
丹江流域浮游植物群落的时空变化及驱动因子
2025-03-04 00:00:00李明叶思思李娟薛旭东胡恩
河南师范大学学报(自然科学版) 2025年2期



摘 要:浮游植物是水域生态系统中的重要指示物种,其群落的组成和分布与环境因子密切相关.丹江是丹江口水库的主要水源,研究丹江流域浮游植物的时空变化特征以及驱动因子,对丹江流域水资源保护十分重要.研究采集了2个时期(枯水期和丰水期)丹江干流和6条支流的水样及浮游植物样品,通过对浮游植物的鉴定及分析,明确不同时间和空间浮游植物群落结构对环境因子的响应.研究鉴定出丹江流域浮游植物7门62属104种,主要以硅藻门(Bacillariophyta)为主.枯水期和丰水期丹江流域优势藻均为曲壳藻(Achnanthes).前20优势藻中11种为硅藻门.丹江流域水温(WT)与多样性指数显著相关,且在枯水期更显著.丹江流域枯水期浮游植物群落结构变化的主要影响因子为总氮(TN),丰水期浮游植物群落结构变化的主要影响因子为WT、pH、溶解氧(DO)和TN.丹江流域浮游植物群落主要受到WT、TN和总磷(TP)环境因子的调控.丰水期网络的节点、边和模块化较枯水期更高且浮游植物之间主要呈现正相关系.上游网络分为4个模块,而下游仅分为2个模块且浮游植物之间主要呈负相关关系.研究为丹江流域水生态的保护提供了理论依据.
关键词:丹江流域;浮游植物;环境因子;多样性指数;时空变化
中图分类号:Q948.8""""" 文献标志码:A文章编号:1000-2367(2025)02-0001-13
收稿日期:2024-06-14;修回日期:2024-08-31.
基金项目:国家自然科学基金(51979236).
作者简介:李明(1986-),男,甘肃庆阳人,西北农林科技大学教授,博士,研究方向为水生态保护,E-mail: lileaf@nwsuaf.edu.cn.
通信作者:胡恩,E-mail: huen777@163.com.
引用本文:李明,叶思思,李娟,等.丹江流域浮游植物群落的时空变化及驱动因子[J].河南师范大学学报(自然科学版),2025,53(2):1-13.(Li Ming,Ye Sisi,Li Juan,et al.Temporal and spatial changes and driving factors of phytoplankton communities in the Danjiang River Basin[J].Journal of Henan Normal University(Natural Science Edition),2025,53(2):1-13.DOI:10.16366/j.cnki.1000-2367.2024.06.14.0002.)
浮游植物是水域生态系统中重要的初级生产者[1],在水域生态系统的能量流动、物质循环和信息传递中起着至关重要的作用[2].浮游植物群落的组成及其多样性的变化直接或间接地影响着其他水生生物的丰度和分布,对水域生态系统的稳定至关重要[3].浮游植物群落的组成及分布受到季节性演替和水环境空间变化的影响[4].在水环境中浮游植物群落受到生物和非生物因子的影响[5].研究表明,非生物因子如水温、光照、营养盐等和生物因子如滤食性鱼类、底栖动物、水生植物、浮游动物等影响浮游植物群落的组成和分布[6].因此,明确浮游植物群落的组成分布、多样性变化以及对环境因子的响应,对水域生态系统的保护与管理十分必要.
河流是地球生态系统的重要组成部分,具有调节气候、涵养水源、保持生物多样性等多种生态功能[7].然而,由于人类活动导致的河流生态系统受损已成为全球关注的热点问题之一[8].科学地评价河流健康状况是进行水生态系统修复及保护的前提[9].浮游植物对水环境变化响应快速,对水质的指示作用灵敏,常被用作河流健康的重要指示物种[10].国内外学者从浮游植物群落结构对环境因子的响应[10-11]、浮游植物多样性对水质的评价[9-10]、浮游植物的季节演替[12-13]等方面进行了大量的研究.然而传统的多样性指数虽然能对水质情况进行评价,但是不能很好地描述浮游植物的生长状况,YE等[14]根据浮游植物丰度提出的多样性指数主要用来描述浮游植物的生长状况,提出的a、k和N分别用来描述物种濒危临界占比、竞争强度以及环境容纳量.由于丹江流域对a、k和N指数的研究尚存在空白,因此亟须开展针对该区域的本地化应用研究.
丹江是汉江的主要支流之一,同时也是丹江口水库的主要水源,丹江流域水环境健康对汉江及其下游地区用水安全十分重要.了解丹江流域浮游植物群落的组成、分布、多样性变化以及对环境因子的响应,可为水域生态系统的保护提供重要的参考.本研究通过实地调查,采集2个时期(枯水期和丰水期)丹江干流和6条支流的水样及浮游植物样品,测定丹江流域水体的环境因子并对浮游植物进行鉴定和计数,拟回答以下问题:(1)丹江流域枯水期和丰水期浮游植群落的变化是否存在差异?(2)丹江流域支流和干流,上游和下游浮游植物群落的变化特征如何?(3)影响丹江流域浮游植物群落的主要环境因子是什么?是如何影响的?通过上述研究,明确丹江流域浮游植物的时空变化特征以及重要的驱动因子,为保护丹江流域水资源提供理论依据.
1 材料方法
1.1 研究区域概述
丹江是汉江最大的支流,发源于秦岭南麓凤凰山,流经陕西、河南和湖北3省,全长443 km,总流域面积16 812 km2,在陕西境内全长249.6 km,流域面积7 510 km2,约占总流域面积的44.7%[15].选取丹江流域为研究区域,包括丹江干流,6条支流分别是麻街河、南秦河、银花河、武关河、清油河和滔河.在干流布设了4个采样点,每条支流按照上游、中游和下游分别布设3个采样点,共计布设了22个采样点,如图1所示.
1.2 水样的采集及测定
于2023年3月和8月开展丹江流域实地调查与样品采集.使用便携式多功能参数仪(Mettler Toledo AG,Switzerland)在采样现场测定水样pH和溶解性固体(TDS),测定前在野外环境下使用标准缓冲液对仪器进行校准,并在每个采样点检查仪器.使用有机玻璃采水器,在水面以下0.5 m处采集1 L水样,装于聚乙烯塑料瓶中,聚乙烯瓶使用前已预先用体积分数5%的HNO3浸泡24 h,并用Milli-Q水冲洗干净.采集完成的样品在野外使用装有冰块的保温箱低温避光保存,并迅速运送到实验室测定总氮(TN)和总磷(TP).TN和TP直接用原水样测定,测定方法参考文献[16].
1.3 浮游植物的采集及鉴定
使用采水器采集水面以下0.5 m的水样,装于1 L的聚乙烯塑料瓶中,并向每升水样中加入15 mL左右的鲁哥试剂进行固定,避免因时间过长而导致样品变质.将固定后的样品沉淀48 h,用虹吸管小心吸出上面不含藻类的“清液”,剩下20~25 mL沉淀物转移至30 mL定量瓶中,再用上述清液洗涤3次沉淀,将洗涤液转入上述定量瓶中,再补加0.5 mL鲁哥试剂后定容至30 mL.
将样品混匀后,立即吸取0.1 mL置于20 mm×20 mm浮游生物计数框中并盖上盖玻片,使用奥林巴斯显微镜(CX31,Olympus,Tokyo,Japan)进行观察,并依据文献[17]对藻类进行鉴定.在10×20倍镜下或者10×40倍镜下进行计数.计数原则为:当藻类密度较大时,单一藻类的个体计数200个即可.如果藻类密度过小,则需全片计数.每瓶藻样取3次计数的平均数.1 L样品中浮游藻类的个体数U可按下式计算:
U=A×VsAc×Va×n,(1)
式中,U表示每升原水样中的浮游藻类数量(L-1);A表示计数框面积(mm2);Ac表示计数面积(mm2);Vs表示1 L原水样沉淀浓缩后的体积(mL);Va表示计数框体积(mL);n表示计数所得浮游藻类的数目.
1.4 数据分析
根据YE等[14]提出的多样性指数,根据式(2)和(3)分别计算濒危物种丰度占比a、竞争系数k和环境容纳量N.濒危物种丰度占比a反映了在生态系统中濒危物种相对于其他物种的丰度.a值越大,说明濒危物种在样本中的丰度占比越高.这对于识别生态系统中可能面临威胁的物种具有重要意义,有助于制定保护策略.竞争系数k是衡量物种之间竞争强度的一个参数.k的值决定了相对丰度随物种排序的变化速率.当k值较大时,表示物种之间的竞争较为激烈,物种丰度的差异也较大;相反,当k值较小时,物种之间的丰度分布更为均匀,竞争相对较弱.这一指标对于分析生态系统内部的物种间互动具有重要作用,能够帮助理解在不同环境压力下物种如何适应和共存.环境容纳量N表示一个特定环境中所能容纳的物种数量.N的大小受环境因素的影响,包括资源可用性、空间限制和环境稳定性.较大的N值表明该环境能够容纳较多的物种,显示出较高的生物多样性,而较小的N值可能意味着环境较为贫瘠或受到较大的生态压力.
对于每一个样品,以相对物种丰度的排序i为自变量,以Pi为因变量,使用指数模型进行拟合:
i=a×eki,(2)
式中,为物种i的相对丰度的计算值,e为自然常数,i为物种排序数.
每一个样品中,所有的物种的相对丰度/生物量占比总和为100%.使用等比数列前n项和,得到:
Sn=a1(1-ekn)1-ek=100%,(3)
式中,Sn为等比数列前n项和,a1=0×ek,N为环境物种数容纳量.通过式(3)可解出N.
常用的α多样性指数包括香农指数(Shannon index)[18]、辛普森指数(Simpson index)[19]和皮卢指数(Pielou index)[20],优势度(Y)用来确定浮游植物的优势种[21],我们根据浮游植物的丰度分别进行计算.
利用CANOCO 5.0软件分析环境因素与常见藻类分类的关系.先用species-sample数据做去趋势对应分析(DCA),因为分析结果中Lengths of gradient的第一轴小于3.0,所以选择RDA分析来反映浮游藻类群落与环境因子之间的关系.利用R语言中的“SparCC”包计算藻类相对丰度之间的两两相关关系(皮尔逊相关性).网络可视化通过Gephi version 0.10.1版本绘制.每个节点表示每种藻的度,每条边代表2个藻类之间的相关性.利用Excel 2019对数据进行分析,Origin 2022软件进行作图和描述性统计分析.利用Arcgis 10.1进行地图的绘制.
2 结 果
2.1 水质因子的时空变化
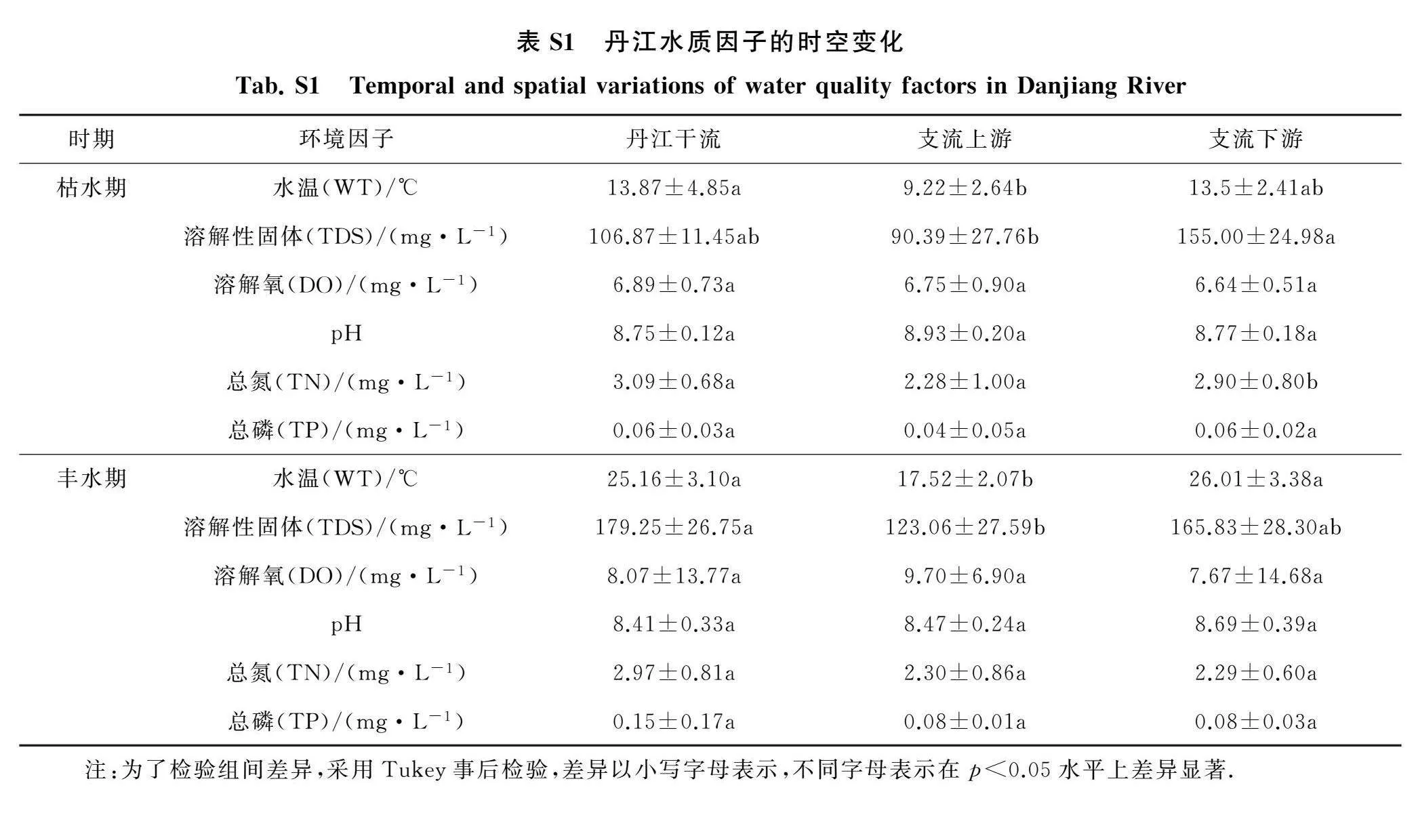
丹江水质因子的时空变化见附录表S1所示.不同时期支流上游的温度均要低于支流下游和丹江干流.枯水期支流上游的TDS要显著低于下游,丹江干流和支流上游的TN质量浓度要显著高于下游.枯水期丹江干流、支流上下游的DO、pH和TP的变化差异不显著.丰水期丹江干流的TDS要显著高于支流上游.丰水期丹江干流、支流上下游的DO、pH、TN和TP的变化差异不显著.
2.2 浮游植物的组成和分布
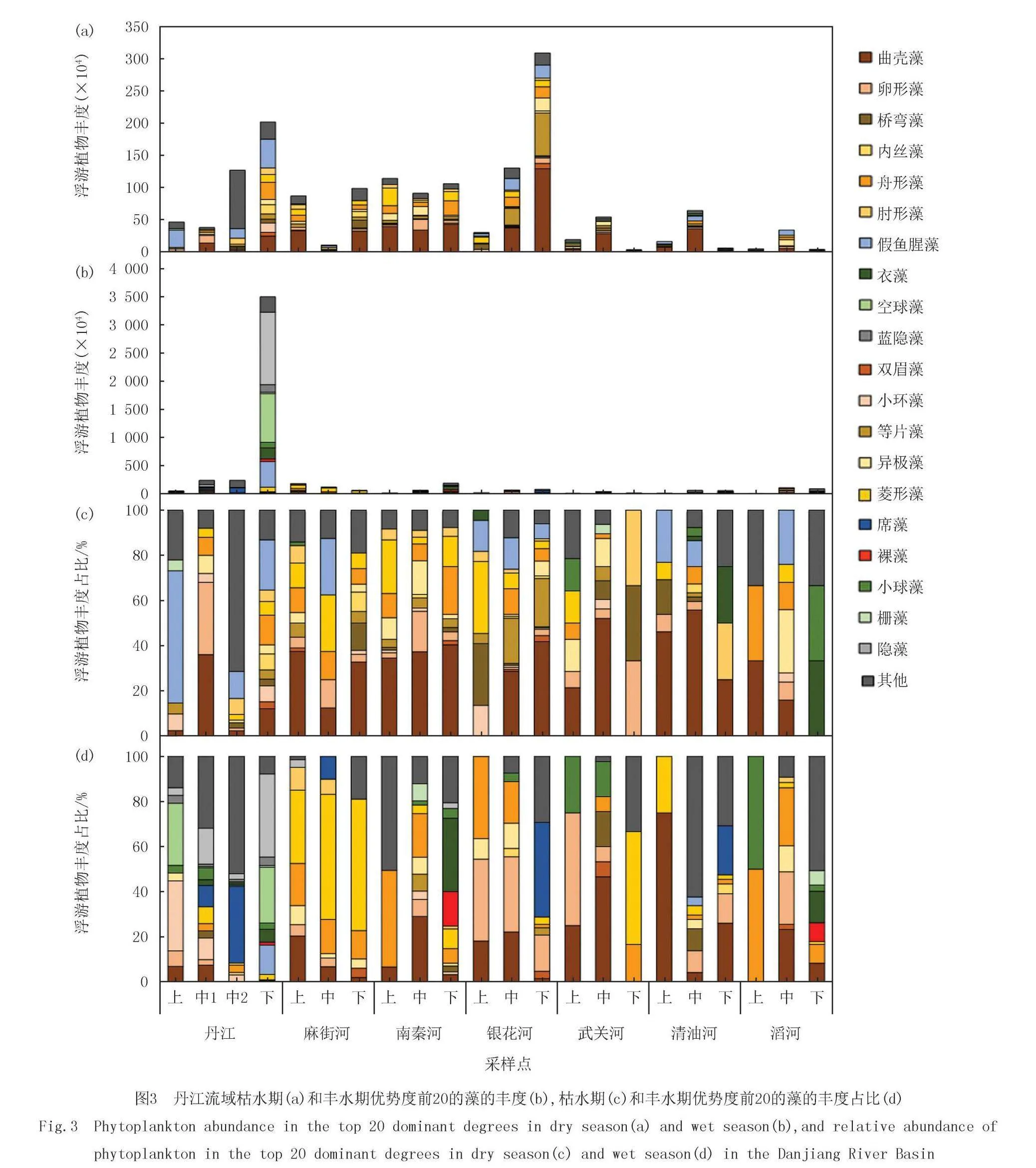
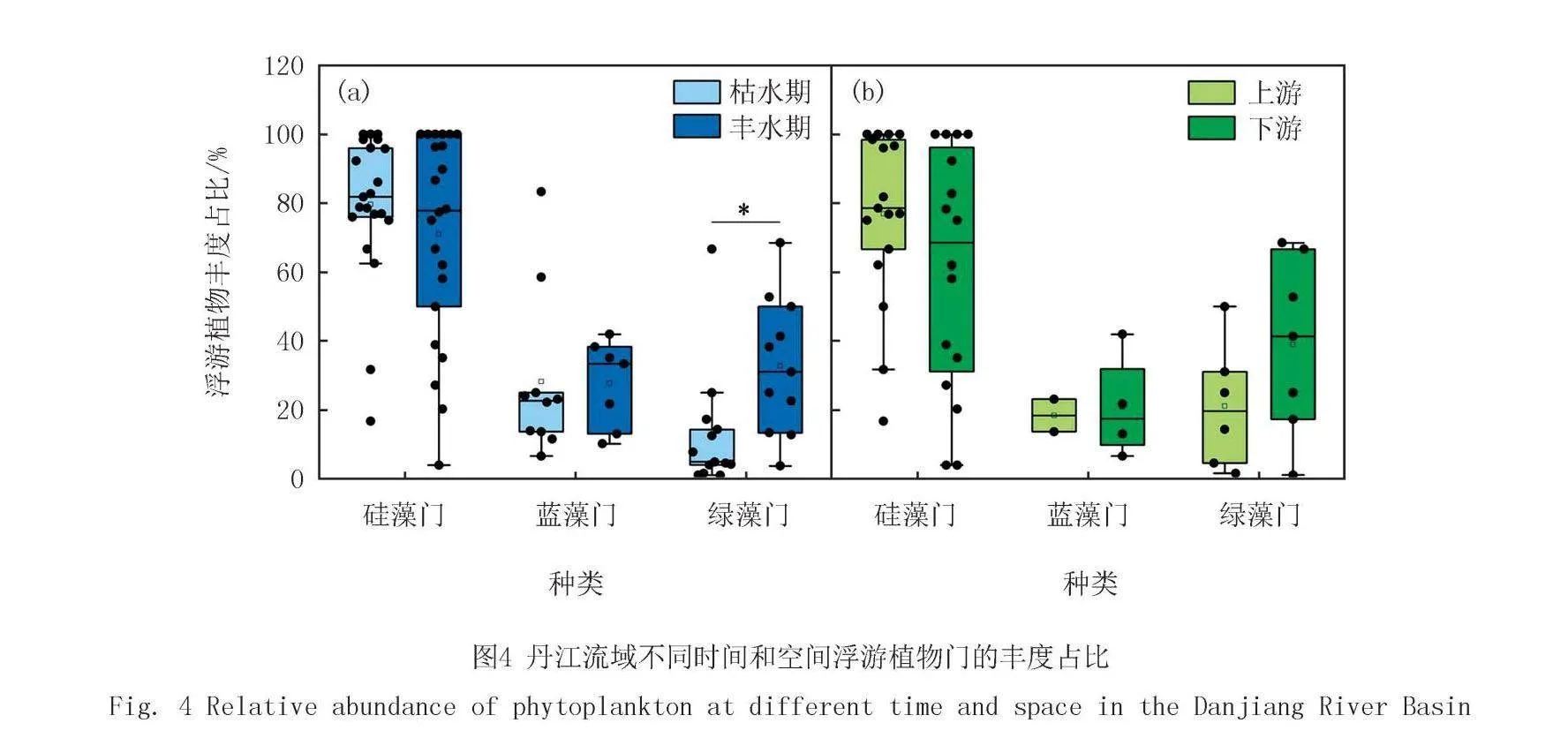
丹江流域共鉴定出浮游植物7门62属104种.丹江流域河流主要以硅藻门(Bacillariophyta)为主(图2).枯水期干流主要为硅藻门,其次是蓝藻门(Cyanobacteria),丰水期干流上游到下游群落组成从已硅藻门占优逐渐变成以隐藻门(Cryptophyta)和绿藻门(Cyanophyta)占优.6条支流上游均以硅藻门占优.枯水期滔河下游以绿藻门占优,其次是金藻门(Chrysophyta).丰水期南秦河下游主要以绿藻门占优其次是硅藻门,滔河下游以绿藻门和硅藻门占优.
优势度前20的浮游植物丰度占比如图3所示.前20优势藻中其中11种为硅藻门,4种为绿藻门,2种为蓝藻门,2种为隐藻门和1种为裸藻门(Euglenophyta).丹江流域优势藻均为曲壳藻(Achnanthes).枯水期干流从上游到下游假鱼腥藻(Pseudanabaena)减少,曲壳藻增多,丰水期干流从上游到下游小环藻(Cyclotella)减少,隐藻(Cryptomonas)增加.
丹江流域前3种优势门分别是硅藻门、蓝藻门和绿藻门(图4).枯水期和丰水期硅藻门的占比分别是80%和71%.蓝藻门枯水期除了在干流有2个样点的占比超过60%外,其余样点的蓝藻门丰度占比均低于30%,丰水期蓝藻门丰度占比不存在异常值,均值为28%.枯水期和丰水期绿藻门丰度占比分别是13%和33%.支流上游硅藻门占比高于下游,下游蓝藻门和绿藻门的占比高于上游.
2.3 浮游植物的多样性指数
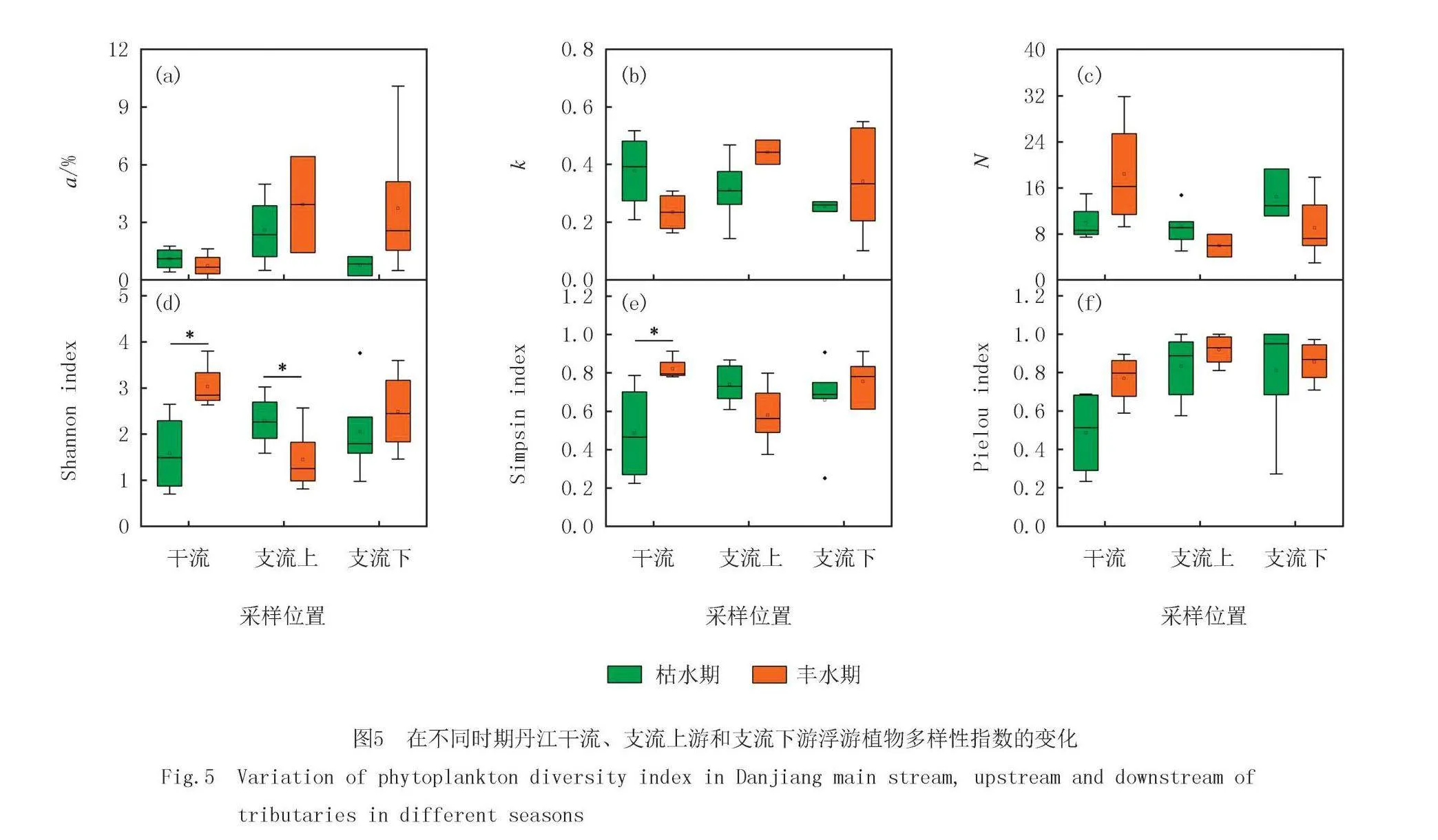
丹江干流浮游植物多样性指数a值枯水期和丰水期较低分别是1.1%和0.7%,支流丰水期的a值较枯水期更高,尤其是支流上游的a值更高.枯水期和丰水期k值均值的变化分别是0.26~0.37和0.23~0.44,不同空间丰水期的k值波动更大.丹江干流和支流下游的N值更高,支流上游的N值更低(图5).
浮游植物α多样性指数包括香农多样性指数、辛普森优势度指数和皮卢均匀度指数.丹江干流和支流下游丰水期的香浓指数较高,干流不同时期香农指数差异性显著.支流上游枯水期的香农指数显著高于丰水期.辛普森指数支流不同时期差异性不显著,干流丰水期显著高于枯水期.支流的皮卢指数要高于干流,干流枯水期和丰水期的皮卢指数分别是0.48和0.77(图5).
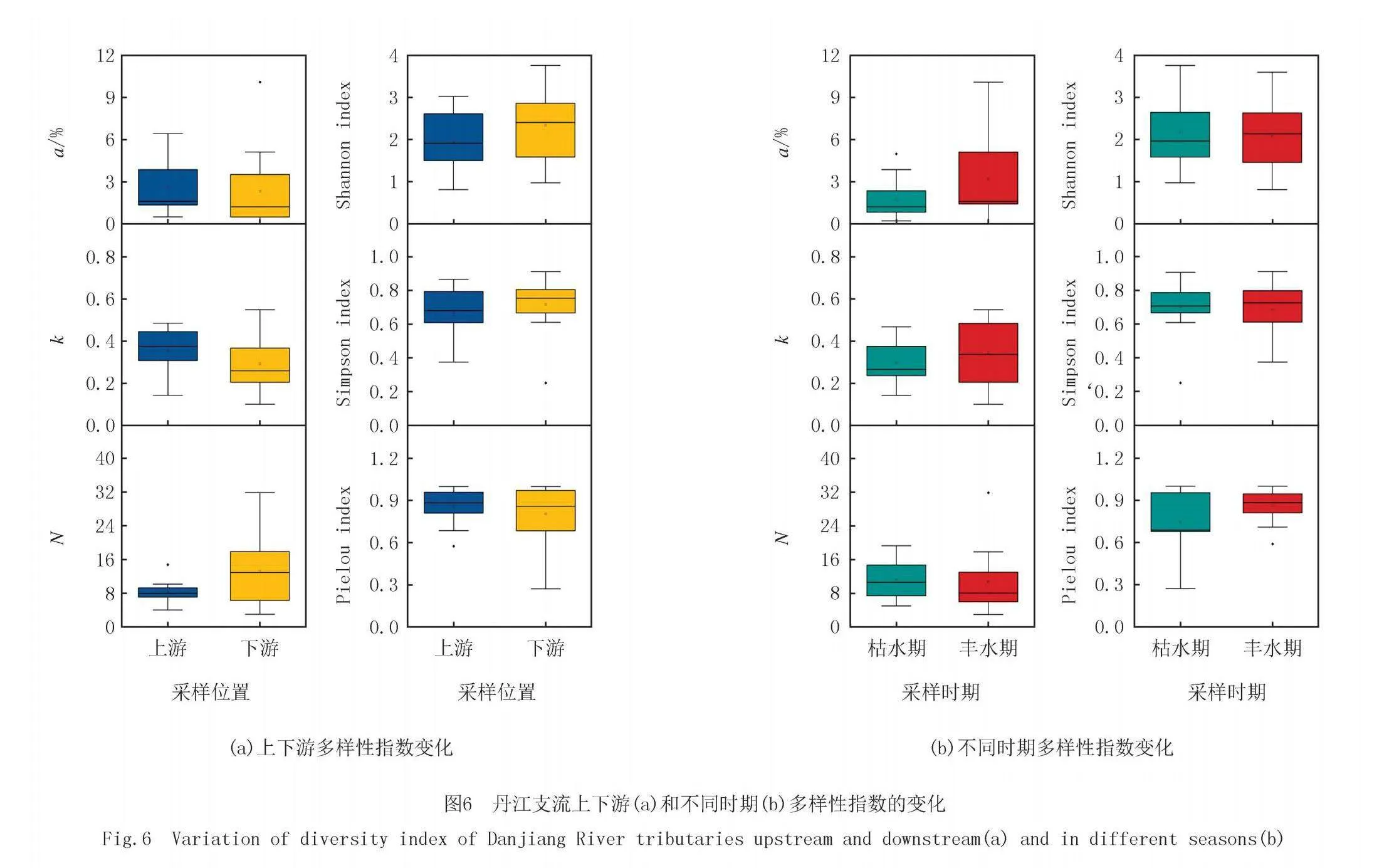
丹江支流上游和下游,枯水期和丰水期的多样性指数如图6所示.支流上游a和k比下游高,下游的N比上游的高.支流下游香农指数和辛普森指数率大于上游,上下游的皮卢指数相近.丰水期a和k高于枯水期,而枯水期和丰水期的N相近.枯水期和丰水期的香农指数和辛普森指数相近,丰水期的皮卢指数高于枯水期.
2.4 浮游植物群落的时间变化
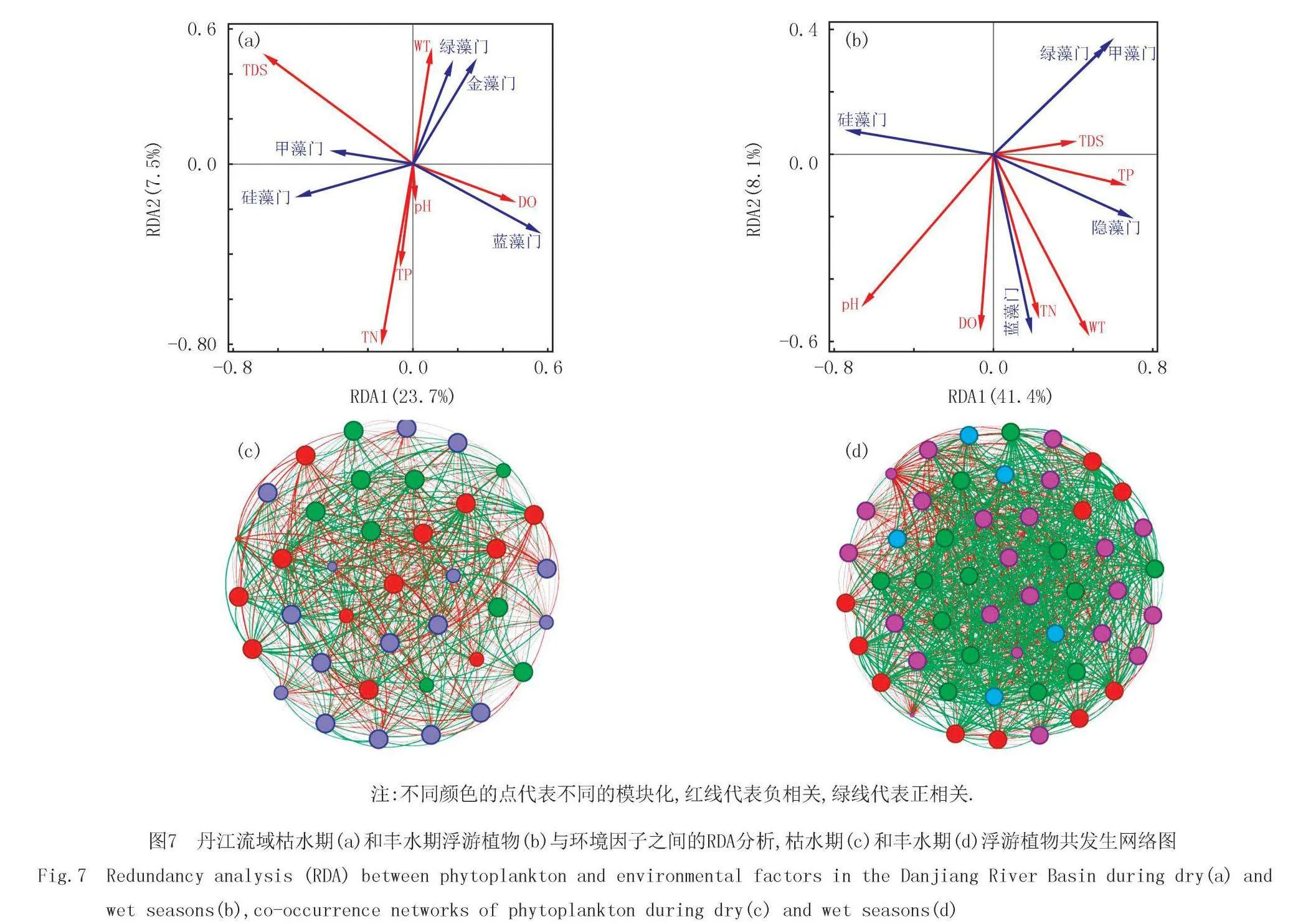
枯水期轴一和轴二的解释变量分别为23.7%和7.5%,累计解释变量为31.2%.丰水期轴一和轴二的解释变量分别为41.4%和8.1%,累计解释变量为49.5%(图7).本研究结果表明,丰水期环境因子对浮游植物群落的解释度更高.枯水期绿藻门和蓝藻门主要受WT和DO的解释度较高,甲藻门主要受TDS的解释度较高,而硅藻门受到的环境解释度较低.丰水期蓝藻门受到的环境因子解释度较大,与pH、DO、TN和TP均呈正相关,隐藻门主要受TP的解释度较高,绿藻门和甲藻门主要受pH的解释度较高且与pH呈负相关,硅藻门与TP呈负相关.
枯水期网络的节点和边数分别为38和697,丰水期网络的节点和边数分别为52和1 324.模块是网络中高度连接的区域,被认为是系统发育、进化或功能上独立的单元.更多的模块通常意味着网络中的节点具有更复杂的分组模式,可能对应不同的功能或生态位[22].丰水期的模块数是4,枯水期的模块数是3,可以将其解释为丰水期较枯水期具有更多的生态位.丰水期浮游植物物种数要高于枯水期.丰水期浮游植物节点之间的连线更趋向于正相关,说明丰水期浮游植物之间的协同作用更强.
2.5 浮游植物群落的空间变化
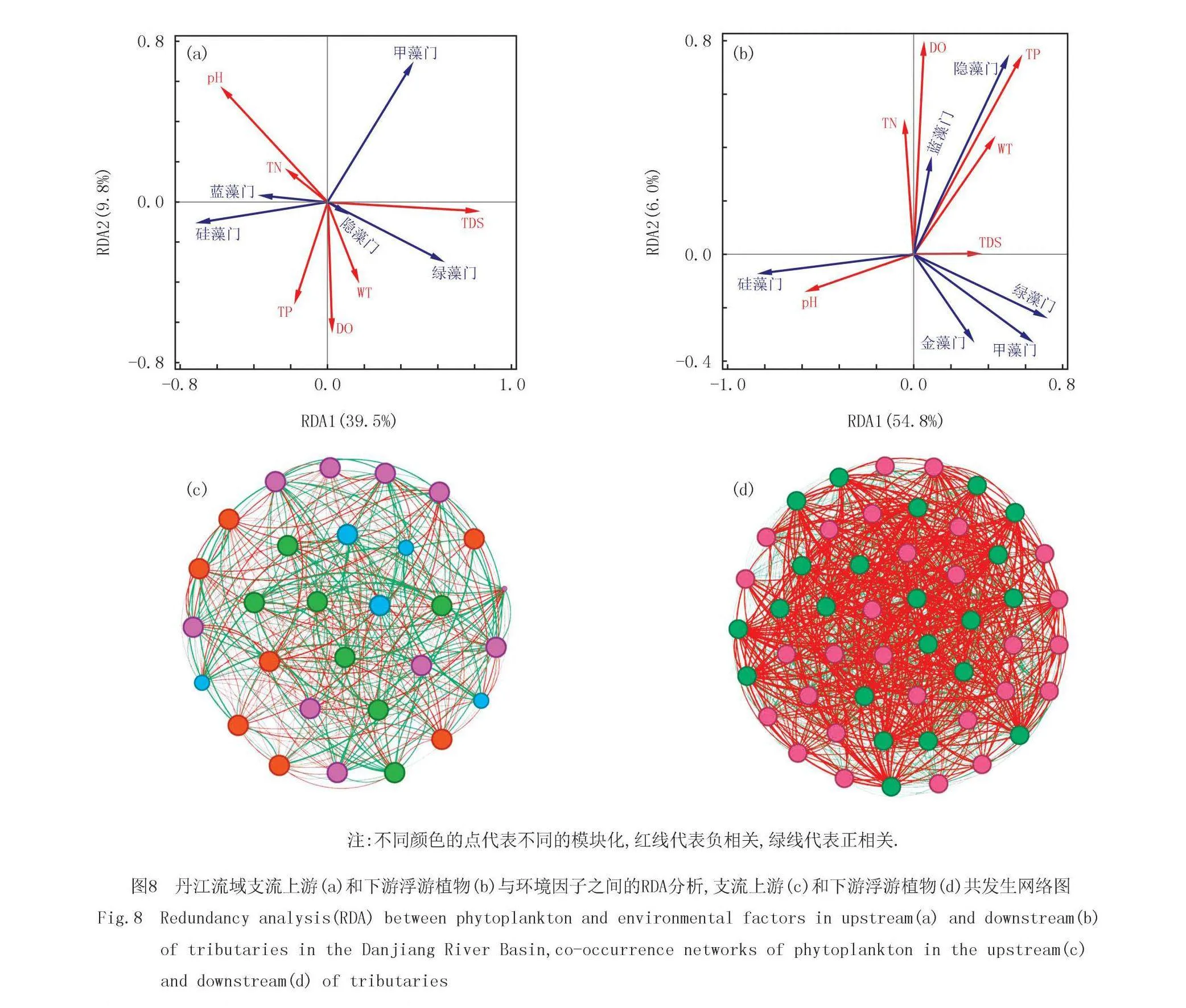
支流上游轴一和轴二的解释变量分别是39.5%和9.8%,累计解释变量为49.3%.支流下游轴一和轴二的解释变量分别是54.8%和6.0%,累计解释变量为60.8%(图8).下游环境因子对浮游植物群落的解释度更高.上游pH和TP对蓝藻门和硅藻门的解释度更高且呈正相关关系.TDS和pH对绿藻门的解释度更高,TDS与绿藻门呈正相关,而pH则与绿藻门呈负相关.下游蓝藻门和隐藻门与TN、DO、TP、WT和TDS均呈正相关关系,其中DO和TP解释度最高.pH对硅藻解释度高,且二者呈正相关关系.
支流上游网络的节点和边数分别为29和403,支流下游网络的节点和边数分别为50和1 225.支流上游的模块数是4,而下游的模块数仅为2.下游浮游植物节点之间的连线更趋向于负相关,可能是由于下游的物种种类多,浮游植物之间的竞争更强.
2.6 浮游植物群落与环境因子的关系
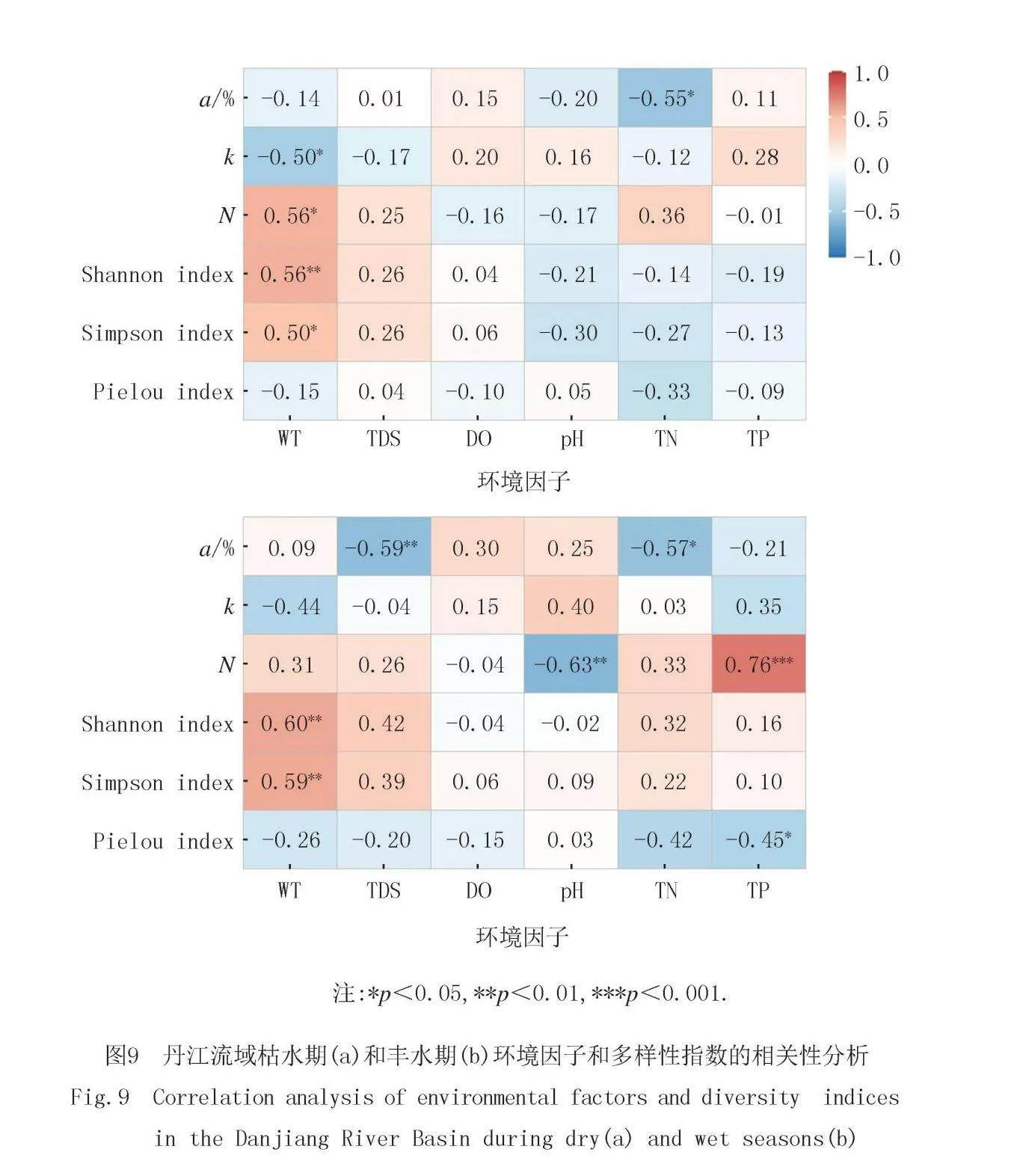
丹江流域枯水期和丰水期浮游植物多样性指数和环境因子的皮尔逊相关性如图9所示.WT与多样性指数更相关,且在枯水期更显著.枯水期WT与k、N、香浓指数和辛普森指数显著相关,TN与a显著负相关.丰水期WT与香农指数和辛普森指数显著正相关,TDS与a显著负相关,pH与N显著负相关,TN与a显著负相关,TP与N显著正相关而与皮卢指数显著负相关.
3 讨 论
3.1 丹江流域浮游植物的时间变化特征
本研究结果表明丹江流域河流主要以硅藻门为主,其次是蓝藻门和绿藻门.陈媛媛等[15]也发现相似的结果,研究发现丹江浮游植物主要以硅藻门为主,占58.33%,其次是蓝藻门占30.56%.首先,硅藻耐受力极强,能够在广泛的环境条件下生存[23].其次,硅藻对硅的需求较高,而河流中的硅通常充足,因为硅源自岩石和土壤的风化和侵蚀,这些物质通过河流不断补充到水体中.最后,河流中许多硅藻能够附着在河床、石头、植物等基质上,这种附着生活方式使它们能够在流水环境中稳定生存,而对于其他的微藻来说,更容易顺流而下迁移到水库或湖泊中.这些因素共同作用,使得硅藻在河流生态系统中占据重要地位,成为河流中常见且数量较多的种类.
本研究发现枯水期和丰水期均以硅藻门占优,其次是蓝藻门和绿藻门.枯水期硅藻门的占比比丰水期更高,而蓝藻门和绿藻门丰水期占比更高.马宝珊等[24]对安宁河浮游植物进行了枯水期和丰水期的调查,发现枯水期和丰水期均以硅藻门占优,其次是蓝藻门和绿藻门,其他类群较少,研究结果与本研究结果一致.赵耿楠等[25]对春秋两季渭河流域干流及秦岭北麓5条典型支流的浮游植物进行了调查,发现春季主要是以硅藻门占优,秋季硅藻门占优,其次是绿藻门,甲藻门和隐藻门.总的来说,枯水期硅藻门更多,可能是由于温度的原因,硅藻的最适温度要比蓝绿藻更低,丰水期温度适合大多数的藻生长,所以在丰水时期,河流中除了硅藻门占优势外,藻类更加丰富,蓝藻、绿藻、甲藻等均会在这个时期进行繁殖.本研究网络分析图也得到了相似的规律,在枯水期藻类的种类更低,且大部分均为硅藻.丰水期网络分析中的节点、边数和模块数均高于枯水期,说明丰水期的藻类种类更加丰富,生态位更多,适合更多的藻生长.并且在丰水期藻之间的连线更趋向于正相关,说明丰水期浮游植物之间的协同作用更强,群落更稳定.
多样性指数是反映浮游植物群落结构的重要参数,它包含物种丰富度和物种均匀度两个方面的含义[18-20].本研究中主要用香浓指数、辛普森指数和皮卢指数描述浮游植物群落的结构特征.为了更全面地探究浮游植物群落的结构特征,本研究计算了YE等[14]提出的3个多样性指数(a、k和N),其中a值表示濒危物种丰度占比,k值表示竞争强度,N值表示物种容纳量,结合上述指数对浮游植物群落结构进行更全面的评价.结果发现丰水期的a值和k值均值均高于枯水期,而N则相反.丰水期的温度更适合藻类的生长,稀有种可以利用更多的生态位.枯水期和丰水期的香农指数和辛普森指数均值差异不大,丰水期皮卢指数均值要略高于枯水期,可能是由于各种藻类大量繁殖,群落中藻的丰度差异减小.
3.2 丹江流域浮游植物的空间变化特征
本研究结果发现丹江干流硅藻和蓝藻占比更高,其次是绿藻和隐藻.丹江支流硅藻占比最高,其次是绿藻和蓝藻.丹江干流比支流藻的种类更加丰富,尤其是在丰水期.首先,丹江干流水量较大、流速较慢,这种环境有利于浮游植物的繁殖和分布,而支流流速较快,不利于浮游植物的生长[26].其次,干流区域流域面积大,上游带来的营养物质易在此处积累,为浮游植物的生长提供了充足的养分[27].最后,干流区域的水体通常较为稳定,水质条件变化较小,适合浮游植物生长[28],而支流常受到更多环境因素的影响,如降水、地形变化等,导致了水体不同位置水质的差异较大,浮游植物生长受到更大的扰动[29].因此,以上的原因可能会导致干流的物种丰富度更多且蓝藻和绿藻的占比增加.
丹江支流上游的硅藻占比下游更高,下游的蓝藻和绿藻占比更高.支流上游网络分为4个模块,而下游仅分为2个模块且浮游植物之间主要呈负相关关系.首先,支流上游来源于山泉水,水温较低,不利于某些温暖水域浮游植物的生长,而硅藻的最适温度要低于蓝绿藻,下游水温会更高[10],因此,上游硅藻更多.其次,上游水体流速较快,浮游生长的藻类难以稳定大量的繁殖,检测到的硅藻可能来源于水流冲击底栖的硅藻,硅藻顺流而下到河流的下游,下游河道变宽,流速变缓,适合藻类进行大量繁殖[30].此外,由于降水原因,土壤中的藻类会随着雨水冲刷而被携带到水体中[31],因此,上下游周边不同的土地利用类型也会影响水体中藻类的组成[32].上游人类活动干扰较弱,而到了下游人类活动干扰加剧[33],下游的点源和面源污染加强,导致水体中氮磷等营养物质增加,水体中的藻类大量繁殖[34].因此,上述原因可能会导致上游硅藻多,而下游蓝绿藻增加.
上游a值和k值均值高于下游,下游的N值高于上游.上游浮游植物濒危临界占比高说明上游中的稀有种更容易存活,但是由于上游营养物质少并且温度较低,加剧了浮游植物间的竞争强度.下游水体中由于人类活动等影响,水体中营养物质增加并且温度更适合大部分藻的生长,因此下游的物种丰富度更高.丹江流域支流下游香农指数和辛普森指数均高于上游.李庆南等[35]对辽河太子河水系的浮游植物进行了采集分析,发现不同区域浮游植物多样性的差异性显著,太子河水系上游地区浮游植物多样性小于下游地区.西枝江流域上游浮游植物多样性指数小于下游[36].上述研究与本研究得到的结果相同.
3.3 浮游植物群落对环境因子的响应
浮游植物群落结构的变化与环境因子紧密相关,且不同时间和空间水体中环境因素对群落结构的影响也存在差异.本研究发现WT与多样性指数更相关,且在枯水期更显著.丹江流域枯水期WT与k、N、香浓指数和辛普森指数显著相关,TN与a显著负相关.丰水期WT与香农指数和辛普森指数显著正相关,TDS与a显著负相关,pH与N显著负相关,TN与a显著负相关,TP与N显著正相关而与皮卢指数显著负相关.丹江流域枯水期浮游植物结构变化的主要影响因子为TN和TDS,丰水期浮游植物结构变化的主要影响因子为WT、pH、DO和TN.上游pH和TP对蓝藻门和硅藻门的影响较大且呈正相关关系.TDS和pH对绿藻门的影响较大,TDS与绿藻门呈正相关,而pH则与绿藻门呈负相关.下游蓝藻门和隐藻门与TN、DO、TP、WT和TDS均呈正相关关系,其中DO和TP影响最大.pH对硅藻影响大,且二者呈正相关关系.综上所述,本研究中丹江流域浮游植物群落主要受到WT、TN和TP环境因子的调控.
水温是影响浮游植物群落组成和分布的关键因子.本研究丹江流域枯水期和支流上游的水温较低,浮游植物主要以硅藻占优,而在丰水期和下游蓝藻和绿藻的占比会增多.水温直接影响浮游植物的生长和代谢速率.随着温度升高,浮游植物的光合作用和呼吸作用速率也会增加,但只有在最适温度范围内才能最大化生长和繁殖[37].超出此范围的高温会导致代谢失衡,影响浮游植物的生长情况[38].浮游植物在不同温度下对营养物质的利用效率不同,这会影响它们在不同温度条件下的竞争能力.不同种类的浮游植物对温度有不同的适应性.一般来说,水温升高会加速浮游植物的生长率,但每种浮游植物都有其最适生长温度范围[39].超过这个范围,浮游植物的生长率可能会下降,甚至导致死亡.对于硅藻来说,生长的最适温度15~25 ℃,而蓝绿藻的最适温度范围更高在25~30 ℃[40].因此,河流硅藻在枯水期以及上游丰度占比更高.
营养盐是浮游植物生长所必须的营养元素[10].氮元素是叶绿素的组成部分,而叶绿素是光合作用中吸收光能的关键色素.磷元素通过多种机制影响藻类的生理和代谢,包括能量代谢、核酸合成、细胞膜结构、细胞分裂和生长、代谢调控以及生态和环境适应[41].本研究中浮游植物群落的结构组成与TN和TP相关.大量研究也发现氮磷与浮游植物密切相关,例如,闵文武等[42]研究发现渭河水系丰水期浮游植物群落结构的主要驱动因子为DO和TN,枯水期为流速、TN和COD.有研究发现蓝藻、绿藻对磷需求较高,蓝藻适宜低氮环境,绿藻偏好氮磷比较高的环境,硅藻适宜较高的硅磷比和氮磷比[43].君珊等[44]研究发现拉萨河中舟形藻、桥弯藻及纤维藻对TN响应显著,水绵、颤藻、异极藻、曲壳藻等物种对TP响应明显.有研究高韦里河的TDS在25~340 mg·L-1[45],一般来说河流中的TDS波动小,不会对浮游植物群落造成很大的扰动,一般淡水河流中的TDS超过1 000 mg·L-1时,浮游植物会受到抑制,本研究中的TDS变化在70~260 mg·L-1,因此对浮游植物群落的结构不会造成影响.然而,由于人类活动加剧,导致氮磷等营养物质输入到河流中,会造成有害藻的大量生长而引起蓝藻水华,危害水域生态系统的健康.
4 结 论
1)丹江流域共鉴定出浮游植物7门62属104种,主要以硅藻门为主.枯水期和丰水期丹江流域优势藻均为曲壳藻.前20优势藻中其中11种为硅藻门,4种为绿藻门,2种为蓝藻门,2种为隐藻门和1种为裸藻门.
2)本研究发现WT与多样性指数更相关,且在枯水期更显著.丹江流域枯水期WT与k、N、香浓指数和辛普森指数显著相关,TN与a显著负相关.丰水期WT与香农指数和辛普森指数显著正相关,TDS与a显著负相关,pH与N显著负相关,TN与a显著负相关,TP与N显著正相关而与皮卢指数显著负相关.
3)丹江流域枯水期浮游植物结构变化的主要影响因子为TN,丰水期浮游植物结构变化的主要影响因子为WT、pH、DO和TN.本研究中丹江流域浮游植物群落主要受到WT、TN和TP环境因子的调控.
4)丰水期网络的节点、边和模块化较枯水期更高且浮游植物之间主要呈现正相关系.上游网络分为4个模块,而下游仅分为2个模块且浮游植物之间主要呈负相关关系.丰水期的群落更复杂且更稳定,而下游群落竞争强度大,容易导致稀有种的灭绝.
附录见电子版(DOI:10.16366/j.cnki.1000-2367.2024.06.14.0002).
参 考 文 献
[1] "NASELLI-FLORES L,PADISK J.Ecosystem services provided by marine and freshwater phytoplankton[J].Hydrobiologia,2023,850(12/13):2691-2706.
[2]宋高飞,张春梅,朱宇轩,等.南水北调中线浮游植物群落分布特征及其影响因子[J].河南师范大学学报(自然科学版),2023,51(4):17-26.
SONG G F,ZHANG C M,ZHU Y X,et al.Distribution characteristics of phytoplankton communities and their influencing factors along the Middle Route of South-to-North Water Diversion Project[J].Journal of Henan Normal University(Natural Science Edition),2023,51(4):17-26.
[3]HENSON S A,CAEL B B,ALLEN S R,et al.Future phytoplankton diversity in a changing climate[J].Nature Communications,2021,12(1):5372.
[4]SOMMER U,ADRIAN R,DE SENERPONT DOMIS L,et al.Beyond the plankton ecology group(PEG)model:mechanisms driving plankton succession[J].Annual Review of Ecology,Evolution,and Systematics,2012,43:429-448.
[5]白海锋,王怡睿,宋进喜,等.渭河陕西段浮游植物群落结构时空变化与影响因子分析[J].环境科学学报,2021,41(8):3290-3301.
BAI H F,WANG Y R,SONG J X,et al.Spatio-temporal characteristics and influencing factors of phytoplankton community structure in the Shaanxi Section of Weihe River,China[J].Acta Scientiae Circumstantiae,2021,41(8):3290-3301.
[6]HERING D,JOHNSON R K,KRAMM S,et al.Assessment of European streams with diatoms,macrophytes,macroinvertebrates and fish:a comparative metric-based analysis of organism response to stress[J].Freshwater Biology,2006,51(9):1757-1785.
[7]胡金,万云,洪涛,等.基于河流物理化学和生物指数的沙颍河流域水生态健康评价[J].应用与环境生物学报,2015,21(5):783-790.
HU J,WAN Y,HONG T,et al.Aquatic ecological health assessment of Shaying River Basin based on indices of physico-chemistry and aquatic organisms[J].Chinese Journal of Applied and Environmental Biology,2015,21(5):783-790.
[8]徐菲,王永刚,张楠,等.河流生态修复相关研究进展[J].生态环境学报,2014,23(3):515-520.
XU F,WANG Y G,ZHANG N,et al.Advances in the assessment of river ecological restoration[J].Ecology and Environmental Sciences,2014,23(3):515-520.
[9]邱阳凌,林育青,刘俊杰,等.淮河干流及主要支流夏季浮游植物群落生物多样性评价[J].环境科学学报,2018,38(4):1665-1672.
QIU Y L,LIN Y Q,LIU J J,et al.The biodiversity assessment of phytoplankton community in summer within main stream and tributary of Huaihe River[J].Acta Scientiae Circumstantiae,2018,38(4):1665-1672.
[10]施军琼,何书晗,欧阳添,等.三峡库区支流花溪河浮游植物种间关联及影响因子分析[J].河南师范大学学报(自然科学版),2022,50(5):110-116.
SHI J Q,HE S H,OUYANG T,et al.Analysis of phytoplankton interspecific association and its influencing factor in Huaxi River,a tributary of the Three Gorges Reservoir[J].Journal of Henan Normal University(Natural Science Edition),2022,50(5):110-116.
[11]窦祥洲,钱秀红,潘维艳,等.小清河入海口河段水质评价及主要污染物分析[J].灌溉排水学报, 2023,42(8):114-120.
DOU X Z,QIAN X H,PAN W Y,et al.Pollutants in the Estuary of Xiaoqing River: Analysis and Water Quality Assessment[J].Journal of Irrigation and Drainage,2023,42(8):114-120.
[12]PACHES M,AGUADO D,MARTNEZ-GUIJARRO R,et al.Long-term study of seasonal changes in phytoplankton community structure in the western Mediterranean(Valencian Community)[J].Environmental Science and Pollution Research International,2019,26(14):14266-14276.
[13]SHAMPRITY P,ATAUL G M,ALMUJADDADE A M,et al.Seasonality of phytoplankton and their relationship with some environmental factors in a pond of old Dhaka[J].BANGLADESH JOURNAL OF BOTANY,2016,45(1):195-201.
[14]YE S S,WEN L,GAO L,et al.Exploring intrinsic distribution of phytoplankton relative abundance and biomass in combination with continental-scale field investigation and microcosm experiment[J].Water Research,2024,248:120853.
[15]陈媛媛,张建军,张军燕,等.丹江陕西段春季浮游植物的群落结构特征[J].安徽农业科学,2012,40(32):15726-15728.
CHEN Y Y,ZHANG J J,ZHANG J Y,et al.Community structure characteristics of phytoplankton in April in Shaanxi section of Danjiang River[J].Journal of Anhui Agricultural Sciences,2012,40(32):15726-15728.
[16]国家环境保护总局水和废水监测分析方法编委会.水和废水监测分析方法[M].4版.北京:中国环境科学出版社,2002.
[17]胡鸿钧,魏印心.中国淡水藻类:系统、分类及生态[M].北京:科学出版社,2006.
[18]SHANNON C E.A mathematical theory of communication[J].Bell System Technical Journal,1948,27(3):379-423.
[19]SIMPSON E H.Measurement of diversity[J].Nature,1949,163(4148):688.
[20]PIELOU E C.The measurement of diversity in different types of biological collections[J].Journal of Theoretical Biology,1966,13:131-144.
[21]赵文.水生生物学[M].北京:中国农业出版社,2005.
[22]OLESEN J M,BASCOMPTE J,DUPONT Y L,et al.The modularity of pollination networks[J].Proceedings of the National Academy of Sciences of the United States of America,2007,104(50):19891-19896.
[23]黎佛林,蔡德所,唐鑫,等.硅藻指数筛选及水质多指标评价体系构建[J].长江科学院院报,2015,32(8):26-33.
LI F L,CAI D S,TANG X,et al.Diatom index screening and multi-index system to assess water quality[J].Journal of Yangtze River Scientific Research Institute,2015,32(8):26-33.
[24]马宝珊,徐滨,魏开金,等.安宁河中游浮游植物群落结构及其与环境因子的关系[J].生态学杂志,2020,39(10):3332-3341.
MA B S,XU B,WEI K J,et al.Phytoplankton community structure and its relation to environmental conditions in the middle Anning River,China[J].Chinese Journal of Ecology,2020,39(10):3332-3341.
[25]赵耿楠,潘保柱,丁一桐,等.渭河干流和秦岭北麓典型支流浮游植物功能群特征及水质评价[J].生态学报,2021,41(8):3226-3237.
ZHAO G N,PAN B Z,DING Y T,et al.Characteristics and water quality evaluation of phytoplankton functional groups in the Weihe River mainstem and its tributaries in the northern foot of the Qinling Mountains[J].Acta Ecologica Sinica,2021,41(8):3226-3237.
[26]贺玉晓,郑永坤,李卫国,等.丹江口水库早春真核浮游植物群落结构特征及其与环境因子的关系[J].环境科学学报,2021,41(6):2192-2200.
HE Y X,ZHENG Y K,LI W G,et al.Characteristics of eukaryotic phytoplankton community structure in early spring and its relationship with environmental factors in Danjiangkou Reservoir[J].Acta Scientiae Circumstantiae,2021,41(6):2192-2200.
[27]HUTCHINS M G,JOHNSON A C,DEFLANDRE-VLANDAS A,et al.Which offers more scope to suppress river phytoplankton blooms:reducing nutrient pollution or riparian shading?[J].Science of the Total Environment,2010,408(21):5065-5077.
[28]XU Y G,LI A J,QIN J H,et al.Seasonal patterns of water quality and phytoplankton dynamics in surface waters in Guangzhou and Foshan,China[J].Science of the Total Environment,2017,590:361-369.
[29]HOU Z J,JIANG Y,LIU Q,et al.Impacts of environmental variables on a phytoplankton community:a case study of the tributaries of a subtropical river,Southern China[J].Water,2018,10(2):152.
[30]周静,苟婷,张洛红,等.流速对不同浮游藻类的生长影响研究[J].生态科学,2018,37(6):75-82.
ZHOU J,GOU T,ZHANG L H,et al.The effect of flow velocity on the growth of different phytoplankton[J].Ecological Science,2018,37(6):75-82.
[31]PAERL H W,OTTEN T G,KUDELA R.Mitigating the expansion of harmful algal blooms across the freshwater-to-marine continuum[J].Environmental Science amp; Technology,2018,52(10):5519-5529.
[32]秦明慧,刘秀丽.海河流域水环境安全评价及动态耦合协调度分析[J].灌溉排水学报,2023, 42(10):63-73.
QIN M H,LIU X L.Evaluation of Water Environmental Security in the Haihe River Basin and Analysis of Dynamic Coupling Coordination[J].Journal of Irrigation and Drainage,2023,42(10):63-73.
[33]RANGECROFT S,VAN LOON A F,MAUREIRA H,et al.An observation-based method to quantify the human influence on hydrological drought:upstream-downstream comparison[J].Hydrological Sciences Journal,2019,64(3):276-87.
[34]WU Y P,CHEN J.Investigating the effects of point source and nonpoint source pollution on the water quality of the East River(Dongjiang) in South China[J].Ecological Indicators,2013,32:294-304.
[35]李庆南,赵文,殷旭旺,等.辽河太子河水系的浮游植物种类多样性研究[J].大连海洋大学学报,2011,26(4):322-327.
LI Q N,ZHAO W,YIN X W,et al.Phytoplankton diversity in taizi river tributary in Liao River[J].Journal of Dalian Ocean University,2011,26(4):322-327.
[36]李思阳,张娟,姚玲爱,等.西枝江流域浮游植物群落结构特征与主要环境因子的关系研究[J].环境科学学报,2016,36(6):1939-1947.
LI S Y,ZHANG J,YAO L A,et al.The community structure of phytoplankton and its relationship with environmental factors in Xizhijiang River[J].Acta Scientiae Circumstantiae,2016,36(6):1939-1947.
[37]王锦旗,宋玉芝,黄进.水温升高对水体性质及水生生物的影响研究进展[J].水生态学杂志,2020,41(1):100-109.
WANG J Q,SONG Y Z,HUANG J.Effects of increasing water temperature on water properties and aquatic organisms:a critical review[J].Journal of Hydroecology,2020,41(1):100-109.
[38]廖一波,陈全震,曾江宁,等.海洋浮游植物的热效应[J].生态学报,2008,28(9):4203-4212.
LIAO Y B,CHEN Q Z,ZENG J N,et al.Thermal effect on the marine phytoplankton in Xiangshan Bay,China[J].Acta Ecologica Sinica,2008,28(9):4203-4212.
[39]COFFEY R,PAUL M,STAMP J,et al.A review of water quality responses to air temperature and precipitation changes:nutrients,algal blooms,sediment,pathogens[J].Journal of the American Water Resources Association,2018,55(4):844-868.
[40]邵海燕,王卿,高春霞,等.长江口浮游植物群落特征及影响因素分析[J].大连海洋大学学报,2024,39(1):124-133.
SHAO H Y,WANG Q,GAO C X,et al.Analysis of phytoplankton community characteristics and influencing factors in the Yangtze River Estuary[J].Journal of Dalian Ocean University,2024,39(1):124-133.
[41]QU Z,DUAN P F,CAO X B,et al.Comparison of monoculture and mixed culture(Scenedesmus obliquus and wild algae) for C,N,and P removal and lipid production[J].Environmental Science and Pollution Research International,2019,26(20):20961-20968.
[42]闵文武,王培培,李丽娟,等.渭河流域浮游植物功能群与环境因子的关系[J].环境科学研究,2015,28(9):1397-1406.
MIN W W,WANG P P,LI L J,et al.Relationship between phytoplankton functional groups and environmental factors in the Wei River Basin[J].Research of Environmental Sciences,2015,28(9):1397-1406.
[43]DORTCH Q,WHITLEDGE T E.Does nitrogen or silicon limit phytoplankton production in the Mississippi River plume and nearby regions?[J].Continental Shelf Research,1992,12(11):1293-1309.
[44]君珊,王东波,周健华,等.拉萨河流域浮游植物群落结构特征及与环境因子的关系[J].生态学报,2019,39(3):787-798.
JUN S,WANG D B,ZHOU J H,et al.Community structures of phytoplankton and its relationship with environmental factors in the Lhasa River[J].Acta Ecologica Sinica,2019,39(3):787-798.
[45]GHOLAMI S,SHIVANNA S.Analysis of agricultural impact on the Cauvery River water around KRS dam[J].World Appl Sci J,2009,6(8):1157-1169.
Temporal and spatial changes and driving factors of phytoplankton communities in the Danjiang River Basin
Li Ming1, Ye Sisi1, Li Juan2, Xue Xudong2, Hu En3
(1. College of Natural Resources and Environment, Northwest Aamp;F University, Yangling 712100, China; 2. Shaanxi Environmental Investigation and Assessment Center, Xi'an 710061, China; 3. Shaanxi Provincial Academy of Environmental Science, Xi'an 710061, China)
Abstract: Phytoplankton serves as an important indicator species in aquatic ecosystem, with its community composition and distribution closely linked to environmental factors. The Danjiang River is the primary water source for the Danjiangkou Reservoir, making it crucial to study the spatial and temporal variations of phytoplankton and their driving factors to protect the water resources of the Danjiang River Basin. In this study, water and phytoplankton samples were collected from the main stream and six tributaries of the Danjiang River during both dry and wet seasons. Phytoplankton species were identified and analyzed to understand their community structure's response to environmental factors over time and space. In the Danjiang River Basin, 104 species of phytoplankton across 7 phyla and 62 genera were identified, with diatoms being the predominant group. Achnanthes was the dominant genus during both dry and wet periods. Among the top 20 dominant algae, 11 species were diatoms. Water temperature(WT) in the Danjiang Basin showed a stronger correlation with the diversity index, especially during the dry season. In the dry season, total nitrogen(TN) was the main factor influencing phytoplankton structures changes, while in the wet season, WT, pH, dissolved oxygen(DO), and TN were the primary influencing factors. The phytoplankton community in the Danjiang Basin was mainly regulated by WT, TN, and total phosphorus(TP). Network analysis revealed that nodes, edges, and modularity were higher during the wet season compared with the dry season, indicating stronger positive relationships among phytoplanktons. The upstream network consisted of four modules, whereas the downstream network had only two, with phytoplankton predominantly negatively correlated with each other. This study provides a theoretical basis for protecting the aquatic ecosystem of the Danjiang River Basin.
Keywords: Danjiang River Basin; phytoplankton; environmental factors; diversity index; temporal and spatial variation
[责任编校 刘洋 赵晓华]
猜你喜欢
农家科技下旬刊(2016年9期)2016-12-15 13:23:52
农业与技术(2016年20期)2016-12-08 21:22:42
商(2016年29期)2016-10-29 11:10:35
中国市场(2016年36期)2016-10-19 05:25:57
商场现代化(2016年12期)2016-06-06 16:46:42
中国科技博览(2016年20期)2016-04-25 14:11:10
河北渔业(2015年11期)2015-11-18 16:37:49
河北渔业(2015年10期)2015-10-15 11:14:57
河北渔业(2015年10期)2015-10-15 11:10:10
湖北农业科学(2015年18期)2015-10-13 21:36:01
