旧石器时代古人类和智人在东亚的扩散
2025-02-25 00:00:00RobinDENNELL
人类学学报 2025年1期





关键词:古人类;智人;扩散;东亚;生物地理学
1 Introduction
Investigations of human evolution within a biogeographical framework began very early inEast Asia1)[1]. The German zoologist Ernst Haeckel (1834-1919) proposed in 1868 that apes andhumans originated on a lost continent called Lemuria, which sank in the Indian Ocean: the ancestorsof chimpanzees and gorilla were then supposedly washed ashore in Africa and the ancestorsof humans and orangutans reached Southeast Asia, from which hominins evolved into humansand eventually dispersed worldwide. Although his scenario lacked any supporting evidence, itencouraged Eugene Dubois to seek employment as a doctor in the Dutch East Indies. Haeckel’stheory seemed confirmed in 1891 when Dubois found at Trinil, Java, the skull and femur that henamed Pithecanthropus, a term that Haeckel had proposed as the first human ancestor. In the early20th century, Henry Osborne (1857-1935) proposed that human origins lay in Central Asia (definedas Mongolia, Xizang and Inner Mongolia)[2, 3]. His ideas were expanded by the American researcherWilliam Diller Matthew (1871-1930), whose paper “Climate and Evolution” was published in1915[4]. In his model, the uplift of the Tibetan Plateau blocked the summer monsoon and causedthe drying out of Central Asia (defined primarily as Inner Mongolia and Xinjiang in China, andMongolia). As a result, the species inhabiting Central Asia would perish, adapt to drier conditions,or leave. However, the uplift of the Himalayas created a barrier that prevented species from movingto the forests of South and Southeast Asia. He further proposed that those species in the center ofthe “cradle” were dynamic and adaptable, but those on the edges were conservative and passive,and simply moved to familiar environments elsewhere. They were then displaced even further bythe more dynamic species in the center that had adapted to new, more arid conditions and wereable to radiate outwards and colonize new environments. As part of that narrative, these primitivetypes included the ancestors of gorillas and chimps that were displaced to Africa by the moredynamic hominins[5]. In Matthew’s view, “The most advanced stages should be nearest the centreof dispersal, the most conservative stages furthest from it”[6]. He effectively inverted Darwin’ssuggestion that humans evolved in Africa because that was where our closest cousins are found;for Matthew, the existence of the chimpanzee and gorilla in Africa indicated that they had beendispersed there by more advanced types (such as humans) that originated in Central Asia. Theseideas were developed by the Canadian Davidson Black (1884-1934), who sought employment atPeking Medical College in order to be near this potential centre of human evolution. His papers(andfurther discoveries at Zhoukoudian and localities such as the Nihewan Basin and Shuidonggouby Licent and Chardin in 1922) did much to establish Central Asia as the most likely cradle ofhumankind[6~9] . Just as Dubois discovered a human ancestor at Trinil, so Black was instrumental inrecognising Sinanthropus as a human ancestor in 1927 on the basis of one tooth from Zhoukoudian.It was only after WW2 that Australopithecus was recognised as a hominin and Africa was acceptedas a more likely place where hominins originated[10, 11].
Although the ideas of Haeckel, Matthew, Black and others about the primacy of East Asia inhuman origins are now historical curiosities, they are rare examples where theories about humanevolution were in place before discoveries appeared to confirm them. A century after the Swedishgeologist Andersson made the first discoveries at Zhoukoudian, it is appropriate to suggest howbiogeography can contribute to our understanding of the deep past of humanity in East Asia.
An enormous amount has been learnt in the last 20 years about the palaeolithic, Pleistoceneand human skeletal record of East Asia. As highlights, we can note that there are now good faunalrecords from the Arctic, Siberia, China, the Japanese islands and Southeast Asia; a superlativeclimatic record from the loess-palaeosol sequence of the Loess Plateau, supplemented by theloess-palaeosol record of Central Asia and marine cores from the south China Sea; and threenew hominin species (Denisovans, H. floresiensis and H. luzonenensis) have been discoveredin East Asia in the last 20 years[12, 13]. All this new information opens up opportunities for moreambitious objectives than writing culture sequences. Here, I propose that a biogeographicperspective could help illuminate the complex and rich record of hominin (including human)dispersals in Pleistocene East Asia. Central concerns of biogeography are to explain the presentdaydistribution of species, their origins and how and why their distribution changed in thepast. Palaeoanthropology has much the same agenda, but one that is complicated by the need toexplain how and when our own species became the only hominin on the planet.
Hominin dispersals and biogeography in East Asia
One reason why East Asia is such an exciting region for studying Pleistocene hominins isthat it is unique in encompassing two biogeographic Realms – the Oriental Realm of South Chinaand Southeast Asia (with a sub-realm of Wallacea) and the Palearctic Realm of North China,Mongolia, Siberia (with its own sub-Realm of the Arctic), and the Japanese Islands, with theQinling Mountains forming the boundary between the two in China (Fig.1) (For those unfamiliarwith the term, a biological Realm is an area of the earth’s surface that contains fauna and flora thatshare the same evolutionary history 2)). Hominins in the Palearctic Realm were usually dependenton seasonal plant foods and a small number of herbivores as prey that were often migratory,and sometimes over large distances (such as horse, reindeer and gazelle). Hunter-foragers in thePalaearctic Realm often faced considerable risk in their dependence on a small range of foodresources, and considerable uncertainty over their predictability[14]. The Oriental Realm containeda far larger range of plant resources, had a more equitable climate, but fewer herd animals thanthe Palearctic Realm, and there was less likelihood that a region would become uninhabitableduring a climatic downturn. Because diets were more broadly based in the Oriental Realm, therewas probably less risk of starvation if a key resource failed and a larger range of fall-back options.Consequently, hominin occupation in these two realms followed very different paths; populationsin each realm often developed in isolation from the other, but at other times, they interacted.Additionally, the boundaries of the Palearctic and Oriental Realms in China shifted when theclimate changed[15], so some animals such as panda (Ailuripoda) occasionally and Hystrix, Macacaand others frequently expanded their distribution north of the Qinling Mountains in warm periods,although only a few Palearctic species extended south of these mountains during cold ones[16].
There are several ways in which biogeography is a useful way of studying dispersalsof Pleistocene hominins in these Realms. Its main advantage is that it places homininswithin a climatic and environmental context that allows different types of evidence (skeletal,archaeological, faunal, botanical etc.) to be studied in an integrated manner. A biogeographicapproach is also appropriate for studying hominins at a continental or regional level and overlong time-spans of the Pleistocene. A third advantage is that it is a useful way of examiningthe opportunities and challenges faced by hominins when adapting to the different types ofenvironments in the Palearctic or Oriental Realms.
When dealing with hominins, two additional factors need considering. The first is thathominins (including humans) are unmatched in their capacity for adaptive change throughdevelopments in technology, social organisation and cognitive ability. Whereas biogeographicstudies of rabbits or rhinos, for example, can assume that their behaviour has not changedsubstantially in the last two million years, studies of hominins always have to incorporate theircapacity for adaptive change in behaviour. A second factor that greatly complicates the study ofhominin biogeography in East Asia is that it was occupied by several types of hominins for mostof the Pleistocene until replaced by Homo sapiens. In regions of East Asia already occupied byindigenous hominin populations, immigration, and population assimilation and/or replacement byH. sapiens are major themes when considering how humans became the sole hominin resident.
2 Biogeographic modelling of hominin dispersals in East Asia
Two sets of models can be used to explain past dispersals of hominins. The first are thosewhere the driving force is the environment – climatic and environmental change, or the behaviourof the fauna. The second set is unique to hominins, and are changes brought about by theiradaptive responses that led to changes in behaviour which in turn opened up new opportunitiesfor dispersal and colonisation.
2.1 Preliminary considerations
First, some points about terminology. A dispersal is the same as range extension – the process bywhich a species extends its distribution into areas previously unoccupied. Range extension always occursat the edge of an animal’s range and therefore usually involves directly only a small portion of the totalpopulation. Migration in bird and animal populations refers primarily to seasonal movements betweenwinter and summer territories; for example, migrating populations of deer or geese will make a two-wayjourney to and from a winter or summer feeding ground. Social scientists often use the terms “migration”
and “dispersal” as though they are interchangeable. This leads to confusion, and the term “migration”
should be restricted to discussions of seasonal, two-way movements. In modern human populations,migration is often used to mean people moving from one country or region to another without making areturn journey: with internal migration, from village to city inside a country, or with external migration,from Mexico to the United States, for example. Hence, we talk about immigration and emigration,although these should be more accurately be referred to as dispersals. Here, I confine “migration”
to meaning seasonal movements, and dispersals as implying an extension of an animal’s range.Metapopulations and palaeodemes
Human populations, like those of many other mammals, are structured in that the totalpopulation is composed of numerous sub-sets that are called metapopulations[18] (Fig. 2). These arespatially separated and inter-dependent parts of networks that share information, mating partners andsometimes scarce materials. Humans are extremely good at forming tight metapopulations that arelarge, complex, mutually supporting, and have a high degree of connectivity between them. In studiesof later prehistory, these networks have attracted much attention, and there are obvious connectionsbetween studies of archaeological networks and demographic metapopulations. In the LatePleistocene, these tight metapopulations were crucial factors in the colonisation of new areas suchas the Arctic, rainforests, Australia, Wallacea, the Japanese islands and the Americas. An unfortunatereality about Palaeolithic evidence is that it is exceptionally difficult to identify specific archaeologicalnetworks of metapopulations from lithic and (sometimes) bone, antler or artistic evidence.
Palaeodemes in human evolutionary studies are the equivalent of metapopulations. Howellstressed the importance of the palaeodeme as “the basic stuff of the hominin fossil record” as aunit of analysis in palaeoanthropology[19, 20]. A deme is “a communal interbreeding populationwithin a species …... distinguishable by reproductive (genetic), geographic, and ecological(habitat) parameters”. Together, demes (sometimes as isolated populations) constitute subspecies,or the “aggregate of local populations of a species inhabiting a geographic subdivision ofthe range of the species”[19]. China is one of the few regions of Asia where it is possible todemonstrate local palaeodemes from its detailed skeletal record[21].
2.2 Models of dispersal driven by environmental factors
We can begin by considering simple biogeographic models that are driven by changes inclimate and environment and which are applicable to all periods of the Pleistocene. Some ofthese are appropriate for studying north-south dispersals, and others for ones that run west toeast. The relevant background evidence is summarised in Dennell[12, 13].
Ebb and flow models
At a species level, an ebb and flow model is the simplest way of modelling the north-southdispersal of hominins within continental Asia in response to changes in climate. For those unfamiliarwith the sea or those for whom English is not the native language, ebb and flow refers to the twice-dailyfall and rise of the tide. The image of tidal range provides an appropriate way of envisaging the likelynorth and south movements of hominins in continental East Asia in response to climate change
In Europe and North America, the main factor determining the distribution of plants and animalswas the repeated advances and retreats of ice sheets that marked glacial and interglacial/interstadialperiods. As ice sheets and glaciers expanded in glacial periods, cold-intolerant species (such ashominins) retreated south into glacial refugia, and cold-tolerant species (such as Arctic lemmingsand musk oxen) expanded south from their interglacial refugia. When ice sheets contracted in warmperiods, hominins and other cold-intolerant species could expand out of their refugia and recoloniseareas previously abandoned, and cold-tolerant species retreated northwards into their Arctic biome.Consequently, the record of hominin occupation in Asia in the Early and Middle Pleistocene ispredominantly a record of occupation in warm periods and abandonment in cold ones[22, 23].
In East Asia, ice-sheets were far less significant than in northwest Europe and NorthAmerica, so the contrast between cold and warm periods is not defined by the limits of icesheets. Instead, the main contrasts in East Asia are mean annual temperature (MAT) and meanannual precipitation (MAP) that are determined by the relative strengths of the winter andsummer monsoons. In cold, dry periods, the winter monsoon pushes cold, dry air from Siberiaand Mongolia further south than in warm, moist periods, when the summer monsoon is able todeliver rain from the Pacific and Indian Oceans further north than in cold, dry periods. Theseshifts in rainfall determine the boundary between the arid and semi-arid regions of China andCentral Asia, and that in turn affects the distribution of hominins.
One area that shows a pattern of repeated hominin expansion and contraction in response toclimate change is the Chinese Loess Plateau, with its superlative sequence of 32 major loess andpalaeosol units denoting cold and warm periods. Today, mean annual temperature at Xifeng in theCentral Loess Plateau is 8.3 oC and mean annual precipitation is 560 mm. At its extremes, during theformation of palaeosol S4 (360-412 kaBP), S5-1 (479-531 kaBP) and S5-3 (585-621 kaBP), the LoessPlateau was sub-tropical and semi-humid, with 200-300 mm more rain than at present and 4-6 oCdegrees warmer[24]; when loess L9 (865-943 kaBP) and L15 (1.24-1.26 Ma) were deposited, itwas like a polar desert with an annual average temperature of only 1.3–3.0 oC degrees and only150-250 mm of rain[25]. During very warm and humid periods, panda browsed in the southernparts of the Loess Plateau[26], whereas during the Last Glacial Maximum(LGM) period, woollymammoth grazed as far south as the Qinling Mountains[27]. One would thus predict that theEarly and Middle Pleistocene hominin occupation of the Loess Plateau was largely confinedto warm periods. Field data conforms this. For example, fieldwork in Lantian county in thesouthern Loess Plateau shows that hominins were present in 17 warm periods but only 4 coldones between 2.1 and 1.2 MaBP[28]. In cold periods, the Plateau was largely abandoned in favourof refugia in valley basins in the Qinling Mountains such as the Luonan and Lushi, where ca.270 palaeolithic sites that date from 1.2 Ma to the last glacial cycle have been found[29-31]. Thesebasins were occupied in cold as well as warm periods. The Yangtse Valley was also anotherlikely glacial refugia for southern populations no longer able to inhabit northern China[32].
The deserts of North and Northwest China – the Northern Arid Area of China (NAAC) – isanother area where we would expect hominins to be present only during periods when rainfall washigher. The Nihewan Basin, which lies north of the Loess Plateau but south of the NAAC, is anotherarea that I have argued was occupied predominantly in the summer months of warm periods duringthe Early and Middle Pleistocene[33], and not continuously as suggested by some researchers[34, 35].
There is also an altitudinal element in this type of climate-induced dispersal: as snow linesretreated in warm periods, highland areas became inhabitable for warm-loving species, and coldtolerantones had to retreat to higher elevations. We might thus expect the hominin occupationrecord in high-elevation regions such as the Tibetan Plateau and Himalayas to be limited to warmerperiods when snowlines retreated. The site of Nwya Devu at 30-45 kaBP and 4,600 m in altitudemay be one example of short-term occupation of the Tibetan Plateau by H.sapiens during a warmpart of the last glacial cycle[36,37]. The Xiahe mandible from Baishiya Karst cave at 3280 m inaltitude is more problematic. Three U-series samples of carbonate from the base of the mandibleproduced dates of c. 160 kaBP, which is in the middle part of the cold period MIS 6. It seemsimprobable (at least to the author) that hominins were able to cope with both high elevations andlow temperatures, as severe as in MIS 4 and MIS2, as early as 160 kaBP. We need to understandbetter the length of the interval between the death of the Xiahe individual and the formation of thecarbonate crust. Additional dating might also confirm the current dating of 160 kaBP.
One major strength of ebb and flow models is that they are testable in two ways. Thefirst is to place sites in a climatic context, as in the Chinese Loess Plateau. Secondly, it may bepossible to obtain data on seasonality from faunal data, and estimates of temperature from proxyindicators such as insects or soil chemistry[38, 39].
Although a simple ebb and flow model is useful in modelling the dispersal of hominins acrosscontinental Asia and their movements northwards during warm, moist periods, it has three mainlimitations. The first is the assumption that when conditions worsen, populations retreated back intotheir refugia. This unlikely because when conditions worsened, all populations would have beenunder stress, including those in refugia. Instead of populations retreating into refugia, it is morelikely that there was a high rate of group extinction, particularly at the edge of the hominin range[40].This might have been occasionally catastrophic – for example, entire groups might perish in ablizzard or during an exceptionally cold winter – but was probably attritional in most cases. Smallgroups are highly vulnerable to extinction through random fatalities caused by accident or illness,by the small number of females that can bear children, by their rates of survival, and by inevitablefluctuations in food supply. Consequently, population levels are likely to have fallen sharply duringclimatic downturns. Tallavaara et al estimate that in Europe, populations totals decreased from ca.410,000 to 130,000 between 30 kaBP and 13 kaBP[41]. In a first attempt to model population levelsin China over the last 50 kaBP, Wang et al plotted radiocarbon dates from archaeological sites indifferent regions of China against the climatic record and inferred that population size was linkedto climate[42] (i.e., the number of dates per period was an approximation of relative population size).Unsurprisingly, they suggested that population levels fell sharply during cold, dry periods of the lastglacial cycle (They also emphasized that their conclusions needed testing with archaeological andenvironmental data). The second weakness of an ebb and flow model is that it treats hominins astotally passive when experiencing climatic and environmental change. Whilst it may be appropriateto model the behaviour of Early Pleistocene hominins in the same way as mice or mammoths, itis much less appropriate when discussing later hominins, who often used cultural adaptations thatenabled them to colonise regions that earlier hominins could not. As seen later, these adaptationshad profound consequences in East Asia. The third limitation of an ebb-and-flow model is that it isunable to explain complexity and variability in hominin skeletal and archaeological records. Here, asource-sink model is more effective (see below).
Habitat fragmentation, loss and recombination: barriers and corridors
Ebb and flow models can be effective ways of modelling hominin movements from northto south, but are less useful in explaining dispersals from east to west. One approach thatmay be useful is to consider how climate change creates barriers or corridors by habitat loss,fragmentation, or recombination. The most extreme example of habitat loss in PleistoceneEast Asia is the drowning of the Sunda Shelf during interglacials that halved the area of landand created numerous islands. In continental East Asia, areas of deserts might become semiaridgrassland during a warm, moist period, and the increased extent of grassland along desertmargins might open an east-west corridor for dispersal by hominins and other animals[43].Li Feng et al have suggested that ca. 35-45 kaBP[44], one such corridor open up that allowedH. sapiens to enter North China via the margins of the Taklamakan desert instead of fromTransbaikalia and Mongolia. In this scenario, humans would have relied upon the streams fed bymeltwater that flowed from the Kunlun Mountains to the south or the Tienshan to the north. Thissuggestion needs testing by fieldwork, but there are various indications that the Taklamakan wasat times more conducive to human settlement than now: Weiwen et al reported stone artefacts onthe southern edge of the Tarim Basin[45]; a grinding stone with starch grains of grasses and datedat 13 kaBP has been found in the hyper-arid Lop Nor Basin[46]; Han et al report a hearth and stoneartefacts dated to ca. 7.0-7.6 kaBP from the hyper-arid Tarim Basin[47].
As another example, areas of dense sub-tropical forest in South China or Southeast Asia mighthave fragmented under cooler and drier conditions, and the resulting mosaic landscape of denseforest, open woodland and perhaps some grassland would have created favourable conditions forhominins and also open corridors for dispersal[48]. Conversely, all these corridors could becomebarriers during periods of climatic change. It is important to note that habitat fragmentation thatbenefitted hominins could occur in both cold, dry periods and warm, moist ones: cold, dry periodswould have increased the extent of desert in Central Asia but created a mosaic landscape inSoutheast Asia, and warm, moist periods benefitted hominins in areas where deserts retreated, butnot necessarily those in areas that became dense sub-tropical and tropical forest or were threatenedby rising sea levels.
On a very large scale, ebb and flow models are useful in showing that range expansionnorthwards in central and east Asia became increasingly difficult in the Middle Pleistocenebecause the expansion of deserts converted much of what I previously called “Savannahstan” into“Aridistan”[12, 43, 49], and thereby created barriers against dispersal either north-south or west-east. InChina, the southern margin of the Mu Us Desert advanced southwards at 2.6, 1.2, 0.7 and 0.2 MaBP,possibly because of a stepwise weakening of the summer monsoon brought about by either furtheruplift of the northern parts of the Tibetan Plateau or increased global ice volumes[50], and the TarimBasin may have reached its present size ca. 500 kaBP following uplift of the Kunlun and TienshanMountains[51-52]. In northern Xinjiang, Northwest China, the earliest loess on the Tienshan occurredca. 800 kaBP, followed by further aridification and desert expansion ca. 650 and 500 kaBP[51]. InCentral Asia, and unlike in China, the increase in loess deposition was progressive, and increasedfrom ca. 7.8 cm/ka between 1.77 and 0. 85 MaBP to 12.1 cm/ka from 0.85 to 0.25 kaBP, when it roseto 20 cm/ka[24, 53]. This trend probably reflects the increasing aridity of this region, and the expansionof the Kara Kum and Kizyl Kum deserts. The expansion of these deserts across Central Asia andnorth China created a near-continuous barrier that limited the northward expansion of homininsfurther south and restricted west-east dispersal (or vice versa) except when there were “windows ofopportunity” in unusually mild interglacial periods.
One window that would have allowed hominins to enter North China from the west wasbetween 580 and 380 kaBP. The sediment record from Lake Baikal, Siberia, for example, showspredominantly interglacial conditions during this interval, with a weakened MIS12 and MIS14,and no indication of mountain glaciation[54]. Hao et al, suggested that “the extra-long NH (northernhemisphere) interglacial climate during MISs 15 – 13 provided favorable conditions for thesecond major dispersal episode of African hominins into Eurasia”[55]. Evidence for such a dispersalis provided by the hominin specimens from Dali and Jinnuishan, which several researchershave suggested should be classified as H. heidlebergensis[56-59]. It was probably through thisopportunistic type of dispersal that some hominins between 580 and 380 kaBP introducedAcheulean assemblages into China across the Movius Line[32, 60].
2.3 Dispersals and source-sink models
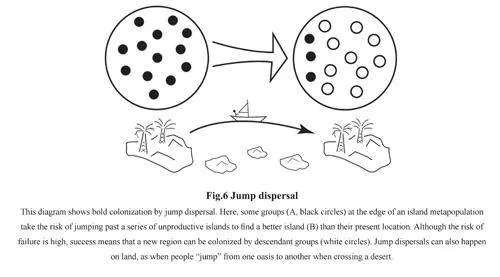
Population response within an ebb and flow model can be modelled in terms of refugia,source and sink populations. In the context ofhuman evolution, this type of model was firstapplied to Middle Pleistocene Europe[61, 62].In cold periods, hominin and warm-tolerantanimal species were confined to glacialrefugia in the Iberian and Italian peninsulas,and Southeast Europe which contained thesource populations of later expansion. Whenconditions improved, some groups movednorthwards and generated successive groupsthat expanded to the northern limit of thehominin range. These were sink populationsin that they depended upon recruitmentfrom groups further south to maintainviability(Fig.3). The strength of this modelwas that it provided an explanation for thevariability of the fossil hominins in Early andMiddle Pleistocene Europe: climatic changeprovided the driving force behind the repeateddispersal, fragmentation and recombinationof hominin groups, and also provided“windows of opportunity” in warm periods forimmigration from Southwest Asia. Martinon-Torres et al later applied this model to MiddlePleistocene China[21], which, like Europe, hasa variable fossil hominin record that does notshow a simple unilinear evolution. Louys andTurner also applied a source and sink modelto East Asia and suggested that Southeast Asiawas a potential refugium and source area forthe sink populations in North China[63].
2.4 Dispersals and naïve faunas
Another approach that might help explain east-west movements is to consider the interactionbetween hominins and their prey fauna. I previously suggested that one reason why H. erectusdispersed eastwards across Asia was that that although much of the Eurasian fauna would havebeen familiar to hominins, hominins would initially have been wholly unfamiliar to the fauna[64].Put another way, the Eurasian fauna would have been naïve as regards its awareness of the potentialdangers posed by a bipedal, tool-making, carnivorous ape (The same would also have applied to themain Japanese islands and the Philippines when these were colonised by H. sapiens).
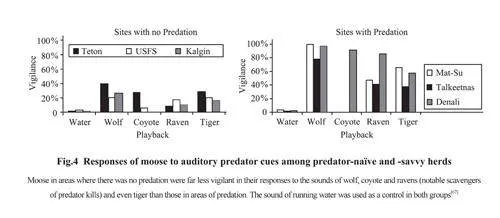
Naïve prey face a steep learning curve in survival when confronted with an invasivepredator such as wolf or humans. Although there are no regions left where the indigenous largemammalfauna is unaware of the potential or actual threat posed by humans, there are examplesfrom North America and northern Scandinavia of how recent faunas have reacted to the reintroductionof predators such as bear and wolf as conservation measures. Berger and colleaguestested “naïve faunas” for their vigilance to predator cues versus the responses in “predator-savvy”populations[65] (Fig.4). They found that predator-savvy moose were far more alert than naïve onesto the sound and scent of bears and wolves, and to raven calls (because these birds scavenge thecarcasses of moose that were killed by bears)[65]. The same researchers also showed that preyextinction is avoided because the prey populations quickly learn the importance of vigilance,and thereby becomes predator-savvy. Significantly, moose changed from being predatornaïveto -savvy in a single generation: they learnt to re-establish a “landscape of fear”[66].
Two points emerge from these findings. The first is that at the initial point of entry, Eurasianhominins had an advantage over their potential prey and carnivorous competitors because thenaïve indigenous fauna was unaware of their abilities; and second, the easiest way for groups ofH. erectus to maintain this advantage was to keep moving into new areas once the local fauna hadbecome predator-savvy to them. This may help explain how H. erectus managed to disperse overa very large area despite its very low population size. The proposed model here is a type of “waveof advance” once proposed for the rapid expansion of modern humans into North America[68],with its dramatic imagery of a “blitzkrieg” of humans targeting prey species that had no previousknowledge of the potentially lethal abilities of Homo sapiens. Here, the same ingredients areinvolved, but at much smaller scales of hominin group size and competence. No extinctionswould have followed: the advantages to the hominin predator would have been short-lived; and itwas unlikely any substantial increase in hominin populations occurred. Pott’s suggestion that thelow density of artefacts at Yuanmou, China, might represent “a relatively brief stop by homininsas they passed through the area” is consistent with the proposed scenario of hominins movingfrom areas where the prey had become vigilant to new areas where the prey was still naïve[69].
The second implication is that if, as suggested by, prey populations can relax or even losetheir vigilance in less than 10 generations, hominins could repeat their temporary advantageover potential prey by returning to areas where the prey had reverted from being predator-savvyto predator-naïve[67] (Fig.5). This way, hominins could still maintain viable local populationsproviding that they also practised a high degree of mobility. Because prey populations learn to bepredator-savvy in only a few years, the advantage to humans would probably not last more than afew hunting seasons, but this could still be important in the initial phase of expansion.
Loss of vigilance amongst prey populations may also have been a major factor affectingthe way hominins (and later, humans) recolonized regions that had been depopulated duringglacial periods. Repeatedly through the Pleistocene, especially across the northern limits of thehominin range, from north China through Central Asia to West and Northwest Europe, homininpopulations declined with each return to cold, dry (i.e., glacial) conditions, especially whenwinters were sub-freezing for prolonged periods. Crucially, however, in those areas wherehominins either declined in number or disappeared, much of the resident fauna would haveremained, or been replaced by, animals better adapted to the cold. Those animals that remainedwould have lost their vigilance regarding hominins, in the same way that moose in Swedenlost their vigilance against wolves when these became locally extinct (see above). Thus, whenhominins were able to move northwards again when the climate improved, they would haveencountered a naïve fauna, and thus recolonisation would have been easier – not just becauseof climate change, but because the prey was initially easier to kill. During the initial process ofcolonisation, they would have been at an advantage if they kept moving into areas where the preyfauna was naïve. A good example of where this process was likely repeated at the onset of eachinterstadial and interglacial is Northwest Europe, and the clearest and best documented exampleof this process is probably the region’s re-colonisation in the final part of the last glacial period,during which humans moved back into the depopulated areas of northern Europe and ultimatelycolonised Britain, Scandinavia and the Baltic region. The same process might have operatedwhen humans re-occupied the North China Plain after the LGM.
Predator competition might also have been a factor that at times inhibited dispersal. Turner forexample argued that hominins were largely excluded from Europe before the Middle Pleistocenebecause of competition with indigenous carnivores[70]. In East Asia, the giant hyaenid Pachycrocutabrevirostris may have impeded dispersal by H. erectus before it died out ca. 500 kaBP[71]. Itwas, however, primarily a scavenger rather than a direct threat as a rival predator[72, 73]. Becausehominins were repeatedly present on the Loess Plateau after 2.1 MaBP[28], and in the NihewanBasin after 1.66 MaBP[33, 35], it might not have been a major deterrent to hominins despiteopposing arguments by Binford and Stone and Boaz et al[74~76]. Additionally, if hominins were“savvy” and not naïve about Pachycrocuta, they might easily have learnt avoidance strategiesin the same way that moose learn to avoid wolves. By the Middle Pleistocene, homininshad probably emerged as the top predator in East Asia as well as in Europe and SW Asia[77].
2.5 Dispersal resulting from cultural adaptations
The remarkable range expansion of the genus Homo owes much to its ability to use know-howand material culture to extend its area of distribution. The ability to produce lots of sharp conchoidalflakes that could be used to strip meat from a carcass, cut wood, plants and skin probably underpinnedthe expansion of Homo erectus across Eurasia to an extent that would not otherwise have beenpossible for a small bipedal hominin. As argued above, its expansion across Eurasia would also havebeen facilitated by a prey fauna that was initially naïve. After 1.5 MaBP in West Asia (but much laterin East Asia), the development of bifacial and large cutting tools (LCT’s) would have further benefittedH. erectus. In the Middle Pleistocene, the presence of H. heidelbergensis at Jinnuishan, ShandongProvince, NE China, shows that this species had overcome many problems of over-wintering in harshclimates, and the development of wooden and later, stone-tipped spears, presumably made huntingsafer and more effective[78]. If the site of Kharganyn Gol 5, Mongolia, has Middle Palaeolithic artefactsin layers 6 and 7[79], Neanderthals or Denisovans were inhabiting an extremely harsh environment.In the last glacial cycle in European Russia, Neanderthals managed to extend their distribution upto 60°N. (roughly the latitude of Perm), where modern winters average -19°C, with 170 – 185 daysof snow per year. Those conditions probably marked the limits of their ecological tolerance[80]. (Thesite of Byzovaia (65°N, ca. 28 kaBP), which Slimak et al regarded as late Mousterian is UpperPalaeolithic[81, 82]). It is also possible that Neanderthals dispersed into Inner Mongolia, North China,between 50 and 40 kaBP if we assume that they made the Mousterian assemblages at Jinnsitai[83].If the mandible from Xiahe Cave is Denisovan and its dating to 160 kaBP is confirmed[84],Denisovans 160 kaBP were able to live in a very cold climate and at altitudes over 3500 m inaltitude, even if only seasonally. This is considerably higher than the Acheulean assemblage fromCona Cave, Georgia, which at 2100 m in altitude was the highest Lower Palaeolithic site in Asia[12].
2.6 Dispersals by Homo sapiens
Homo sapiens was qualitatively different from other hominins in its ability to colonise areas thathad never been inhabited by hominins, to colonise all regions already inhabited; and to eradicate allcompetitors. In those respects, humans have been incredibly successful as an invasive species.
Colonisation of new environments
East Asia shows better than any other area in the world how successful our species hasbeen in inventing ways of colonising new environments. Four new environments were colonised:oceanic islands, the Arctic, rainforests and the Tibetan Plateau above 4000 m in altitude. Eachcan be considered in turn.
Dispersals by sea travel
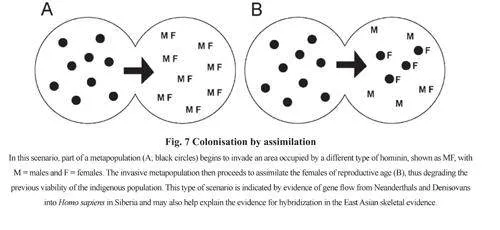
Colonisation by sea voyaging is a classic example of a “jump dispersal”, by which people(and also plants and animals) rapidly cross an area to reach a new place to settle (Fig.6). Thiscan be intentional or accidental. Intentional sea-faring requires the cognitive and technologicalabilities to build a sea-worthy boat or raft, provide it with food, fresh water and other provisions,and navigate it safely across open water. As seen below, there are numerous examples ofmaritime colonisation after 60 kaBP by H. sapiens that were almost certainly intentional.However, there are also claims that earlier hominins were also capable of intentional seavoyaging long before 60 kaBP. Morwood et al, for example, when discussing the discoveryof stone artefacts 800 kaBP on Flores, “conclude that H. erectus in this region was capableof repeated water crossings using watercraft”[85]. As the earliest evidence from Flores is nowdated to 1.0 MaBP[86], this evidence can also be seen as an even earlier example of intentionalsea voyaging by H. erectus. Bednarik asserted that “Navigation capability was apparentlyfirst developed between one million years and 800 kaBP in Southeast Asia” and described thebuilding of a raft with the type of simple tools found in Early Palaeolithic assemblages[87]. Acrew of five later sailed it from Timor to Australia. However, the fact that H. sapiens todaycan build a raft with simple stone tools and sail it to a known destination does not necessarilyimply that H. erectus had the cognitive and co-operative skills (or the impulse) to build a raftand sail into the unknown. On the other side of the world, the discovery on Crete of stoneartefacts dated to the last interglacial or even earlier and also Cyprus has led to speculationof sea-voyaging in the Mediterranean long before H. sapiens appeared in Europe[88~91].
An alternative view is that these discoveries are examples of inadvertent oceanic dispersal.This type of colonisation is surprisingly common over geological time, and is probably themechanism by which monkeys and other mammals reached South America from Africa[92], ormammals on Madagascar[93, 94]. Simpson termed this type of dispersal as a “sweep stake”, in thesense that only a few individuals were likely to be successful[95] 3). The key point here is that “thelonger a period of time in which a very rare event can occur, the more likely it is to do so”[96]. AsRuxton and Wilkinson point out regarding Flores and other islands in Wallacea “while accidentalarrival on an island would be merely improbable (for any particular island in any particular year)[97].But given the large number of islands, tsunamis and river flood events, such colonisation might bequite likely to happen to at least some islands over a time period of archaeological interest”.
The hominins that reached Flores, Indonesia 1 MaBP (and possibly several timesthereafter[98]), and Luzon, the Philippines 700 kaBP are probably examples of inadvertentdispersal[99], and show that over long periods of time, some individuals drifting by accident willreach land. These accidental dispersals may have resulted from hominins floating on mats ofvegetation detached from coastlines after a tsunami or cyclone[98, 100]. There are several recentexamples of humans being swept out to sea on vegetation. Ruxton and Wilkinson mention awoman rescued after clinging to driftwood for 80 km over six days after Hurricane Mitch in1998[97], one individual rescued after five days and another after nine days after the 1995 Pacifictsunami of 1995, and a pregnant woman who drifted 100 km in seven days on floating vegetationafter the 2002 Indonesian tsunami before rescue. As they note, “These anecdotal accounts clearlyillustrate the possibility of individuals being washed up on remote islands following tsunamisand similar events”[97]. To reiterate the point that even if 99% of such events do not result incolonisation of an off-shore island, over tens and even hundreds of thousands of years, a few will.As Ihara et al concede after their simulation study of unplanned marine dispersal “The resultsof our simulations indicate that the accidental colonization hypothesis has limited plausibility,although it cannot be excluded as invalid”[101].
Because hominins 100 – 200 kaBP may have been capable of building and sailing a seaworthyboat or raft, it is harder to dismiss the possibility of intentional sea travel with the evidence forhuman presence at Talepu on Sulawesi 100 – 200 ka and around the same time in Crete and possiblyCyprus[102]. If that was the case, we might expect further similar evidence from islands such as Malta,Corsica and Sardinia in the Mediterranean, or Timor and Lombok in Wallacea. Nevertheless, analternative scenario is that hominins were building boats 100 – 200 kaBP that were used close to theshore, but occasionally some were blown out to sea and a few inadvertently landed on an island. Atpresent, it is not possible to provide a definite answer to when intentional sea faring first began.
On current evidence, intentional sea voyaging appears unique to H. sapiens and resulted inEast Asia in the colonisation of Sahul – the conjoined landmass of Australia, New Guinea andTasmania – Wallacea – the islands between Java/Borneo and Sahul – the main Japanese islands, theRyuku Islands between Taiwan and southern Japan, the Philippines, the Kurile and Aleutian Islandsand possibly coastal North America. Sahul – the conjoined landmass of Australia, New Guineaand Tasmania – was reached by 50 – 55 kaBP[103] (and possibly as early as 65 kaBP if one acceptsthe dates and associated material from Madjedbebe rock shelter[104]). The fact that Australia wasreached by 50 – 55 kaBP means that the main colonisation of Wallacea had to begin by that time,although the oldest evidence from this region dates from 43,283 – 44,631 BP cal at Laili, Timor[105].The discrepancy may be an artefact of dating techniques, as the earliest dates from Wallacea arefrom 14C, but the earliest Australian dates were obtained from OSL. The extinction of H. floresiensisat Liang Bua, Flores, at ca. 50 kaBP may imply that H. sapiens arrived at that time. Fishing for tunaand other pelagic fish is evidenced at Jerimalai, Timor, by 42 kaBP[106].
The Japanese Islands were another region that could only have been colonised by seacrossings. The shortest and most direct route is across the Tsushima Strait, which was reducedfrom ca. 100 km width to ca. 20 km during the LGM[107], but colonists may also have arrived fromthe coasts of South China, Siberia, or via the Ryuku Islands[108]. The earliest site on PalaeoHonshu– the conjoined islands of Honshu, Kyushu and Shikoku – is Idemaruyama and is dated at ca.38 kaBP and its inhabitants used obsidian from the island of Kozushima, at least 45 km offshore[109]. Okinawa in the Ryuku Islands between Taiwan and Kyushu was colonised by 36 kaBP.It was probably reached by sailing from Taiwan because the powerful north-flowing KurishioCurrent would have made sailing south from Kyushu extremely difficult. Reaching Okinawa wasparticularly difficult because it involved crossing 105 km of open water between Taiwan and thenearest island of Yonaguni, and at least 220 km of open water between the southern and centralRyuku Islands. Interestingly, a reed boat was successfully paddled from Taiwan to Yonaguni in 45hours in 2019[110]. The evidence from Okinawa is also important in showing the world’s earliestexample of a species being translocated by humans: at the cave of Sakatari, layer 2, dated at 19635to 23425 BP cal, has evidence of pig (Sus scrofa), which was not native to the island but had tobe introduced[111, 112]. This translocation involved considerable foresight in allowing pigs to breedon the island and not eating them on arrival. It is not yet clear when Luzon in the Philippineswas colonised by our species but was probably after 52 kaBP, when H. luzonensis is recorded[113].
North America may also have been reached by sea along the “kelp highway” of the Kurileand Aleutian Islands that lie south and north of the Kamchatka Peninsula. Although Beringia isusually regarded as the most likely region from which humans reached North America, the richmarine environments along the coast of Northeast Siberia, and the proven ability of humansto undertake sea voyaging in East Asia by the end of the Pleistocene make a maritime route aplausible scenario for the colonisation of North America[114, 115].
There is so much that we do not yet know about these first sea voyagers - in particular,whether their boats were made from reed, skin, bark or wood; the communities (now submerged)that made and used them; their reliance upon marine resources for their subsistence; their sourcepopulations; and how some coastal communities in East Asia were able to grow sufficientlylarge to be able to create an emigrant population that colonized the Japanese islands and thePhilippines. Regarding Sahul, it was not colonised by accident as an unplanned one-way trip,but most likely as a result of planned migration involving numerous groups and numerous returnvoyages by several boat loads totalling perhaps 1000-2000 people for mainland Australia[116,117].In a recent simulation study, Bradshaw et al suggest a founding population of between 1300 and1550 individuals who could have arrived after one voyage, or (perhaps more likely) throughseveral voyages of ca. 130 people over 700 – 900 years[118]. And perhaps even more likely, bynumerous voyages of small groups (perhaps only a dozen or so) over several centuries. Theseestimates imply that the coastal regions of Sunda, mainland Southeast Asia and China hadcommunities that were large enough to send overseas substantial numbers of people, albeit mostlikely in small groups over several centuries. Sadly, we know nothing about them because theyexisted when sea levels were 40 – 60 m below present levels.
Dispersals into the Arctic
Humans were probably hunting mammoths inside the Arctic Circle before 40 kaBP, whensea levels were 50 – 80 m below present levels. At Sopochnaya Karga (the SK site), at 71° N, theskeleton of a mammoth, dated at ca. 40 – 45 kaBP, was found that showed injuries before deathand cut-marked bone[119, 120]. Two ribs had a notch-like lesion consistent with ones caused byspears, and post-mortem damage was also evident on the tip of one tusk and the mandible. Thereis also an assemblage from the site of Bunge-Toll 1885, where remains of woolly mammoth,woolly rhinoceros and bison were found, as well as a wolf humerus that had a puncture-cutwound that was likely caused by a human projectile. The humerus was dated to 4465+950-700 BP,which is close in age to a date of 47600 +2600-2000 BP for the mammoth remains[120]. Although it isuncertain whether these two sites denote rare events or were already part of a regular pattern ofmammoth hunting by humans in the Far North, they imply that humans 40 kaBP were huntingmuch further north than Neandertals appear to have managed. Conclusive evidence of mammothhunting comes from the site known as Yana RHS (rhino horn site) and the nearby Yana MassAccumulation of Mammoth (YMAM) site. These also lie at 71° N. and were used on at leastthree occasions between 27 and 29 kaBP[121, 122].
Although we have a basic outline of when humans first colonised the Arctic, there isstill much we need to understand. One issue is whether they had domestic dogs 30 kaBP, asargued by some[123-126], but doubted by others[127, 128]; another is how people moved over frozenlandscapes by skis, sleds, and snowshoes. We also need to learn more about the social networksthat would have been critically important in the harsh landscapes of the Arctic, and which mayhave been the means by which anthraxolite (a soft black rock) was present at Yana 600 km fromits source in the New Siberian Islands[129]. Ethnographic sources and experimental archaeologywould also lead to better understanding of how clothing was made, and fire was used in a largelytreeless landscape. One interesting clue is that the lack of wear on the toe bones of the 40 ka-oldTianyuandong individual may provide the earliest evidence for the habitual use of footwear[130].
Dispersals into rainforest
Homo sapiens was probably the first hominin to adapt to rainforests[131].
These were arguably the most challenging of the environments in East Asia that were colonisedfor the first time, Animals that were large enough to be worth hunting tended to be largely solitaryor lived in small groups (Tab.1); those living on the ground were often difficult to see and track, andpursuing them was impeded by dense vegetation, and those (such as monkeys) that lived in the highcanopy were even more difficult to hunt. With plants, it is necessary to know which can be eaten, andat what time of year, and whether it is the roots (as with tubers such as yams), stems or fruits that areedible; some are poisonous when picked and require processing by washing or boiling before they canbe eaten. Rainforests can also be unhealthy places to live because of water- or insect-borne diseasessince open wounds can easily fester and because much of the smaller fauna can be venomous.Although rainforests have been described as the world’s largest natural pharmacy, great skill andknowledge is required in knowing what plants (and which parts of them) can serve as medicines, andhow these can be used. Xhauflair et al observed that inhabitants of Palawan used 95 plant species[132],and quote Conklin who recorded over 1100 species of usable plants among the Huannóo of Mindoroin the Philippines. For humans to adapt to living in rainforests was thus a major achievement,particularly for a creature that originated and long flourished in grasslands and open woodlands.
The earliest clear evidence of humans in rainforest is from Lida Ajer, Sumatra, where two humanteeth were found in an assemblage largely consisting of orangutan, and dated at 65 – 73 kaBP[133].Evidence from Niah Cave, Borneo, shows that it was first occupied ca. 50 kaBP. The H. sapiens craniumis dated to ca. 45 – 39 kaBP cal. Humans were hunting bearded pig (29%), orangutan (6%) and leafmonkeys that are both terrestrial and arboreal (16%). There is evidence from starch grains of deeprootedyam (Dioscorea hispida and Dioscorea spp.) and sago palm (Caryota mitis or Eugeissona spp.)at 48 kaBP cal. These are particularly interesting because they need detoxifying before they are edible.According to Barker and colleagues[134], a piece of Dioscorea hispida the size of an apple can be fatalif eaten raw. Seeds of Pangium edule were also found; in ethnographic contexts, the hydrocyanic acidin these nuts could be removed by burying the ripe fruits or boiled seeds in a pit for 10 – 14 days andthen boiling them, or by burying the seeds with ash for up to 40 days[134]. In the cave, there are pitscontaining nut fragments dated at 39 and 35 ka that may have been used for detoxifying nuts.
There is also evidence of deliberate forest burning to create or enlarge open spaces that wouldprovide edible tubers and other plant foods, and for trapping those animals attracted to such patches.High frequencies of pollen of Justicia – a coloniser of burned areas – consistently occur in forestphases. This evidence is consistent with high frequencies of charcoal fragments in pollen coresfrom the Sulu Sea north of Borneo and dating back to 50 kaBP, and possibly an indicator of whenour species arrived in the region and began to modify their surroundings. According to Hunt andcolleagues study of the local vegetation at Niah, the vegetation was cyclic, and varied between lowlandand montane forest, and more open savannah-like environments indicating lower temperatures andrainfall compared with the present day[135, 136]. The interesting point here is that people may havereached the site ca. 50 ka during a climatic phase when habitats were fairly open, and biomassburning may have taken place during more forest-rich phases as a way of creating or enlargingopen spaces; people were not therefore necessarily foraging under closed canopy high forest[135].
Humans were also living wholly in rainforests in Sri Lanka before 40 kaBP. Excavationsof the caves of Batadomba lena, Fa-Hien lena and Kitulgala lena have produced remains of H.sapiens associated with a microlithic assemblage and an extensive bone industry that includedpoints that were probably used as projectiles or in snares as early as 36 to 38 kaBP[137~139].Faunal data indicates that semi-arboreal and arboreal primates comprise most of the mammalianassemblages; other resources exploited at Batadomba-lena include mouse deer, giant squirrel,mongoose, jungle cat and civet, as well as Canarium sp. nuts and starchy rainforest plants[139],all of which imply dedicated rainforest subsistence. Roberts and colleagues showed from stablecarbon and oxygen isotopic analysis of human and other animal bones from these caves thathuman diet was overwhelmingly from rainforest foods[140]. This lifestyle was maintained from38 kaBP through the rest of the Pleistocene (including the last glacial maximum) and into theHolocene up to when agriculture was introduced ca. 3000 BP. The rainforests of Sri Lanka weretherefore not only a resource zone but also a refugium during the last glacial maximum.
Xiaodong in Yunnan Province in southwest China has the earliest Hoabhinianassemblage – another link to southeast Asia – that is dated to 45 kaBP[141]. Because this sitelies in rainforest, this site may indicate the earliest colonisation of rainforest in mainlandsoutheast Asia. There is still much to learn from this site and others in Southeast Asiaand the Philippines about the range and type of plant and animal foods, and the type ofsocial networks that linked groups in this resource-rich but challenging environment.
Dispersal onto the Tibetan Plateau
The excavation of a blade assemblage at Nwya Devu shows that humans were living at4600 m in altitude as early as 30-40 kaBP[36, 37]. This discovery implies that by that time peoplewere able to cope with hypoxia – illness caused by a shortage of oxygen – as well as extremewinter cold. Huerta-Sánchez et al have suggested that this ability may have been derived fromDenisovan-like DNA[142]. As the Xiahe mandible, found at 3600 m in altitude on the north-eastpart of the Tibetan Plateau, is attributed to a Denisovan, the earliest inhabitants of the Plateaumay have been Denisovan who interbred with H. sapiens after 40 kaBP. Further a DNA analysisis needed to elucidate the demographic history of this region.
2.7 Dispersals, colonisation and population dynamics
The main issues here are the colonisation of abandoned environments and ones alreadyinhabited by other types of hominins.
Colonisation of abandoned environments
There are three examples from North China of humans colonising environments that hadbeen abandoned. On an ebb and flow model, these areas would have been abandoned because theirinhabitants moved south towards refugia in the Qinling Mountains when the climate deteriorated. Theresulting vacuum was filled by people adapted to cold conditions coming in from the north. The firstis evidence that Neandertals may have expanded ca. 47-42 kaBP eastwards into Inner Mongolia, ca.2000 km east of the previously recorded easternmost location in the Altai region of Siberia[83]. Thesecond immigration event is evident in the Shuidonggou (SDG) site cluster. The lower cultural layer ofSDG 1 contains an IUP (initial upper palaeolithic) blade-based technology produced by Levallois andprismatic methods of core reduction dated to ca. 43 kaBP and similar in age and composition to the IUPassemblages seen in the Altai Mountains and Mongolia[143]. The individual from Tianyuandong, dated at40328±816 BP cal may have been part of this immigration event[144].
The third immigration event into north China is evidenced by the late glacial microbladeindustry which probably originated in Siberia and appeared in north China at sites such asYoufang ca. 26-29 kaBP and Longwangshan ca. 27-25 kaBP cal and ultimately spread as farsouth as Lingjing in Central China[145~147].
Colonisation of inhabited environments
The study of hominin palaeo-demography in the eastern part of Asia is complicated by thefact that it was inhabited by several types of hominins in the Middle and Upper Pleistocene priorto the arrival of H. sapiens: in continental Asia, H. erectus, H. heidelbergensis, Neanderthalsand Denisovans; H. luzonensis in the Philippines, and H. floresiensis on Flores. By 50 kaBP,H. sapiens was the sole resident. We therefore need to consider the role of immigration, andpopulation assimilation and replacement.
Colonisation through assimilation of the indigenous population
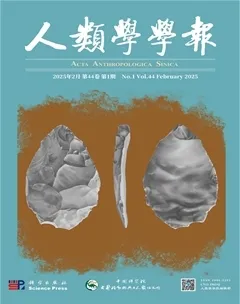
On some occasions, parts of a metapopulation began to invade an area that was occupied bya different type of hominin. The invasive metapopulation then proceeded to assimilate the femalesof reproductive age, thus degrading the previous viability of the indigenous population (Fig.7).This type of scenario is indicated by evidence of gene flow from Neandertals and Denisovansinto Homo sapiens outside Africa, and resulting in an “interbreeding bonanza”between Homosapiens, Neandertals and Denisovans[148] . Although we will never know the nature of theseencounters between H. sapiens and Neandertals, their long-term consequences may have beeneither negative (such as male hybrid infertility[149]), or positive if leading to “hybrid vigour”,enhanced immunity[150], and the acquisition of beneficial genes such as those that facilitated lifeat high altitudes such as the Tibetan Plateau[142] or in cold environments.
Colonisation through population replacement
Although it used to be widely thought that H. sapiens replaced all of its contemporaryhominins such as Neanderthals and Denisovans, total replacement without any interbreedingwas in fact rare. In East Asia, only two populations were completely replaced by the arrival ofH. sapiens at ca. 50 kaBP, and both were small and localised. The first was H. luzonensis in thePhilippines, and the second, H. floresiensis on Flores. Both were island populations that had beenisolated for a long time and were particularly vulnerable to an invasive predator. These homininswere small, probably had only a rudimentary technology, and above all, were naïve in that theyhad no prior experience of Homo sapiens. They were in the equivalent position of the dodo onMauritius that became extinct within a century after the arrival of humans. Their extinction wouldnot necessarily have been violent as it would have been sufficient for H. sapiens to appropriatetheir foraging grounds and displace them to marginal habitats.
Dispersals and social networks
Hominin (including human) dispersals can take place without a complex social network, aswhen a community relocates to a new area, or part of it forms a new or daughter community.
Rather than individual groups entering, learning about and using a new niche, a hominin thatwas socially complex and linguistically competent could operate as part of an integrated networkof groups, each of which could operate independently but maintain links that would allow sharingof information over a large area[151, 152]. Successful networks would operate with a high degree ofmutual reciprocity, so that risks could be pooled: groups would operate within a support networkso that individuals could rely on neighbouring groups if they encountered local adversity.
Networked dispersals are probably unique to H. sapiens and probably under-pinned themaritime colonisation of Wallacea, Australia, the Japanese and Ryuku Islands, the Philippinesand perhaps the Americas as these require the skills, materials, people and information of severalcommunities[64]. Effective networks would also have been essential when humans dispersed intoTransbaikalia, Mongolia and North China, where winters were harsh, and main prey species werehorse and gazelle that are migratory and have enormous annual territories of up to 30000 km2 [153-154].High mobility and high unpredictability of prey species and climate would have provided strongincentives for groups to operate as part of a social network as a way of minimising risk.
Evidence of palaeolithic networks is elusive, but a good example is the distribution of beadsfrom the Altai through Transbaikalia, Mongolia and North China in the IUP[155]. An important unifyingcharacteristic of this region is the use of ostrich egg shell beads and pendants made from animal teeth,as well as the occasional use of ochre. These first appear in Transbaikalia ca. 42–45 kaBP[156-158], andca. 40–42 kaBP in the Tolbor Valley of northern Mongolia[159-162]. The ornaments from Upper Cave,Zhoukoudian, are most like those from Denisova, and both assemblages include perforated canine teethof various types of deer and small carnivores, round beads, bone pendants, and perforated shells, noneof which are found in south China but are widespread across Mongolia and southern Siberia[155, 158]. Thehunting groups in this enormous region would have been part of an extensive network of groups thatoperated over an enormous territory in pursuit of highly mobile and often unpredictable migratory preysuch as horse and gazelle.
Other examples from East Asia are the long-distance exchange networks in obsidian in NEAsia (Japan, Korea, China, eastern Siberia) after 40 kaBP, with some obsidian transported 600 kmby sea between Hokkaido and Kamchatka, and up to 1000 km from Japan into Siberia[163, 164]; andanthraxolite at Yana that came from 600 km away[129].
3 Discussion: an outline hominin biogeography of East Asia
Hominin dispersals are the outcome of processes that are usually multi-facetted, and areonly rarely explained by a single model or variable. There are also profound differences in thetypes of dispersals undertaken by H. sapiens and earlier hominins. The main types of dispersalsthat occurred in East Asia in the Early and Middle Pleistocene are summarised in Fig.8 andFig.9. In the Palearctic Realm of continental Eurasia, climatic change was the main driver, withthe repeated alternation of cold and dry, and warm and moist periods of the Pleistocene. Withthe sparse data available for the Early and Middle Pleistocene, a simple ebb and flow model isan appropriate way of showing how hominins moved north when the climate permitted, andretreated south or become extinct at the limits of their range when the climate deteriorated. In thecase of China, the hominin skeletal record is sufficiently detailed to allow the identification ofpalaeodemes and a source-sink model offers greater insights into explaining that complexity.
Dispersals west to east (or vice versa) in the Palearctic Realm of East Asia can be modelledin several ways. The climatic changes that determined movements north to south would alsohave created corridors that permitted west-east movements if arid regions became grassland, orbarriers that prevented such movement if grasslands became desert. Hominin expansion acrosscontinental Asia can also be explained as simple range expansion by a “creep and crawl” modelof gradual dispersal, or by a more dynamic process in which hominins searched out prey faunasthat were initially naïve and easier to hunt.
For the Oriental Realm of South China and Southeast Asia, the two main factors thatunderpinned hominin dispersals in the Early and Middle Pleistocene were the repeatedfragmentation and recombination of rainforest that opened up or closed down corridors ofmore open vegetation, and changes in sea level that exposed or drowned land in coastalregions. These changes in sea level had far greater consequences in the continental parts ofthe Oriental Realm, particularly on the Sunda Shelf and the South China Sea than in the sub-Realm of Wallacea, where changes in sea level did not alter the basic geography of the region.As example, when sea levels were 50 metres below present levels, the islands of Borneo,Sumatra and Java were conjoined to each other and the Asian mainland, whereas the site ofJerimalai on Timor that is now within one kilometre of the coast was still usable as a basefor fishing 42 kaBP when sea levels were ca. 50 m lower[165]. Coastline bathymetry was alsoimportant. The post-glacial rise in sea level drowned 84% of the island of Palawan north ofBorneo[166], but hardly affected the coastlines ofmany Wallacean islands[165]. As Kealy et al pointout, the main consequence of rises and falls insea level in Wallacea were their effects on interislandvisibility[167]. Additionally, there is thefactor of oceanic drift that probably resulted inthe accidental dispersal of H. floresiensis and H.luzonensis (and their associated fauna) to Floresand Luzon respectively.
Dispersals in East Asia after ca. 70 kaBP byH. sapiens were very different, and were drivenby innovations that occurred more rapidly andwere more far-reaching than those of earlierhominins(Fig.10). The main innovation thatoccurred in both the Palearctic and OrientalRealms was the ability to colonise off-shoreislands such as those of Wallacea, PalaeoHonshu,the Philippines, the Ryukus islands and Sahul bya series of jump dispersals involving complex,inter-connected networks and probably hundredsof individuals. As Sahul was colonised by 50-55kaBP (and perhaps earlier), seafaring must havebegun earlier in Southeast Asia and Wallacea. Inthe Oriental Realm, humans were learning howto inhabit rainforest by 63-73 kaBP in Sumatra,and by 50 kaBP in Borneo and probably aroundthe same time in South China. In the PalearcticRealm, humans learnt how to inhabit regions ofextreme cold and thereby colonised the Arcticas well as eastern Siberia and Mongolia, andthe high Tibetan Plateau above 4000 m by 30 kaBP. This new-found ability to inhabit cold andharsh environments such as Mongolia also enabled humans to repeatedly move southwards intoNorth China and occupy a landscape that had been abandoned by communities whose sourcepopulations were further south in the valley basins of the Qingling Mountains and Yangtse Valley.
The other important issue involved in dispersals after 50 kaBP is the interaction between H. sapiensand its contemporaries. Genetic evidence indicates that assimilation was the main outcome of contactbetween humans, Neanderthals and Denisovans. Total replacement affected only the isolated islandpopulations of H. floresiensis and H. luzonensis on Flores and Luzon respectively.
4 Concluding statement
A focus on dispersals opens a wide range of doors through which we can investigatethe palaeolithic record of regions such as East Asia. A biogeographic perspective requiresconsideration of the prevailing climate, vegetation, relief and type of terrain, as well as theway the climatic changes of the Pleistocene would have created corridors for or barriers todispersal. Hominin dispersals can also be better understood if we assess the nature of their prey,and whether it was naïve or vigilant when encountering hominin hunters. The extraordinarydispersals by our species into rainforests, across open seas to off-shore islands, to the Arctic orthe highest parts of the Tibetan Plateau necessitate examining the types of technical, social andcognitive adaptations that made those dispersals possible. East Asia is particularly suitable forthese types of studies because of its diversity of landscapes and adaptations of its inhabitants.
Acknowledgement: I am grateful to the extensive and thoughtful comments of the reviewer whoread the first draft of this paper.
猜你喜欢
青少年科技博览(中学版)(2023年1期)2023-03-17 00:44:24
科学大众(2022年23期)2023-01-30 07:03:18
中国老年学杂志(2022年6期)2022-05-11 07:54:48
河北地质大学学报(2021年5期)2021-11-08 06:27:24
文艺生活·中旬刊(2018年5期)2018-09-26 09:51:08
董事会(2017年8期)2017-08-29 15:14:27
感悟(2017年4期)2017-05-02 20:34:23
企业技术开发·下旬刊(2016年9期)2016-11-23 03:02:08
科技资讯(2016年18期)2016-11-15 07:45:11
经济研究导刊(2015年21期)2015-11-03 12:23:45