棉苗的致病力
抗性测定结果表明.突变株XC-6—2的Mtr性状在无性及有性单孢后代均能稳定遗传。以Mtr突变株XC—6—2为亲本建立单游动孢子第1代(ZG,),并测定其2G1代单孢株对棉苗子叶的致病力。表2、图1显示,Mtr突变株XC—6—2ZG:代单孢株在棉苗子叶上的平均病斑扩展速率分布范围分别为1.75—5.88 mm/d,变异系数为35.92%。经方差分析(表2)显示,Mtr突变株XC—6—2对棉苗的致病力在其2G1代群体间及其与亲本间存在极显著差异(F=2.97,F0.01=1.88)。
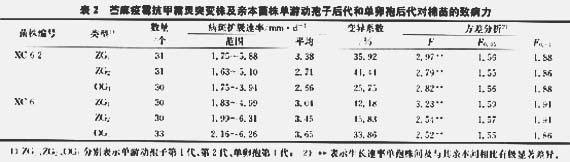
随机选取XC-6—2ZG,代单孢株为亲本,建立单游动孢子第2代(ZG2)群体,并进一步测定了XC—6—2 ZG2代单孢株对棉苗的致病力。测定结果(表2、图1)为,31个单游动孢子株在棉苗子叶上平均病斑扩展速率分布范围为1.63—5.10 mm/d,平均为2.71 mm/d,表明突变株对棉苗的致病力在其单游动孢子第2代(ZG2)仍持续发生分离,且甲孢第2代群体对棉苗致病力的变异系数(CV=41.44%)较其第1代单孢株的变异系数(CV=35.92%)高。方差分析表明,苎麻疫霉Mtr突变株XC-6—2对棉苗的致病力在其ZG2代单孢株间及与亲本间相比均有极显著差异(F=2,79,F0.01=1.86)。

2.2.2突变株单卵孢后代对棉苗的致病力
以Mtr突变株X0 6—2ZG:代为亲本建立单卵孢第一代(OGl)群体,测定了其单卵孢株对棉苗的致病力,结果(表2,图1)显示,OG,代单孢株所致病斑扩展速率在1.75—3.94 mm/d,平均为2.66 mm/d,变异系数(CV=25.75%)明显小于其单游动孢子后代对棉苗致病力的变异系数。突变株XC-6-2单卵孢第1代(OG)对棉苗致病力具有极显著差异,提示苎麻疫霉突变株对棉苗的致病力在其有性单卵孢后代发生较大变异,即苎麻疫霉Mtr突变株对棉苗的致病力在其单卵孢后代亦不能稳定遗传。
采用同样的方法测定了野生型亲本菌株XC-6和WH-2单游动孢子和单卵孢后代对棉苗的致病力,得到类似结果(表2,图1)。
综上所述,苎麻疫霉Mtr突变株对棉苗的致病力在/乙:和Z乙2代持续发生分离,且XC 6-2对棉苗致病力性状在单卵孢后代亦不能稳定遗传。同时,测定结果还显示,该突变株的致病力性状在单游动孢子后代及单卵孢后代的遗传变异特点与其野生型亲本基本一致。
3小结与讨论
疫霉菌致病力的遗传与变异以及控制该性状的遗传背景十分复杂。一般认为,疫霉菌的致病力在无性繁殖过程中可以较稳定地保持,而在有性生殖后代发生复杂的分离或变异。但有报道指出,一些疫霉菌的致病力在单游动孢子后代发生变异,甚至在人工培养基上经长期培养后,出现致病力衰退或丧失的现象。有关疫霉菌致病力发生变异的遗传机制,Gallegly研究认为致病力的遗传与变异可能由细胞核因子控制,而Caten认为该性状与细胞质中的线粒体基因有关;另外,有报道认为异核现象、细胞核杂合性均可能是引起疫霉菌致病力发生变异的重要因子。
关于甲霜灵对疫霉菌生物学性状的影响,报道认为从田间分离或室内诱变获得的致病疫霉(p.infes—tans)抗甲霜灵(Mtr)菌株,除抗药性性状发生变异外,其他生物学性状如致病力等亦可能发生改变。作者研究了苎麻疫霉抗甲霜灵突变株对棉苗的致病力及其在单孢后代的遗传。方差分析结果显示,苎麻疫霉抗甲霜灵菌株对棉苗致病力与其亲本无显著差异,提示病菌对甲霜灵的抗性突变并未改变其对棉苗的致病力。抗甲霜灵突变株无性及有性单孢后代对棉苗致病力的遗传分析显示,Mtr突变株与其野生型亲本菌株对棉苗的致病力在其2c1和zG2代发生持续分离或变异,同时其致病力在单卵孢第1代(G1)群体间亦有极显著差异,但单卵孢后代群体对棉苗致病力间的变异系数明显低于单游动孢子群体间的变异系数。上述对比研究结果表明苎麻疫霉Mtr突变株对棉苗的致病力性状与其野生型亲本间有相似的遗传变异特点。因此,该抗性突变株在田间一旦产生,将与野生型菌株间有相同的竞争能力,并且当遭遇苯基酰胺类(如甲霜灵)高选择压的情况下,该抗性个体被保留并可能不断扩散和蔓延而导致抗性群体的出现和该类杀菌剂的失效。至于控制该性状的基因有待进一步通过分子手段加以研究阐明。
