
抚育对长柄双花木叶枝氮磷钾生态化学计量的影响
2024-12-31 00:00:00夏晨晨王纪杰童玮缪璐阳汪玥玲王国忠郑伟健姜年春
福建农业科技 2024年10期





摘要:长柄双花木Disanthus cercidifoliusvar.longipes为国家Ⅱ级保护野生植物,有关抚育间伐对长柄双花木地径、树高生长及叶枝氮(N)、磷(P)、钾(K)的生态化学计量特征的影响尚未被认识。以钱江源国家森林公园长柄双花木群落为对象,建立未抚育(对照)、上层抚育、下层抚育、全抚育等4处理3重复的定位样地,监测了该植物地径、树高的年生长量,采样分析了叶片、枝条的N、P、K含量,并计算其化学计量。结果表明:抚育促进了长柄双花木的生长,与对照相比,地径、树高年生长量显著增加了20.0%~40.0%、20.0%~90.0%(P<0.05);全抚育叶片P含量显著下降(P<0.05),而K含量显著增加(P<0.05);枝条N含量在全抚育后显著下降(P<0.05),K含量在上层抚育后也显著降低(P<0.05);下层抚育和全抚育后叶片N∶P、K∶P显著升高(P<0.05),叶片N∶K显著下降(P<0.05),上层抚育显著提高了枝条N∶P、N∶K(P<0.05)。叶片中共有10对元素及化学计量比间的相关性达显著或极显著水平(P<0.05或P<0.01),枝条中仅有N∶P与N∶K具有显著性相关(P<0.05)。综上,抚育促进了长柄双花木的生长,改变了N、P、K在叶片、枝条中的积累与分配。
关键词:抚育;长柄双花木;叶片;枝条;生态化学计量
中图分类号:S718.5文献标志码:A文章编号:0253−2301(2024)10−0057−06
DOI:10.13651/j.cnki.fjnykj.2024.10.009
夏晨晨,王纪杰,童玮,等.抚育对长柄双花木叶枝氮磷钾生态化学计量的影响[J].福建农业科技,2024,55(10):57−62.
Effect of Tending on the Ecological Stoichiometry of Nitrogen,Phosphorus and Potassium in theLeaves and Branches of Disanthus cercidifolius var.Longipes
XIA Chen-chen1,WANG Ji-jie2,TONG Wei3,MIAO Lu-yang1,WANG Yue-ling1,WANG Guo-zhong1,ZHENG Wei-jian1,JIANG Nian-chun1*
(1.Kaihua County Forest Farm,Kaihua,Zhejiang 324300,China;2.Sichuan Institute Forestry and GrasslandInvestigation and Planning,Chengdu,Sichuan 610081,China;3.College of Environmental and Resource Sciences,Zhejiang A&F University,Hangzhou,Zhejiang 311300,China)
Abstract:Disanthus cercidifolius var.longipes is a second-class protected wild plant in China.The effects of thinning and nurturing on the ground diameter,tree height growth,and the ecological stoichiometric characteristics of nitrogen(N),phosphorus(P)and potassium(K)in the leaves and branches of D.cercidifolius var.longipes have not been fully understood.By taking the community of D.cercidifolius var.longipes in Qianjiangyuan National Forest Park as the object of study,the four-treatment and three-replicate permanent plot was established,including no nurturing(control),upper nurturing,lower nurturing,and full nurturing.The annual growth of ground diameter and tree heightof the plant was monitored.The N,P,and K contents in the leaves and branches were sampled and analyzed,and theirstoichiometry was calculated.The results showed that nurturing promoted the growth of D.cercidifolius var.longipes.Compared with the control,the annual growth of ground diameter and tree height significantly increased by 20.0%−40.0%and 20.0%−90.0%(P<0.05).After full nurturing,the P content in the leaves significantly decreased(P<0.05),while the K content significantly increased(P<0.05).The N content in the branches significantly decreased after full nurturing(P<0.05),and the K content also significantly decreased after upper story nurturing(P<0.05).The N∶P and K∶P in the leaves of D.cercidifolius var.longipes plant significantly increased after lower and full nurturing(P<0.05),while the N∶K in the leaves significantly decreased(P<0.05).The upper story thinning significantly increased the N∶P and N∶K in the branches(P<0.05).There were 10 pairs of elements and stoichio-metric ratios in the leaves that showed significant or extremely significant level correlations(P<0.05 or P<0.01),and only N∶P and N∶K in the branches showed significant correlations(P<0.05).In summary,nurturing promoted the growth of D.cercidifolius var.longipes,and changed the accumulation and distribution of N,P,and K in leaves and branches.
Key words:Tending;Disanthus cercidifolius var.Longipes;Leaves;Branches;Ecological stoichiometry
氮(N)、磷(P)、钾(K)是植物生长的必需元素,不同植物叶片、枝条中这些营养元素含量及其化学计量比具有相对的稳定性,可以作为判断植物生长的限制性营养元素指标[1],已在珙桐Davida involucrate、梵净山冷杉Abies fanjingshanensis、天目铁木Ostrya rehderiana等珍稀濒危植物[2−3]中进行了相关的研究,促进了这些濒危物种保护。
长柄双花木Disanthus cercidifolius var.longipes为国家Ⅱ级重点保护野生植物[4],零星分布于浙江、江西、湖南、广东的极少部分地区,叶形叶色美丽,花型花色精致。对长柄双花木的研究主要集中于种群结构、群落特征及植物区系等[5−9],磷素是限制该植物生长的主要营养元素[10],长柄双花木对光照的需求特性表现为中性树种,上层林木砍伐后将有可能形成以该植物为优势种的灌丛群落[11]。研究发现清理母树临近植物的枝叶、去除林下灌木等措施,促进了百山祖冷杉幼苗和母树的生长,为百山祖冷杉有效保育提供了技术支持[12]。抚育是否能促进长柄双花木的生长?是否对该植物氮磷钾化学计量比也产生影响?基于以上问题,本研究在钱江源国家森林公园设置了上层抚育、下层抚育、全抚育等3种生境改良措施的长柄双花木样地,并以未抚育作为对照,定位监测长柄双花木的地径、树高生长,采样并分析不同抚育长柄双花叶片、枝条的氮、磷、钾含量,阐明不同抚育对该植物生长及叶枝N、P、K化学计量的影响,可为长柄双花木有效保护的生境管理提供基础和技术支持。
1材料与方法
1.1研究区概况
研究区位于浙江省开化县齐溪镇钱江源国家森林公园,分布中心地理位置为29°24′05″N,118°13′15″E。属亚热带季风气候,年平均气温16.3℃,极端最高气温41.3℃,极端最低气温-11.2℃,稳定通过10℃的天数为237.4 d,≥10℃积温5 125.4℃。无霜期250 d,年平均降水量1 909 mm,年平均相对湿度81%,年日照时数1 785.1 h[10]。土壤为发育于花岗岩的红壤,土壤pH 5.3,土壤有机碳26.1 g·kg−1,碱解氮、有效磷和速效钾含量分别为103.3、4.2、104.8 mg·kg−1。主要乔木层树种为杉木,年龄32 a,平均胸径20.5 cm,平均树高14.0 m。
1.2试验设计与采样
2022年6月,在长柄双花木分布区建立4种不同样地,分别为未抚育(对照)、上层抚育、下层抚育和全抚育,面积分别为200 m2,重复3次,不同样地的抚育措施见表1。每个样地中至少包含5丛自然生长的树龄一致的长柄双花木。
2022年9月,调查不同样地中5丛长柄双花木的地径和树高,并进行定株标识;2023年9月,对长柄双花木生长情况进行复测,以计算地径和树高年生长量;同时用修枝剪采集调查植株东、西、南、北4个方向一年生的枝条、叶片,带回实验室备用[13]。
1.3样品处理与测定
采回的样品在实验室中用去离子水清洗后于105℃杀青30 min,而后在80℃烘干至恒重,用高速粉碎机将样品粉碎过0.149 mm后备用。N含量采用碳氮元素分析仪测定;H2SO4-H2O2消煮,采用钼蓝比色-分光光度法测定P含量,火焰分光光度法测定K含量[14]。
2结果与分析
2.1抚育对长柄双花木生长的影响
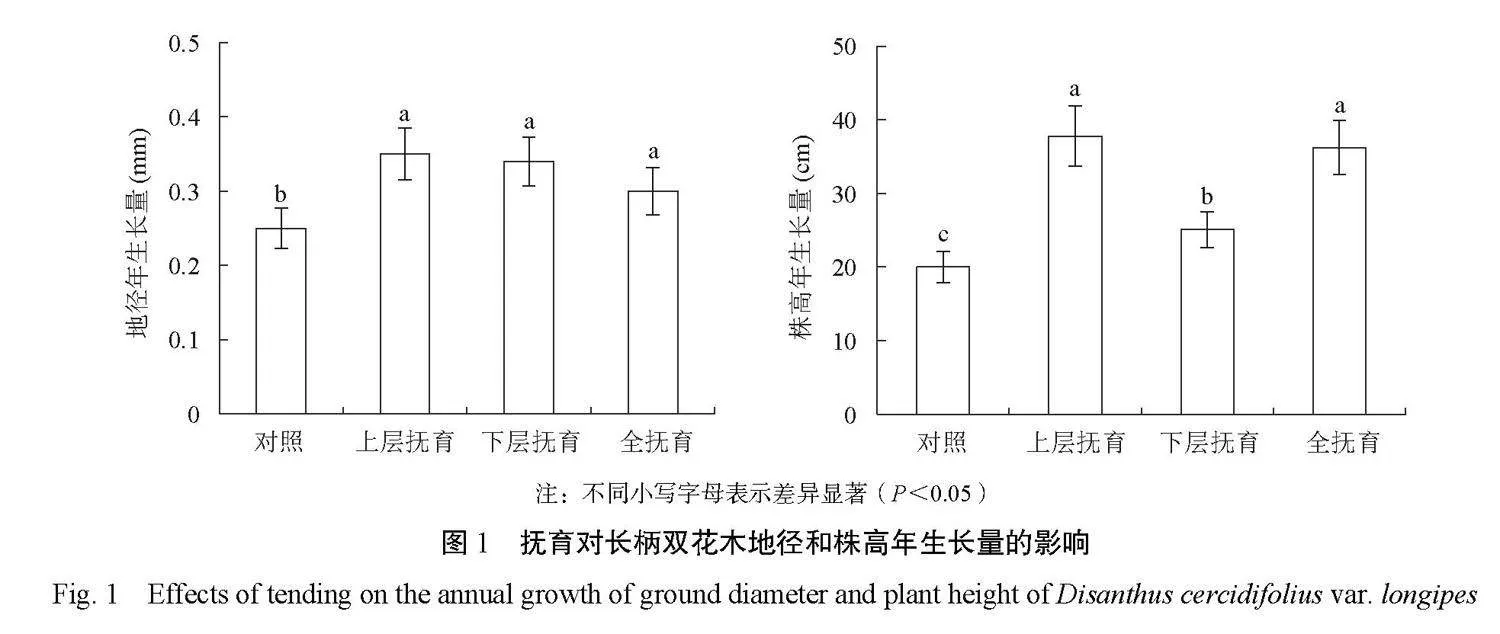
由图1可知,抚育促进了长柄双花木的生长,与对照相比,抚育后长柄双花木的地径年生长量显著增粗了20.0%~40.0%(P<0.05),株高年生长量显著增高了20.0%~90.0%(P<0.05)。
2.2抚育对长柄双花木叶枝氮磷钾含量的影响
2.2.1叶枝氮含量由图2可知,长柄双花木叶片N含量介于43.31~48.65 g·kg−1,不同抚育措施间没有显著性差异(P>0.05);枝条N含量介于17.99~24.66 g·kg−1,与对照相比,全抚育后枝条N含量显著下降了18.2%(P<0.05),也显著低于上层抚育和下层抚育(P<0.05)。
2.2.2叶枝磷含量由图3可知,长柄双花木叶片P含量介于0.56~0.78 g·kg−1,与对照相比,全抚育后叶片P含量显著下降了27.9%(P<0.05);枝条P含量介于0.47~0.55 g·kg−1,不同抚育措施间没有显著性差异(P>0.05)。
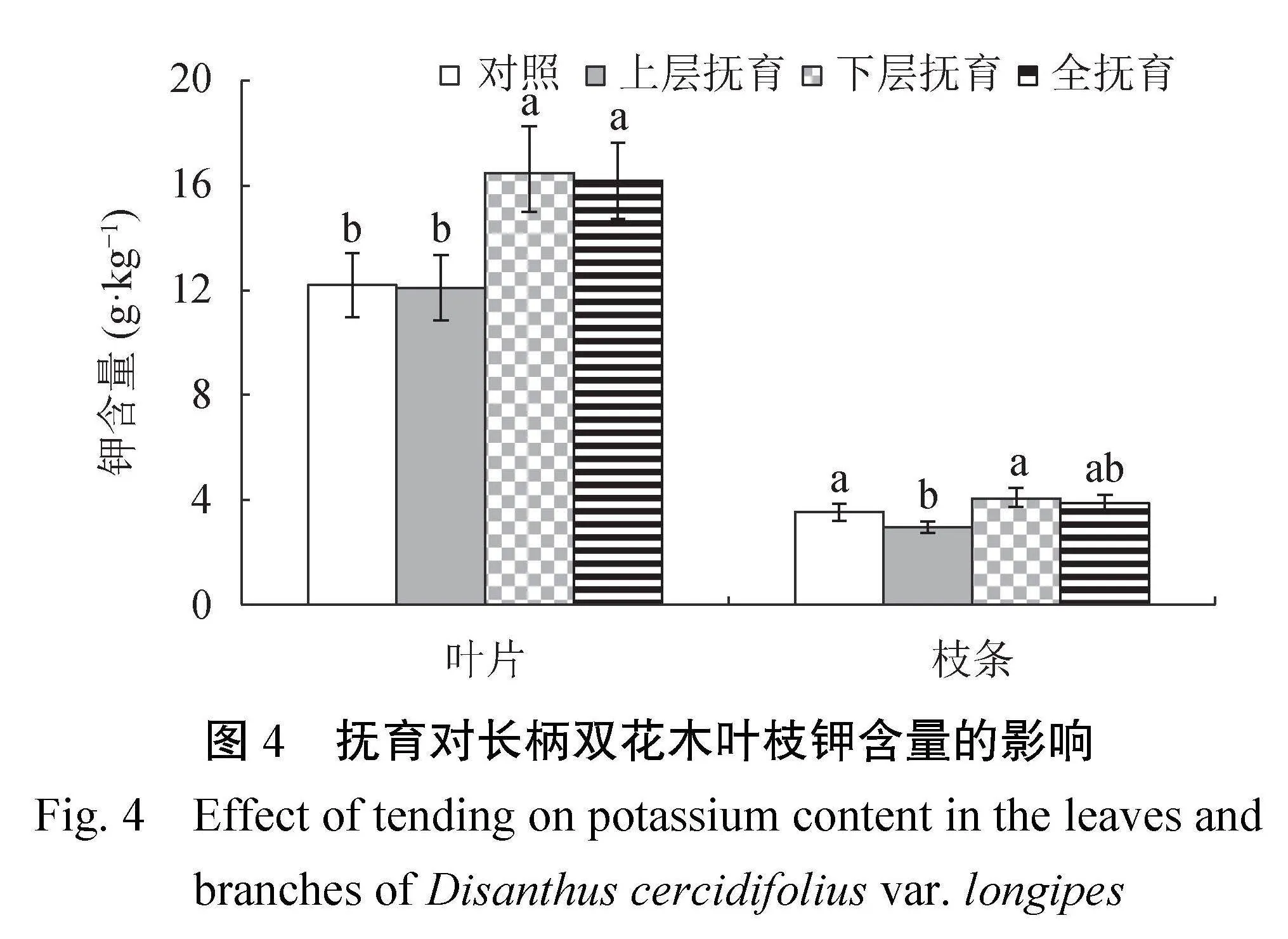
2.2.3叶枝钾含量由图4可知,长柄双花木叶片K含量介于12.09~16.47 g·kg−1,与对照相比,下层抚育、全抚育后叶片K含量显著提高了32.7%~35.1%(P<0.05),也显著高于上层抚育(P<0.05));枝条K含量介于2.95~4.05 g·kg−1,与对照相比,上层抚育后枝条K含量显著下降了16.2%(P<0.05),也显著低于下层抚育(P<0.05)。
2.3 抚育对长柄双花木叶枝氮磷钾化学计量比的影响
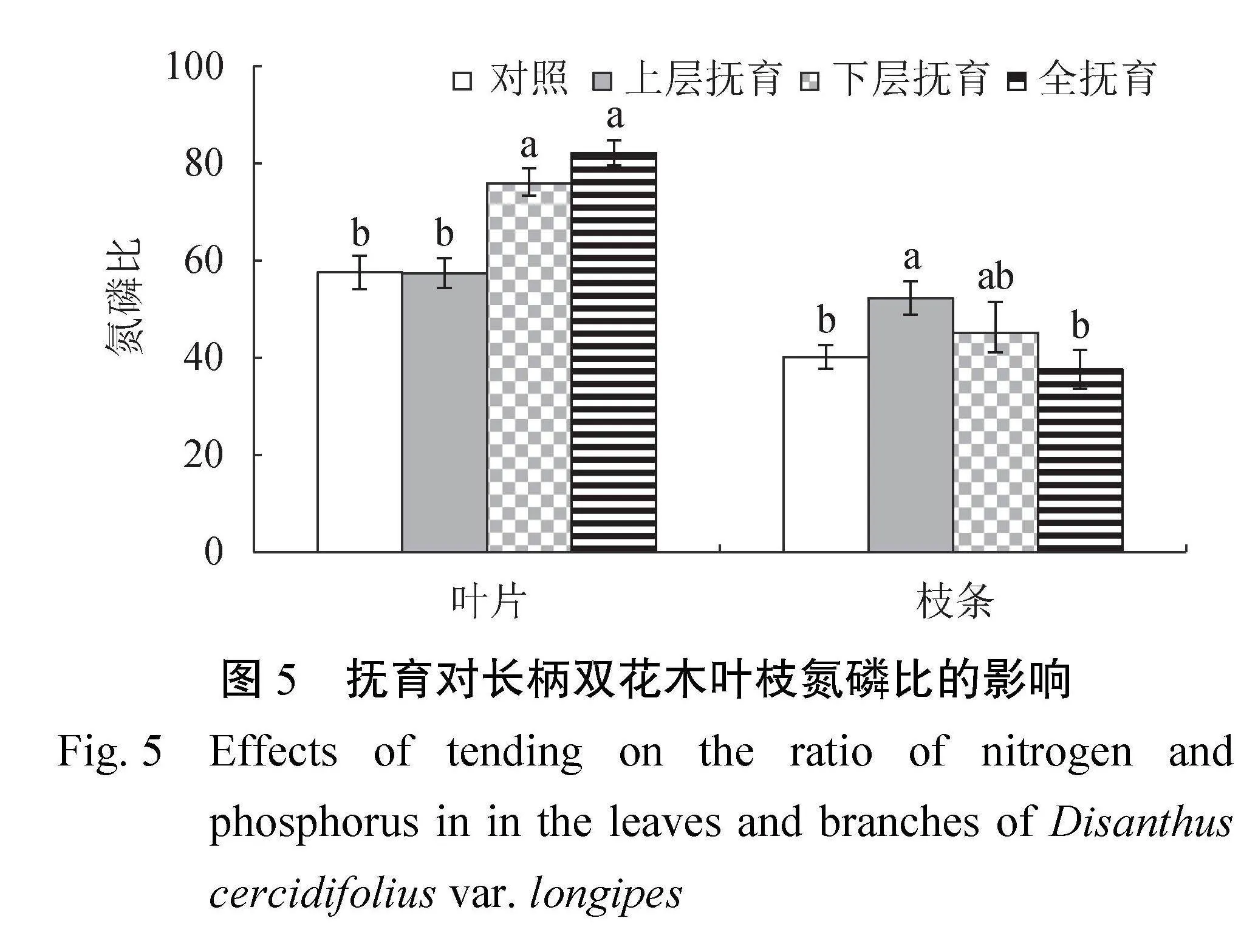
2.3.1 叶枝氮磷比 由图 5 可知,长柄双花木叶片N∶P介于57.45~82.22,与对照相比,下层抚育、全抚育后叶片N∶P显著提高了31.8%~42.8%(P<0.05),也显著高于上层抚育(P<0.05);枝条N∶P介于37.6~52.3,与对照相比,上层抚育后枝条N∶P显著提高30.2%(P<0.05),也显著高于全抚育(P<0.05)。
2.3.2叶枝氮钾比由图6可知,长柄双花木叶片N:K介于2.84~3.66,与对照相比,下层抚育、全抚育后叶片N∶K显著下降了19.4%~22.5%(P<0.05),也显著低于上层抚育(P<0.05);枝条N∶K介于4.67~8.35,与对照相比,上层抚育后枝条N∶K显著提高了33.8%(P<0.05),而全抚育后枝条N∶K显著下降了25.3%(P<0.05)。
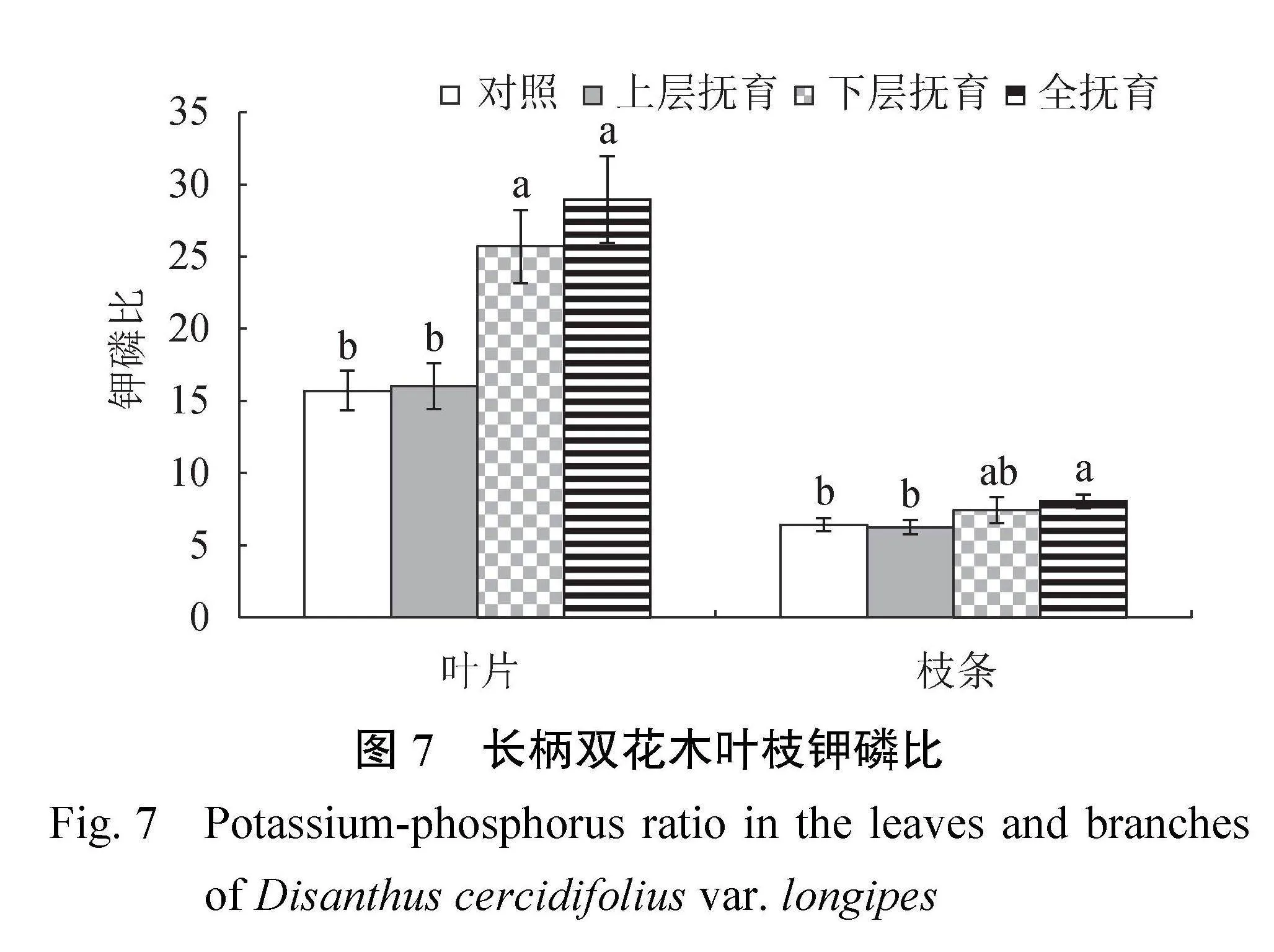
2.3.3叶枝钾磷比由图7可知,长柄双花木叶片K∶P介于15.72~28.94,与对照相比,下层抚育、全抚育后叶片K∶P显著上升了63.5%~84.1%(P<0.05),也显著高于上层抚育(P<0.05);枝条K∶P介于6.26~8.06,与对照相比,下层抚育后枝条K∶P显著提高25.3%(P<0.05),也显著高于上层抚育(P<0.05))。
2.4长柄双花木叶枝氮磷钾化学计量与生长的相关性
由表2可知,长柄双花木叶片P与K、N∶P、K∶P呈显著或极显著(P<0.05或P<0.01)负相关,与N∶K呈极显著正相关(P<0.01);叶片K与N∶P、K∶P呈极显著正相关(P<0.01),与N∶K呈极显著负相关(P<0.01);叶片N∶P与N∶K间呈极显著负相关(P<0.01),与K∶P呈极显著正相关(P<0.01);叶片N∶K与K∶P呈极显著负相关(P<0.01)。
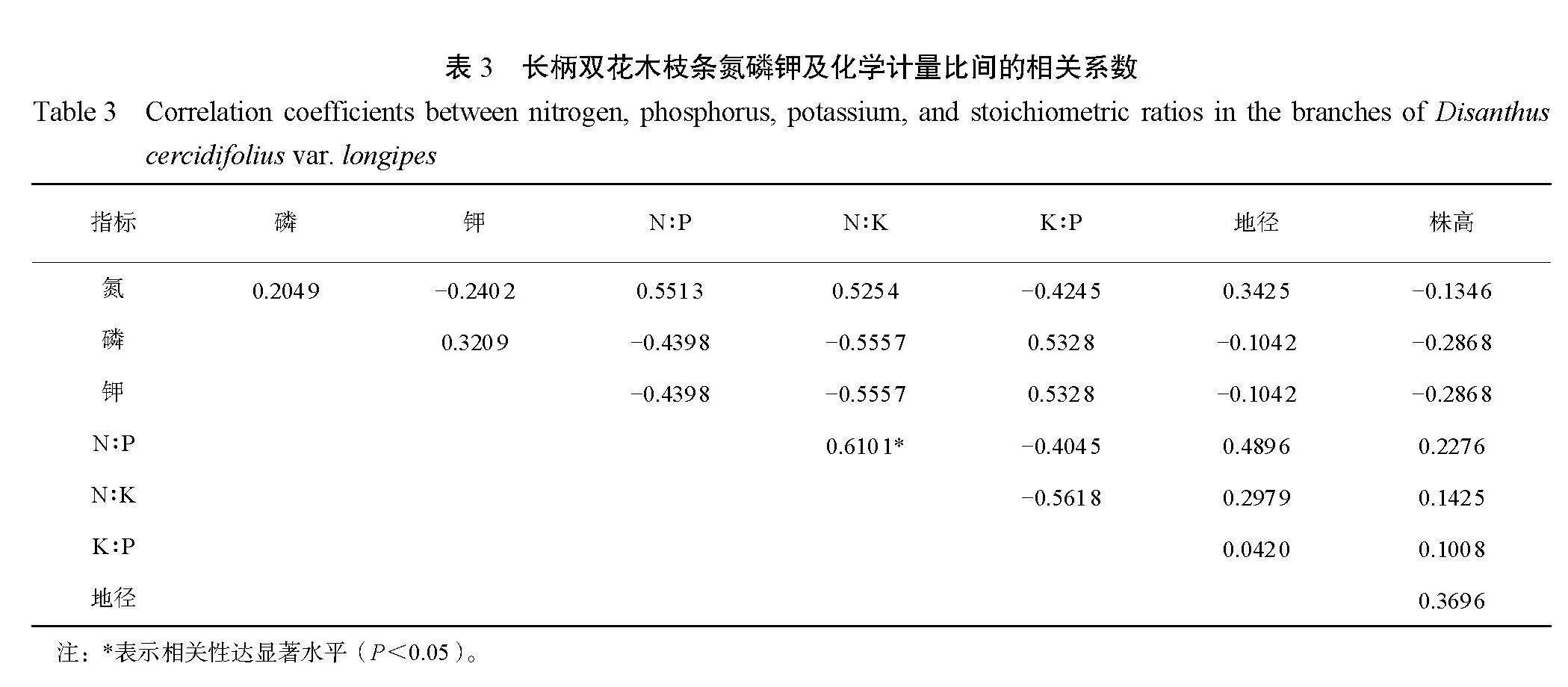
由表 3 可知,长柄双花木枝条 N∶P 与 N∶K 的 相关性达显著水平(P<0.05)。叶、枝氮磷钾含 量及化学计量比与地径、株高间的相关性均不显著 (P>0.05)。
3讨论与结论
抚育是适时适量伐除部分林木,调整林分密度,优化林分结构,促进林木生长的营林措施[15]。本研究表明抚育促进了长柄双花木的生长,地径、株高年生长量增加了20.0%~40.0%、20.0%~90.0%,这与前人研究结果相似[16]。
不同植物叶片N、P、K含量具有一定的内稳性,同时体现了不同植物对环境的适应性。本研究中长柄双花木叶片N、P、K平均含量为45.65、0.68、14.23 g·kg−1,其中N含量高于全国常绿木本植物叶片(14.71 g·kg−1)[17],主要原因是落叶树种叶片N含量高于常绿树种[18];P含量则低于全国常绿木本植物(0.96 g·kg−1)[18];K含量高于全国灌丛(8.38 g·kg−1)水平[19]。与前人研究结果相比,叶片N、K较高,而P含量则较低[10]。下层抚育和全抚育后的长柄双花木叶片P含量显著下降,而K含量显著增加,这与间伐后白桦叶片磷含量显著低于未间伐的研究结果一致[20]。全抚育后的枝条N含量显著降低,上层抚育后枝条K含量也显著下降,这与抚育提高了山杨枝条N含量的结果相反[20],这主要是不同树种、不同器官对氮磷需求及利用能力不同所引起的。
N、P、K三者之间的比值可以作为判断植物生长的限制性元素。当N∶P小于14或大于16时,植物生长的限制性元素分别是N素或P素[21],当N∶K>2.1、K∶P<3.4时,K是植物生长的限制性元素[22]。本研究中长柄双花木叶枝N∶P为37.6~82.2,N∶K为2.84~8.35,K∶P为6.26~28.94,表明P是限制该植物生长主要元素,这与前人研究结果一致[10]。下层抚育和全抚育后的长柄双花木叶片N∶P、K∶P显著升高,叶片N∶K显著下降,上层抚育显著提高了长柄双花木枝条N∶P、N∶K,这进一步说明抚育有利于N、K在叶枝中的积累,但并没有减轻磷素对长柄双花木生长的影响。
长柄双花木叶枝N、P、K及其化学量比与地径、树高生长的相关性分析表明,叶片中共有10对元素或化学计量比间的相关性达显著或极显著水平(P<0.05,0.01),而枝条中仅N∶P与N∶K的相关性达显著水平(P<0.05),化学元素及计量比与地径、树高生长间没有显著性相关(P>0.05)。
参考文献:
[1]贺金生,韩兴国.生态化学计量学:探索从个体到生态系统的统一化理论[J].植物生态学报,2010,34(1):2−6.
[2]崔明阳,张珍明,刘峰,等.梵净山冷杉和珙桐的碳氮磷分布特征[J].贵州农业科学,2016,44(2):48−51
[3]叶子豪,吴伟峰,彭健健,等.天目铁木和多脉铁木不同器官氮磷钾生态化学计量特征[J].浙江农林大学学报,2021,38(2):426−432
[4]陈涛梅,徐侠,周鑫洋,等.长柄双花木濒危机制研究进展及保护策略建议[J/OL].分子植物育种,2024:https://link.cnki.net/urlid/46.1068.S.20240328.1815.022.
[5]谢国文,谭巨清,曾宇鹏,等.国家重点保护物种长柄双花木南岭群落植物区系与资源[J].广东教育学院学报,2010,30(5):79−87.
[6]缪绅裕,曾庆昌,陈志明,等.南岭大东山长柄双花木群落物种组成与种群结构特征分析[J].植物资源与环境学报.,2014,23(1):51−57.
[7]王国兵,徐定兰,吴钦树,等.江西官山国家级自然保护区长柄双花木生长规律及种群年龄结构[J].南方林业科学,2017,45(4):8−12.
[8]耿梦娅,陈芳清,吕坤,等.长柄双花木叶性状异速生长关系随发育阶段和海拔梯度的变化[J].广西植物,2019,39(10):1387−1397.
[9]孟艺宏,徐刚标,卢孟柱,等.长柄双花木种群遗传结构及种群历史[J].林业科学,2020,56(7):55−62.
[10]周世水,沈汉,郑成忠,等.不同年龄长柄双花木枝叶氮磷钾生态化学计量特征[J].浙江林业科技,2022,42(2):27−31.
[11]李根有,陈征海,邱瑶德,等.浙江省长柄双花木数量分布与林学特性[J].浙江林学院学报,2002,19(1):20−23.
[12]吴友贵.拯救极度濒危植物全方位保护百山祖冷杉[J].浙江林业,2019(9):36−37.
[13]吴家森,蒋仲龙,吕爱华,等.不同年龄杨梅各器官氮、磷、钾化学计量特征[J].江西农业大学学报,2019,41(3):447−453.
[14]鲍士旦.土壤农化分析[M].北京:中国农业出版社,2000.
[15]国家质量监督检验检疫总局,中国国家标准化管理委员会.森林抚育规程:GB/T−15781[S].北京:中国标准出版社,2015.
[16]郝鑫海,牟长城,崔雅如,等.基于双哑变量模型预测透光抚育强度对次生林内红松生长的影响[J].应用生态学报,2024,35(6):1463−1473.
[17]任书杰,于贵瑞,陶波,等.中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究[J].环境科学,2007,28(12):2665−2673.
[18]HE J SH,FANG J Y,WANG ZH H,et al.Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China[J].Ecology,2006,149:115−122.
[19]秦海,李俊祥,高三平,等.中国660种陆生植物叶片8种元素含量特征[J].生态学报,2010,30(5):1247−1257.
[20]牛鉴祺,吕彦飞,王树力.抚育间伐对杨桦次生林非结构性碳水化合物质量分数和碳氮磷生态化学计量特征的影响[J].东北林业大学学报,2024,52(6):51−57.
[21]姜沛沛,曹扬,陈云明,王芳.不同林龄油松人工林植物、凋落物与土壤C、N、P化学计量特征[J].生态学报,2016,36(19):6188−6197.
[22]皮发剑,袁丛军,喻理飞,等.黔中天然次生林主要优势树种叶片生态化学计量特征[J].生态环境学报,2016,25(5):801−807.
(责任编辑:柯文辉)
