
果园红壤供氮水平季节性变化及对长期套种绿肥的响应研究
2024-12-31 00:00:00钟珍梅李春燕邢世和翁伯琦
草地学报 2024年8期







摘要:本研究以柿子(Diospyros kaki)园为对象,研究了长期(近20年)套种豆科绿肥对土壤可溶性氮和氮水解酶季节性变化的影响。试验设3个处理:套种平托花生(Arachis pintoi)(TPA)、套种圆叶决明(Chamaecrista rotundifolia)(TPC)和清耕果园(CK)。结果表明:长期套种2种豆科绿肥没有改变果园红壤供氮水平的季节性变化规律,6月各处理硝态氮(NO-3-N)、可溶性总氮(Total soluble nitrogen,TSN)和可溶性有机氮(Soluble organic nitrogen,SON)含量最高,分别为17.71~21.81,31.40~75.12和7.77~47.65 mg·kg-1;12月NO-3-N,TSN和SON含量最低,分别为4.88~6.02,15.17~17.45和5.03~7.92 mg·kg-1。3月、6月和8月铵态氮(NH+4-N)含量高于4月、10月和12月;6月4种氮水解酶活性最高,8月和12月活性最低。长期套种2种豆科绿肥显著提高了生长季3—8月NO-3N,TSN,SON和NH+4-N含量以及蛋白酶、天冬酰胺酶、脲酶和谷氨酰胺酶活性。因此,长期套种圆叶决明和平托花生是通过增加生长季的可溶性氮含量和氮水化酶活性来增加土壤氮供应。
关键词:豆科绿肥套种;可溶性氮;季节性变化;氮水解酶;定位试验
中图分类号:Q556""" 文献标识码:A"""" 文章编号:1007-0435(2024)08-2514-09
Seasonal Variation of Nitrogen Supply Level in Soil of Orchard and Its
Response to Interplanting Green Manure
ZHONG Zhen-mei1,3, LI Chun-yan1,3, XING Shi-he2*, WENG Bo-qi1,3*
(1. Institute of Resources, Environment and Soil Fertilizer, Fujian Academy of Agricultural Sciences, Fuzhou, Fujian Province
350013, China; 2. College of Resource and Environmental Science, Fujian agriculture and forest university, Fuzhou, Fujian
Province 350002, China; 3. Fujian Engineering and Technology Research Center for Circular Agriculture, Fuzhou,
Fujian Province 350003, China)
Abstract:Legume mulch is an important approach to fix nitrogen from air and change nitrogen supply in soil. This study was conducted to determine the effects of 20-year legume mulch on seasonal changes of soil soluble nitrogen and N-hydration enzymes in persimmon orchard. Three treatments were Arachis pintoi mulch (TPA),Chamaecrista.rotundifolia mulch (TPC),and no vegetation mulch (CK). Our results showed C.rotundifolia and A. pintoi mulch had no significantly effect on seasonal changes of 4 types of nitrogen concentrations and N-hydration enzyme activities. Among different treatments,the highest concentrations of NO-3-N (from 17.71 to 21.81 mg·kg-1),TSN (from 31.40 to 75.12 mg·kg-1) and SON (from 7.77 to 47.65 mg·kg-1) were observed in June,and the minimum values with 4.88~6.02 mg·kg-1 NO-3-N,15.17~17.45 mg·kg-1 TSN and 5.03~7.92 mg·kg-1 SON were in December. NH+4-N concentration was higher in March,June and August than in April,October and December. Simultaneously,the highest activities of urease,protease,L-asparaginase and L-glutaminase were in June,and the minimum values were observed in August or December. However,C.rotundifolia and A. pintoi mulch significantly increased the concentration of soil NO-3-N,TSN,SON,NH+4-N,and the activities of protease,urease,L-asparaginase,urease and L-glutaminase in growing season. Our study indicated that the long-term C.rotundifolia and A. pintoi mulch increased soil nitrogen supply by adding soluble nitrogen concentrations and N-hydration enzyme activities in growing reason.
Key words:Legume interplanting;Soluble nitrogen;Seasonal variation;N-hydration enzymes;Long-term experiment
果园生草技术起源于欧美,在欧美、日本等国已实施多年[1],在防控杂草、防治和减少水土流失等方面效果明显[2-4]。Wei等[5]从中国知网和Web of Science数据库中检索2016年2月之前有关果园生草的论文,根据meta-analysis法设定的标准最后选定了国内73篇果园生草研究论文,涉及360个观测指标,结果表明果园生草增加了土壤可溶性有机碳(SOC)、总氮(TN)和有效氮、有效磷和总钾含量,在提高土壤肥力方面效果显著。套种豆科植物是果园生草的一种类型,其不仅能提高土壤有机质含量,提高土壤肥力和质量,还能通过豆科植物的生物固氮作用提高植物和土壤氮含量[6]。
目前,关于果园套种豆科绿肥对氮的影响研究主要集中在总氮和有效氮含量的变化方面。研究表明,豆科植物对土壤氮含量的影响大于非豆科植物[7-8],果园套种白三叶(Trifolium repens L.)[7,9]和皇冠野豌豆(Crown vetch)[7]能显著提高土壤总氮含量,果园套种绿肥会改变土壤硝态氮(NO-3-N)、铵态氮(NH+4-N)的含量[10-12],但对可溶性有机氮(SON)的影响研究较少。SON是土壤有机氮库中最活跃的组分之一,在土壤氮素循环过程和植物氮素营养供给中具有重要的作用[13-14],且与土壤供氮水平直接相关[15],因此需加强果园套种绿肥后SON含量变化的研究。当前绿肥套种的土壤肥力效应研究主要集中在比较不同模式方面,关于长期套种豆科绿肥之后果园红壤可溶性氮季节性变化的研究报道也较少。
土壤中的氮主要以有机氮的形式存在,在土壤酶及土壤微生物的作用下,大分子有机氮通过水解或矿化分解为小分子有机氮和无机氮,然后被植物或土壤微生物吸收利用[16-17]。因此,土壤酶活性被认为是土壤微生物功能发挥和土壤肥力的关键驱动因素。土壤脲酶、蛋白酶、天冬酰胺酶和谷氨酰胺酶等是土壤氮循环的重要水解酶,直接参与土壤含氮有机物的分解和转化,在土壤氮循环过程中发挥着重要的作用[18]。由于土壤脲酶、蛋白酶、天冬酰胺酶和谷氨酰胺酶活性强度可用来表示土壤氮素供应强度,研究这4种土壤酶活性的变化及与土壤肥力之间的关系,对揭示土壤肥力状况,评估土壤供氮能力具有重要的意义。
圆叶决明和平托花生均为多年生豆科绿肥,20世纪80年代引进中国后,在我国南方地区具有很强的适应性,被广泛应用于防治水土流失、提高土壤肥力、改善土壤理化性状等[19-21]。目前关于套种圆叶决明和平托花生对土壤肥力的影响研究大多属于短期研究[19-21],而长期套种这两种绿肥对果园红壤可溶性氮及氮水解酶的影响研究报道较少,且绿肥套种之后果园红壤SON及氮水解酶活性的季节性变化规律研究鲜见报道。本研究开展长期套种圆叶决明和平托花生后果园红壤SON及氮水解酶的季节性变化规律研究,旨在揭示豆科绿肥套种对土壤供氮水平的影响规律及作用机理。
1 材料与方法
1.1 研究区概述及试验设计
研究区位于福建省尤溪县玉池村,地理坐标26°25′N,117°57′E,属亚热带季风性湿润气候,年实有日照时数1 781.7 h,年降水量1600~1800 mm,年均温19.2℃,7月均温26.6~28.9℃,1月均温9.0~12.0℃,无霜期312 d以上;80%的降雨主要集中在2月至10月之间。试验地所处海拔150 m,坡向为东南方向,坡度15°;成土母质为第四纪红色粘土,土壤为红壤。试验区建立于1996年,至今已连续套种圆叶决明和平托花生20多年。试验设置3个处理,即(1)对照(CK):果园清耕;(2)TPC:果园套种圆叶决明;(3)TPA:果园套种平托花生,圆叶决明和平托花生环绕柿树(Diospyros kaki)主干周围直径20 cm以外种植。平托花生为多年生草本植物,不刈割,全年覆盖,无枯草层,圆叶决明在研究区为一年生,5月种子自发萌发,6月中下旬快速生长,到10月地上部结荚完成生育期,随后刈割地上部覆盖在果树周围的地表,使其自然腐解;清耕果园每年人工除草3~4次。每处理每棵果树施NPK复合肥[16∶16∶16] 0.35 kg、钙镁磷肥(FCMP) 0.25 kg,氯化钾(KCl) 0.5 kg。每个处理2个重复小区,每小区长25 m,宽4 m,面积100 m2,由8个梯台组成,每个梯台上种植有大小一致的1棵果树,共16棵果树。1995—2012年种植的果树为桃树,2012年至今种植的果树为柿树。
1.2 土壤样品采集
土壤样品采集时间为2017年的3月15日、4月15日、6月15日、8月15日、10月15日和12月15日。首先,将每个小区的8棵果树按从上到下的顺序编号为1~8,每小区选编号为2、4和6的为取样果树,即为3次重复。其次,以选取的果树树冠环形投影线为基准,沿基准线外30 cm环形线设置5~6个取样点进行取样,取样深度0~20 cm。将取得的土样混合均匀,再次将2个重复小区同编号的2棵果树采集的土壤按不同土层再次混匀,用四分法取500 g装入封口袋中封好,放入保鲜箱中带回实验室,测定土壤可溶性氮含量和氮水解酶活性(图1)。
1.3 研究方法
1.3.1 土壤温度和湿度的测定 在地表5 cm处埋设温度计,每隔5 d分别于早(8:00)、中(12:00)和晚(6:00)记录土壤温度,持续1年,取3次温度的平均值计为日均温,每月的日均温平均值即为月均温。土壤含水量采用烘干法测定,土壤鲜样称初始重,在60℃烘干至恒重再称重,2次称量之差即为土壤含水量,用百分比表示。
1.3.2 土壤可溶性氮测定 称取10.00 g鲜土过2 mm筛,置于100 mL离心管中,加入2 mol·L-1的KCl溶液50 mL,250 r·min-1震荡1 h后静置30 min,使用0.45 μm微孔滤膜过滤,制得待测母液。取待测母液5 mL用连续流动注射分析仪(Flowsys,Systea,意大利)测定土壤NO-3-N和NH+4-N含量。取待测母液5 mL稀释2倍,用TOC仪(TOC-L,CPH/CPN,Shimadzu,日本)测定土壤TSN含量。SON含量用以下公式计算SON=TSN—(NH+4-N)—(NO-3-N)。
1.3.3 土壤氮水解酶测定 参考Kandeler and Gerber(1988)的方法测定脲酶活性[22],土壤脲酶活性用经脲酶水解后每克土产生的铵态氮含量表示,单位为mg·g-1。参考Lowry(1951)的方法测定土壤蛋白酶活性[23],土壤蛋白酶活性用经蛋白酶水解每克土产生的络氨酸含量表示,单位为mg·g-1。参考Kanazawa amp; Kiyota(1995)的纳氏试剂法测定L-天冬酰胺酶和L-谷氨酰胺酶活性[24],土壤L-天冬酰胺酶和谷氨酰胺酶活性用经酶水解后每克土产生的铵态氮含量表示,单位为mg·g-1。
1.4 数据处理
数据采用Excel 2016整理,利用统计分析软件SPSS 12.0进行统计分析,处理间的差异显著性采用单因素方差分析法和Duncan新复极差法进行统计分析。数据用“平均值±标准差”表示,差异显著性水平Plt;0.05。
2 结果与分析
2.1 套种豆科绿肥对红壤温湿度的影响
研究区的年日均温如图2所示。CK、TPC和TPA处理月均温范围分别为15.58~32.83℃、16.50~31.33℃和16.33~29.00℃。1年当中3种处理最低月均温均出现在12月,最高月均温则出现在6月。6—10月是一年中的高温期,该时期TPA和TPC处理月均温低于CK,12—4月的月均温则高于CK,表明套种绿肥有利于低温和高温季节的温度调节。
不同月份3种处理下土壤含水量存在差异(图3)。雨季(3—6月)土壤含水量高于其他季节;6月和8月TPA和TPC处理的土壤含水量显著高于CK(P=0.02,0.001lt;0.05)。3月和12月,TPA处理土壤含水量也显著高于CK(P=0.03,0.002lt;0.05),但在TPC处理与CK之间差异不显著。4月和10月,3种处理之间没有显著差异。
2.2 双因素方差分析
双因素方差分析结果如表1所示。校正模型的F值和P值分别为122.153和0.000lt;0.01,表明线性模型具有统计学意义,且处理、月份及其2因素的交互作用对NO-3-N、TSN、SON、NH+4-N含量及脲酶、天冬酰胺酶和谷氨酰胺酶活性均有显著影响。处理和月份对蛋白酶活性有显著影响。
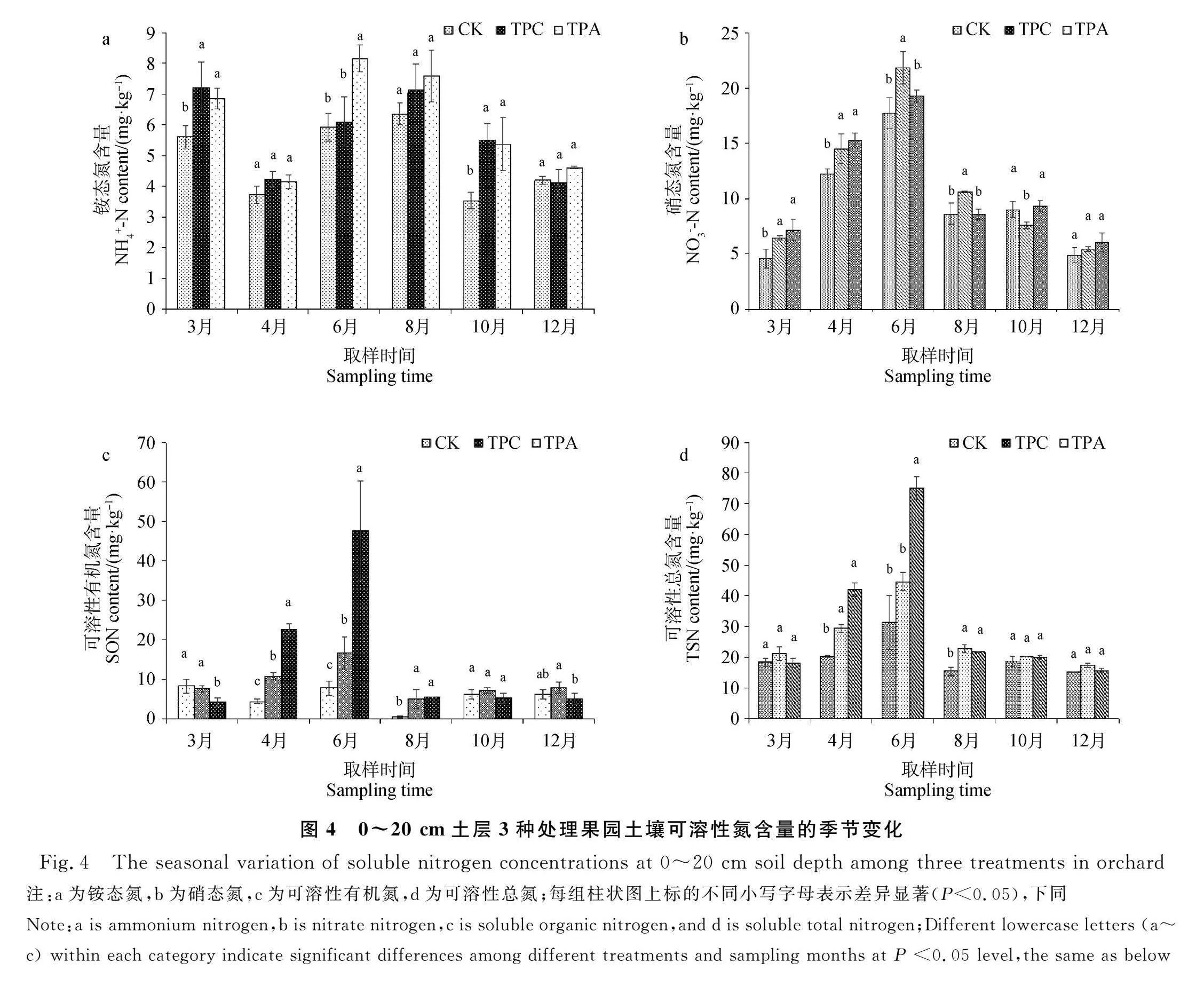
2.3 果园红壤可溶性氮含量的变化
3种处理的土壤可溶性氮呈相似的季节变化规律(表2)。NO-3-N,TSN和SON含量最高出现在平均温度和土壤含水量最高的6月,分别为19.61,50.38和24.05 mg·kg-1,NO-3-N和TSN含量最低值出现在平均温度最低的12月,分别为5.44和16.09 mg·kg-1,SON含量最低值在8月,为3.68 mg·kg-1。NH+4-N含量的季节分布不一致,3月、6月和8月NH+4-N含量最高,4月含量最低。套种绿肥圆叶决明和平托花生对土壤可溶性氮有显著影响(表2)。TPA处理NH+4-N,SON和TSN含量最高,分别为6.12,14.84和32.12 mg·kg-1,TPC处理NO-3-N含量最高,为11.06 mg·kg-1。与CK相比,TPC和TPA处理均显著提高土壤NH+4-N,NO-3-N,TSN和SON含量,且TPA处理TSN和SON含量高于TPC。
从图4可知,与CK相比,3月、4月、6月和8月,TPC处理NO-3-N和TSN含量显著增加了42.05%,18.52%,23.18%,22.91%和14.66%,45.15%,42.15%,47.45%;3月和10月TPC处理NH+4-N含量较CK显著增加了28.53%和55.92%,4月、6月和8月TPC处理SON含量较CK也显著增加了146.76%,115.16%和983.31%。与CK相比,3月和4月,TPA处理NO-3-N含量显著提高了57.41%和24.42%,而6月和8月则相反;4月、6月和8月,TPA处理TSN和SON含量显著提高了107.15%,139.24%,40.76%和421.15%,513.47%,1 096.52%。
2.4 果园红壤氮水解酶活性
3种处理的4种土壤氮水解酶活性在全年均表现出明显的季节规律(表3)。6月份脲酶、天冬酰胺酶、谷氨酰胺和蛋白酶的酶活性最高,分别为407.18,163.71,69.80和6.21 mg·g-1。12月脲酶和谷氨酰胺酶活性最低,分别为216.73和38.27 mg·g-1。3月天冬酰胺酶活性最低,为72.78 mg·g-1,8月蛋白酶活性最低,为3.69 mg·g-1。圆叶决明和平托花生套种均对4种酶的活性有显著影响(表3)。TPA处理酶活性最高,CK最低。TPA处理脲酶活性分别比CK和TPC显著提高17.85%和8.96%。与CK相比,TPA和TPC处理天冬酰胺酶、谷氨酰胺酶和蛋白酶活性均显著提高,且TPA和TPC处理之间谷氨酰胺酶活性也差异显著,TPA处理谷氨酰胺酶活性比TPC提高8.96%。
从图5可知,与CK相比,3—12月,TPA处理均显著提高了蛋白酶活性,3月、8月、10月和12月TPC处理蛋白酶活性也显著增加。3月、4月、6月和8月,TPA处理的脲酶活性显著高于CK,而TPC处理的脲酶活性仅在6月和8月显著高于CK。3—10月,TPA和TPC处理天冬酰胺酶活性较CK显著提高;3—8月,TPA和TPC处理谷氨酰胺酶活性较CK显著提高,TPA和TPC在3月和6月之间存在显著差异,TPA处理具有较高的天冬酰胺酶和谷氨酰胺酶活性。
2.5 相关性分析
各指标相关性分析结果表明,土壤温度和脲酶、天冬酰胺酶和蛋白酶活性以及NO-3-N,NH+4-N和TSN的含量显著正相关;土壤含水量与天冬酰胺酶和谷氨酰胺酶活性及NH+4-N和TSN含量显著正相关。氮水解酶活性与可溶性氮含量的相关性分析结果表明,脲酶和蛋白酶与NO-3-N,TSN和SON含量呈显著正相关;天冬酰胺酶和谷氨酰胺酶活性与4种可溶性氮均呈显著正相关。TSN与NO-3-N,NH+4-N和SON含量均呈显著正相关,SON与NO-3-N和NH+4-N含量也呈显著正相关。
3 讨论
本研究发现,果园红壤SON含量及4种氮水解酶变化具有明显的季节性。6月温度较高,是植物快速生长的季节,植物快速生长可能产生更多的根系分泌物,这可能是导致果园红壤SON含量最高的原因之一。8月是全年温度最高的月份,脲酶和蛋白酶活性急剧下降,表明高温可能抑制土壤酶活性,这可能是导致8月果园红壤SON含量最低的原因。而到12月至次年3月是全年的低温期,该时期4种氮水解酶活性和SON含量也较低。赵满兴等[25]研究表明,黄土丘陵区人工林SON含量随季节变化明显,6月最高,11月最低。周碧青等[26]研究发现,亚热带龙眼(Dimocarpus longan)园和枇杷(Eriobotrya japonica)园土壤SON的季节变化表现为夏、冬季高于春、秋季,表明土壤SON变化具有明显季节性。由于季节性变化涉及的生物因子和非生物因子复杂,仅套种绿肥这一项管理措施不足以显著影响其季节性变化趋势。圆叶决明和平托花生均是多年生豆科绿肥,其中平托花生地上部全年不枯萎,而圆叶决明引种到福建后生长具有明显的季节性[19-20],即每年5月下旬萌发生长,10月下旬地上部枯死,刈割后覆盖于地表。尽管2种豆科绿肥生长存在一定的季节性差异,但其与对照相比,土壤可溶性氮含量和氮水解酶活性变化仍表现出一致的季节性规律。套种绿肥显著增加了生长季果园红壤可溶性氮含量和氮水解酶活性,表明绿肥的生长可以改变土壤局部微环境,进而影响土壤供氮水平。
研究表明,豆科绿肥是一种具有多种生态功能的植物,在提高土壤营养物质含量尤其是氮营养方面具有显著效果[27]。本研究结果表明,长期套种平托花生,4—8月均显著提高了果园红壤SON含量,而套种圆叶决明则在3—8月期间显著提高果园红壤SON含量。Xing等[28]研究表明,土壤SON含量受到土壤有机质的输入影响较大,且受到土壤类型、植被覆盖、土壤管理措施及土壤理化性状等的影响[29]。4—8月为平托花生旺盛生长季节,植物根系凋落物及根系分泌物进入土壤,增加了土壤有机质输入,这可能是导致套种平托花生后土壤SON含量增加的主要原因;而圆叶决明的生长季节在5—10月,套种圆叶决明也提高了该时期0~20 cm土层SON含量,此外,豆科绿肥生长季根瘤菌的固氮作用也可能导致土壤氮含量的增加[21]。钟珍梅等[30]研究表明,低温条件下圆叶决明腐解需要4个月,圆叶决明11月刈割翻压,到次年3月,腐解进入氮释放阶段,因此3—4月SON含量显著提高。Dinesh等[31]研究发现,套种豆科绿肥蔓草虫豆[Cajanus scarabaeoides (L.)]、山珠豆(Centrosema pubescens)、毛蔓豆(Calopogonium mucunoides)、野葛[Pueraria lobata (Willd.) Ohwi]12年能显著提高土壤SON含量,这与本研究结果一致。同时也有学者[32]认为,当减少对土壤扰动时,能降低土壤的反硝化反应,提高土壤的SON含量。本研究中表层土壤扰动的减少可能也是增加SON含量的原因之一。此外,SON含量与植物根系分泌的有机物密切相关[27],在微生物作用下,植物根系分泌的有机物能通过土壤微生物作用转化为SON[27,33]。
本研究还发现,3—4月,套种平托花生提高了4种氮水解酶活性,套种圆叶决明则提高了天冬酰胺酶和谷氨酰胺酶活性,6—8月,套种2种豆科绿肥均提高了4种氮水解酶活性。土壤酶活性的变化反映了土壤微生物功能的变化[18],3—8月,套种2种绿肥后果园红壤4种可溶性氮含量也显著增加。3—8月是植物营养生长开始至旺盛生长的阶段,豆科绿肥生长会分泌根系分泌物,改变了土壤微生物生存的微环境,进而导致土壤氮水解酶活性增强[10,27],而水解酶活性的增加直接导致土壤溶解性氮含量增加。相关性分析也表明,脲酶和蛋白酶与NO-3-N,TSN和SON含量呈显著正相关,天冬酰胺酶和谷氨酰胺酶活性与4种可溶性氮均呈显著正相关,表明脲酶和蛋白酶参与了土壤NO-3-N,TSN和SON的形成过程,而天冬酰胺酶和谷氨酰胺酶则参与了4种溶解性氮的形成过程。
4 结论
长期套种圆叶决明和平托花生未改变土壤可溶性氮和氮水解酶的季节性变化规律,3种处理4种溶解性氮(NO-3-N,NH+4-N,TSN和SON)含量和4种氮水解酶的活性均在6月最高,12月至次年3月出现低谷。长期套种圆叶决明和平托花生在大部分月份显著提高了NO-3-N,TSN和SON含量和4种氮水解酶的活性,且套种平托花生的效果强于圆叶决明。土壤温度是影响土壤氮解酶活性和可溶性氮含量的主要因素,其次为土壤含水量。脲酶和蛋白酶与NO-3-N,TSN和SON含量呈显著正相关;天冬酰胺酶和谷氨酰胺酶活性与4种可溶性氮均显著正相关。
参考文献
[1] HARTWIG N L,AMMON H U. Cover crops and living mulches[J]. Weed Science,2002,50(6):688-699
[2] LORIN M,JEUFFROY M-H,BUTIER A,et al. Under sowing winter oil seed rape with frost-sensitive legume living mulches to improve weed control[J]. European Journal of Agronomy,2015,71:96-105
[3] SHI Z H,YAN F L,LI L,et al. Interrill erosion from disturbed and undisturbed samples in relation to topsoil aggregate stability in red soils from subtropical China[J]. Catena,2010,81(3):240-248
[4] MASSIMO P,PAOLO T,CERD A. Mulching practices for reducing soil water erosion:A review[J]. Earth-Science Reviews,2016,161:191-203
[5] WEI H,XIANG Y,LIU Y,et al. Effects of sod cultivation on soil nutrients in orchards across China:Ameta-analysis[J].Soil amp; Tillage Research,2017,169:16-24
[6] SWAROOP MEENA R,DAS A,SINGH YADAV G,et al.Legumes for soil health and sustainable management[M]. Berlin:Springer,2018:387-408
[7] QIAN X,GU J,PAN H,et al. Effects of living mulches on the soil nutrient contents,enzyme activities,and bacterial community diversities of apple orchard soils[J]. European Journal of Soil Biology,2015,70:23-30
[8] CONSTANTIN J,BEAUDOIN N,LAURENT F,et al. Cumulative effects of catch crops on nitrogen uptake,leaching and net mineralization[J]. Plant Soil,2011,341:137-154
[9] XU L,ZHOU P,HAN Q,et al. Spatial distribution of soil organic matter and nutrients in the pear orchard under clean and sod cultivation models[J]. Journal of Integrative Agriculture,2013,12(2):344-351
[10]张道勇,李会科,郭宏,等. 间作白三叶对苹果/白三叶复合系统土壤微生物量碳、氮及酶活性的影响[J]. 水土保持研究,2015,22(5):39-45
[11]毕明浩,梁斌,董静,等. 果园生草对氮素表层累积及径流损失的影响[J].水土保持学报,2017,31(3):102-105
[12]白龙,陈雪,王洲,等.苜蓿和老芒麦生草对苹果园土壤氮素矿化的影响[J].园艺学报,2015,42(12):2469-2477
[13]CANFIELD D E,GLAZER A N,FALKOWSKI P G. The evolution and future of earth’s nitrogen cycle[J]. Science,2003,330:192-196
[14]VAN K C,CLOUGH T,VAN GROENIGEN J W. Dissolved organic nitrogen:an overlooked Pathway of nitrogen loss from agricultural systems[J]. Journal of Environmental Quality,2009,38(2):393-401
[15]ZHANG Y L,XU W J,DUAN P P,et al. Evaluation and simulation of nitrogen mineralization of paddy soils in Mollisols area of Northeast China under waterlogged incubation[J]. PLoS One,2017,12(2):1-19
[16]BURNS R G,DEFOREST J L,MARXSEN,JRGEN,et al. Soil enzymes in a changing environment:Current knowledge and future directions[J]. Soil Biology amp; Biochemistry,2013,58:216-234
[17]SINSABAUGH R L. Phenol oxidase,peroxidase and organic matter dynamics of soil[J]. Soil Biology Biochemistry,2010,42:391-404
[18]CALDWELL B A. Enzyme activities as a component of soil biodiversity:a review[J]. Pedobiologia,2005,49:637-644
[19]翁伯琦,罗涛,应朝阳,等. 福建红壤区适生牧草种质筛选及其套种于山地果园的生态效应[J]. 热带作物学报,2004,25(2):95-101
[20]黄毅斌,应朝阳,郑仲登,等.生态牧草筛选及其在生态果园应用的研究[J]. 中国生态农业学报,2001,9(3):48-51
[21]叶花兰,李春燕,郑向丽,等. 豆科牧草圆叶决明的研究进展[J]. 中国农学通报,2008,24(11):467-470
[22]KANDELER E AND GERBER H. Short-term assay of soil urease activity using colorimetric determination of ammonium[J]. Biology and Fertility of Soils,1988,6:68-72
[23]LOWRY O H,ROSEBROUGH N J,FARR A L. et al.Protein measurement with the Folin phenol reagent[J]. The Journal of biological Chemistry,1951,193:265-215
[24]KANAZAWA S, KIYOTA H. Estimation of L-glutaminase and L-asparaginase activities in soils by the indophenol method[J]. Soil Science and Plant Nutrition,1995,41(2):305-311
[25]赵满兴,白二磊,刘慧,等. 黄土丘陵区人工林土壤可溶性氮组分季节变化[J]. 水土保持学报,2019,33(2):258-263
[26]周碧青,陈成榕,张黎明,等. 亚热带不同果园土壤可溶性有机氮季节动态及其影响因素[J]. 农业环境科学学报,2016,35(9):1735-1741
[27]曹卫东,包兴国,徐昌旭,等. 中国绿肥科研 60 年回顾与未来展望[J]. 植物营养与肥料学报,2017,23(6):1450-1461
[28]XING S H,ZHOU B Q,ZHANG L M, et al. Evaluating the mechanisms of the impacts of key factors on soil soluble organic nitrogen concentrations in subtropical mountain ecosystems[J]. Science of the Total Environment,2019,651:2187-2196
[29]杨庆,钟珍梅,翁伯琦.农业生态系统可溶性有机氮研究进展[J].福建农业学报,2015,30(12):1223-1230
[30]钟珍梅,游小凤,翁伯琦,等. 不同温度下圆叶决明降解过程红壤可溶性氮及氮水解酶动态变化研究[J]. 草地学报,2020,28(1):95-103
[31]DINESH R,SURYANARAYANA M A,GHOSHAL CHAUDHURI S,et al. Long-term effects of leguminous cover crops on biochemical and biological properties in the organic.and mineral layers of soils of a coconut plantation[J]. European Journal of Soil Biology,2006,42:147-157
[32]CONSTANTIN J,BEAUDOIN N,LAURENT F,et al. Cumulative effects of catch crops on nitrogen uptake,leaching and net mineralization[J]. Plant Soil,2011,341:137-154
[33]武姣娜,魏晓东,李霞,等. 植物氮素利用效率的研究进展[J]. 植物生理学报,2018,54(9):1401-1408
[34]赵路红,李昌珍,康迪,等. 黄土丘陵区退耕地土壤可溶性氮组分季节变化与水热关系[J]. 生态学报,2018,38(2):689-697
[35]彭正萍. 植物氮素吸收、运转和分配调控机制研究[J]. 河北农业大学学报,2019,42(2):1-5
(责任编辑 刘婷婷)
