
红凉伞不同着生部位成熟果实内含物质与种子胎萌的关系
2024-12-27 00:00:00陈苗苗艾星梅赵财宝李翠萍孙媛媛
山东农业科学 2024年12期


摘要:为探明红凉伞成熟果实内含物质与种子胎萌的关系,本试验以同一枝条上的外侧胎萌果实与内侧非胎萌果实为材料,研究其形态以及外果皮、中果皮、内果皮、胚乳和胎萌种子胚根的可溶性蛋白、可溶性糖、淀粉含量和抗氧化酶、淀粉酶活性变化,并对这些指标进行相关性分析。结果表明,红凉伞种子胎萌与挂果时长相关,胎萌与非胎萌果实横纵径差异不大,但种子的百粒重、果实不同部位的内含物质均存在显著差异,且非胎萌果实高于胎萌果实,其中内果皮的含量相对较高;胎萌发生后,果实不同部位的可溶性蛋白、可溶性糖和淀粉含量以及a-淀粉酶、B-淀粉酶活性均显著降低,而在胚根中迅速积累,为胚胎的生长发育提供能量,但超氧化物歧化酶(SOD)和过氧化物酶(POD)活性在胚根中最低。相关性分析表明,对于胎萌果实,SOD活性与可溶性蛋白含量呈显著负相关,POD活性与可溶性糖、淀粉含量呈显著或极显著负相关,淀粉含量与a-淀粉酶、B-淀粉酶活性呈负相关,说明果实不同部位的可溶性蛋白、碳水化合物和这几种酶可能协同参与了胚发育的代谢过程。
关键词:红凉伞;种子胎萌;果实内含物质;成熟期;果实形态;生理特性
中图分类号:S686 文献标识号:A 文章编号:1001-4942(2024)12-0040-07
红凉伞(Ardisia crenata var.bicolor)为报春花科(Primulaceae)紫金牛属(Ardisia)植物,叶背、花梗及花瓣均带紫红色,是典型的双色叶树种,果实鲜红艳丽且数量较多,挂果期可达1年或更长时间,具有很高的观赏价值。在《中国植物志》(第五十八卷)中,把红凉伞作为朱砂根的变种来处理,两者性状存在着不同程度的重叠和过渡,导致两者缺乏明显的区分界限;本研究前期通过对花器官的解剖结构观察发现,两者在花芽分化上存在一定的差异性;刘雄伟等也对朱砂根及变种红凉伞的叶绿体全基因组结构进行了解析,探讨了物种间的系统发育关系,其结果可为报春花科植物的分类鉴定、系统进化及资源开发利用研究奠定基础。目前对红凉伞的研究主要集中在化学成分提取和药理活性分析等方面,结果表明:红凉伞根中含有三萜皂苷类、香豆索类、糖苷类及甾体类等多种生物活性化合物;魏江存等采用TLC法对红凉伞根茎中的主要成分进行鉴别,显示主要为岩白菜素。叶洪波等证实,岩白菜素类成分具有较好的抗炎活性。此外,红凉伞提取物可抑制乳腺癌细胞增殖、迁移和侵袭,促进癌细胞凋亡,对病毒转移具有重要的抑制作用,是著名的民族解毒草药。
以上看出,截至目前对红凉伞繁殖特性的研究较少。红凉伞果实成熟后不脱落,挂果期长,种子具有一种特殊的繁殖方式——胎生。收获时红凉伞种子具有较高的初始含水量,平均为52%左右,不能耐受成熟脱水,也没有休眠特性,会在母体上直接萌发,脱落后可快速成苗。通过前期形态观测发现,红凉伞同一侧枝上的成熟种子有些胎萌,有些非胎萌,据统计,同一个花序的胎萌率最高可达80%以上。但成熟果实不同部位的内含物质是否存在差异,是否与种子胎萌有关,目前这方面研究尚未见报道。为此,本试验以红凉伞胎萌和非胎萌果实为材料,通过测定两者果实不同部位碳水化合物、可溶性蛋白含量及抗氧化酶活性的差异,研究其与胎萌的关系,以期为进一步研究红凉伞种子的胎萌机理提供理论指导。
1材料与方法
1.1试验材料
红凉伞植株由实生苗繁育而来,均为4-5年生成株,植于西南林业大学苗圃基地(25°03′N,102°46′E)。该地属于亚热带高原季风气候,年平均温度15℃,年平均降水量约900 mm。2021年5月中旬,从植株底部相同侧枝上分别采集新鲜成熟且饱满无病虫害的胎萌(侧枝外侧)与非胎萌(侧枝内侧)果实,每株各采集20粒,采集10株,共400粒。将胎萌与非胎萌果实分别混合,立即带回实验室备用。
1.2试验方法
用天平分别称量胎萌和非胎萌果实鲜重,计算百粒重。然后,用自来水将果实冲洗干净后迅速擦干水分,分别剥离胎萌与非胎萌红凉伞果实的外果皮、中果皮、内果皮、胚乳及胎萌种子胚根,再分别称种子重量(去除果皮),并将不同部位样品用液氮速冻后放入-80℃冰箱中冷冻保存备用。
1.3测定指标及方法
红凉伞果实可溶性蛋白含量测定采用考马斯亮蓝法,可溶性糖、淀粉含量测定采用蒽酮比色法,超氧化物歧化酶(SOD)活性测定采用氮蓝四唑法,过氧化物酶(POD)活性测定采用愈创木酚法,a-淀粉酶和B-淀粉酶活性测定采用3,5-二硝基水杨酸法。以上指标测定均采用但方的方法,重复3次,取平均值。
1.4数据统计与分析
采用Microsoft Excel 2021对数据进行统计与作图,利用SPSS 21.0软件进行单因素方差分析和Duncan's多重比较(d=0.05)并进行相关性分析,图表中数据均用mean±SE表示。
2结果与分析
2.1红凉伞果实发育过程中的形态变化
对红凉伞果实发育过程中的形态观察可见:4月下旬花芽开始由外向内分化,6月中下旬盛花期结束后花朵开始脱落(图1A),此时子房开始膨大,果实呈绿色,9月初由绿色变成黄绿色(图1B),10月中下旬到11月初果实成熟,呈鲜红色(图1C);次年4月上旬果实表皮逐渐凸起,胚根伸长呈弯曲状并附着于果皮表面(图1D),5月中旬胎萌率逐渐增加,每个侧枝最高可达90%以上,同时非胎萌果实共同着生于同一侧枝上,剥离果皮后可见不同程度伸长的胚根(图1E),而种子由内种皮包裹。可以看出,红凉伞果实生长过程呈“慢-快-慢”的变化趋势,达到转色成熟后的果实大多数挂果半年以上才会发生胎萌。对胎萌和非胎萌果实进行纵切比较,结果(图1F)显示,两种果实的外果皮、海绵状中果皮、木质化内果皮、胚乳在形态和横径大小上无明显差异,但胎萌果实可见凸起的绿色胚轴,而非胎萌果实胚无明显变化。
从百粒重来看,红凉伞非胎萌果实和种子分别为24.3 g和14.2 g,均高于胎萌果实和种子(图2),但两者在果实百粒重上差异不显著,去除果皮后的胎萌种子百粒重显著低于非胎萌种子(Plt;0.05)。这说明胎萌的发生受果皮的影响较小,主要与果实着生部位和种子内含物质的积累有关。
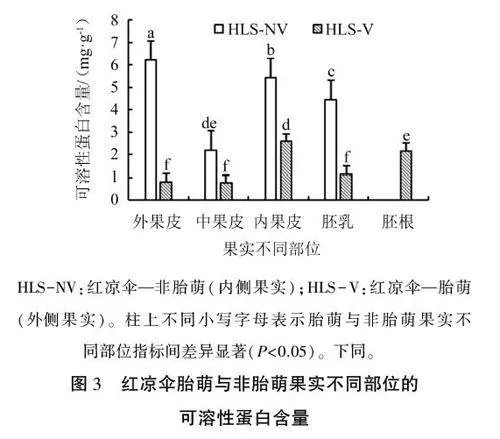
2.2红凉伞胎萌与非胎萌果实不同部位的可溶性蛋白含量差异
红凉伞果实不同部位的可溶性蛋白含量差异较大(图3),其中以非胎萌果实外果皮的可溶性蛋白含量最高,为6.21 mg·g-1,其次为内果皮和胚乳,中果皮的含量最低,且其各部位的可溶性蛋白含量均显著高于胎萌果实。种子胎萌发生后,内果皮和胚根中积累较多的可溶性蛋白,而其余部位的可溶性蛋白含量均迅速降低,以为胚胎的生长发育提供能量。
2.3红凉伞胎萌与非胎萌果实不同部位的可溶性糖和淀粉含量差异
由图4A可以看出,红凉伞非胎萌果实中果皮的可溶性糖含量最高,其次为外果皮和内果皮且两者差异不显著,胚乳的含量最低;胎萌发生后,果实外果皮、中果皮、内果皮的可溶性糖含量较非胎萌果实相同部位均显著下降,而胚根、胚乳的含量则显著上升,胚根的含量为其他部位的2.35-4.20倍。从非胎萌果实不同部位的淀粉含量(图4B)来看,内果皮的淀粉含量最高,其次为胚乳;胎萌发生后,内果皮和胚乳的淀粉含量迅速下降,而胚根的淀粉含量达到最高,说明胚根中的淀粉积累可能与内果皮、胚乳中的淀粉含量降低有关,其余部位的淀粉含量均处于较低水平,且差异不显著。
2.4红凉伞胎萌与非胎萌果实不同部位的抗氧化酶活性差异
由图5A可以看出,无论是胎萌还是非胎萌,红凉伞果实外果皮的SOD活性均最高,而中果皮、内果皮、胚根的SOD活性均很低,且两者间无显著差异;胎萌发生后胚乳的SOD活性迅速降低。由图5B可以看出,POD活性以非胎萌果实内果皮最高,表现为内果皮gt;外果皮gt;胚乳gt;中果皮;胎萌发生后,外果皮和内果皮的POD活性迅速降低,而中果皮和胚乳的POD活性均升高,但胚乳的POD活性升幅不大,外果皮、胚根的POD活性均显著低于其他部位。
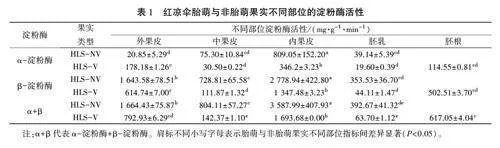
2.5红凉伞胎萌与非胎萌果实不同部位的淀粉酶活性差异
从表1中可以看出,相同侧枝上胎萌与非胎萌成熟果实不同部位的淀粉酶活性均存在较大的差异,其中,内果皮的a-淀粉酶、B-淀粉酶活性最高,均显著高于同一果实其他部位。果实不同部位的B-淀粉酶活性均大幅高于a-淀粉酶活性,且非胎萌果实a-淀粉酶+B-淀粉酶活性均高于胎萌果实,说明胎萌发生后,种子淀粉酶活性迅速降低,这可为胚胎发育提供能量,而胚根的淀粉酶活性处于较高水平,其中内果皮可能起主要作用。
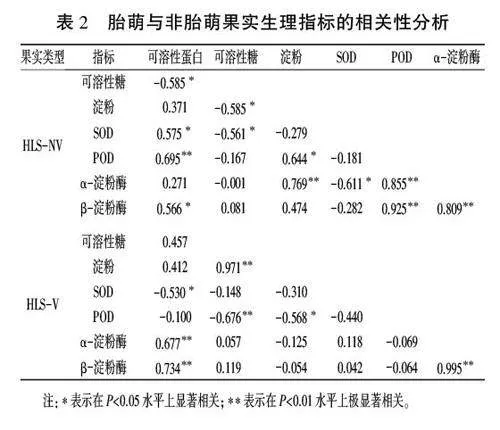
2.6红凉伞胎萌与非胎萌果实生理指标的相关性分析
对红凉伞胎萌和非胎萌果实碳水化合物含量、可溶性蛋白含量、酶活性各指标之间的相关性进行分析,结果如表2所示。其中,非胎萌果实的可溶性糖含量与可溶性蛋白含量、淀粉含量、SOD活性呈显著负相关,而可溶性蛋白含量与SOD、POD、B-淀粉酶活性呈显著或极显著正相关,淀粉含量与POD、a-淀粉酶活性呈显著或极显著正相关,a-淀粉酶、B-淀粉酶活性与SOD活性呈显著或不显著负相关,而与POD活性均呈极显著正相关。对于胎萌果实,可溶性糖含量与淀粉含量、可溶性蛋白含量与a-淀粉酶和B-淀粉酶活性均呈极显著正相关,SOD活性与可溶性蛋白含量、POD活性与可溶性糖和淀粉含量呈显著或极显著负相关,淀粉含量与a-淀粉酶、B-淀粉酶活性呈负相关,但未达显著水平。这说明胎萌的发生与果实不同部位生理特性的协同作用有关。
3讨论与结论
胎生植物包括种子胎生和营养体胎生,主要受自身的遗传特性和外界不良环境条件的影响。种子胎生对农作物来说是一种不利的性状,主要受高温、高湿、收获前过多的降水量、病虫害、营养条件和射线等外部环境的诱导,进而导致水稻、番茄、油菜等种子的产量和品质降低,造成较大的经济损失。但对于一些特殊植物而言,胎生具有兼性繁殖特性,且优势明显,胎生物种种子的发芽活力、抗逆性等均高于非胎生种子,且优良性状可遗传给后代,更易适应复杂多变的自然环境。红凉伞种子挂果期长达1年以上,过程中果实成熟至红色时形态上均无差异性,不能判断是否胎生,随着挂果时间延长至5个月后果皮表面逐渐凸起,胎萌即发生,半年左右可达到胎萌的高发期,但相同侧枝上的果实有些胎萌,有些非胎萌,且主要是从枝条的外侧向内、下部逐渐往上,与花芽分化的顺序一致,这可能与营养物质的优先分配和积累有关。胎萌和非胎萌果实百粒重、横纵径变化不大,但胎萌种子的百粒重迅速降低,说明胚轴的伸长主要与种子营养物质的消耗有关。因此,可以根据果皮颜色、果实转色时间、形状大小的变化等来判断种子胎萌的发生及难易程度。
生长于热带、亚热带的某些顽拗性种子通常具有胎萌现象,繁殖器官都有很高的含水量,如红树的胚胎、种皮、果皮含水量均高于54%,从母株上脱落后即可发芽,或在成熟过程中发生胎萌。研究表明,种子发生胎萌的前提条件是达到生理成熟,未成熟的种子即使是高度胎生的材料或通过一定的后熟时间,其种子也不会发生胎萌。在长期的环境适应过程中,胎萌种子除了形态结构发生相应的变化以外,逐渐进化出一套与其生境紧密契合的生理生态适应机制。种子内的碳水化合物可为胎生器官的萌发提供所需能量,其代谢水平的变化与种子萌发密切相关。研究表明,米勒魔芋珠芽萌发以后,蛋白质含量逐渐下降,黄瓜种子胎萌后的可溶性糖含量也逐渐降低,而胎生后的桐花树种子、百合珠芽等的淀粉含量呈不断积累的趋势,说明种子或营养体器官胎萌需要消耗大量的蛋白质和糖分,而淀粉代谢是影响珠芽形态发育的关键。本研究中,红凉伞种子胎萌后,果实不同部位可溶性蛋白、可溶性糖和淀粉含量大多呈下降趋势,而在胚根中迅速积累,说明对胚根的伸长生长具有重要的促进作用。
此外,抗氧化酶和淀粉酶活性的变化也是影响种子萌发的重要因素。研究表明,胎萌春大豆种子中的过氧化物酶活性比不易胎萌的种子高,而高枝假木贼种子的POD、SOD和CAT等酶活性在萌发不同阶段的协同作用均与胎萌有关,为种子在逆境中胎萌并正常生长提供保护。种子中的淀粉合成和分解受多种酶类物质的影响与调控,水稻成熟后期易于在穗上发芽的种子其a-淀粉酶活性明显高于不易在穗上发芽的种子,表明a-淀粉酶可能对后期胚的发育具有一定的调节作用。Neill等在研究玉米种子时发现,胎萌突变型种子的a-淀粉酶活性在胚胎发育后期显著增加,而野生型玉米种子的a-淀粉酶活性较低或下降,说明a-淀粉酶是在种子发育过程中合成的。大豆种子几乎不含a-淀粉酶,但B-淀粉酶含量很高,始熟期后,易胎萌品种B-淀粉酶活性呈下降趋势,而不易胎萌品种则急剧上升,且易胎萌品种B-淀粉酶活性总是低于不易胎萌品种。这虽不能证明B-淀粉酶具有谷类中a-淀粉酶的功能,但至少说明其作为一种贮藏蛋白,参与萌发和发育过程中的合成和分解。本研究中,非胎萌果实外果皮、胚乳的SOD活性和外果皮、内果皮的POD活性均显著高于胎萌果实,种子胎萌后胚根的酶活性最低:而内果皮的a-淀粉酶、B-淀粉酶活性均显著高于同一果实其他部位,且非胎生果实高于胎生果实。
本研究相关性分析表明,胎萌果实中SOD活性与可溶性蛋白含量呈显著负相关,POD活性与可溶性糖、淀粉含量呈显著或极显著负相关,淀粉含量与a-淀粉酶、B-淀粉酶活性呈负相关,说明胎萌发生后,包括淀粉酶在内的酶活性迅速降低,为胚胎提供能量。
综上看出,红凉伞果实胎萌与挂果时长、着生部位以及成熟度有关,果实不同部位的可溶性蛋白、碳水化合物协同参与胚发育的代谢过程,尤其是内果皮,它可能具有重要的调控作用。但造成这种生理上的差异性是受遗传还是环境条件的影响尚不明确。随着多基因组学的快速发展,研究发现种子萌发主要与植物激素信号传导通路中的合成和受体基因有关,因此,需进一步从植物激素调控以及胎萌相关基因表达等方面深入研究,全面探讨胎萌发育的遗传机制。
猜你喜欢
房地产导刊(2020年9期)2020-10-28 08:37:24
中国果树(2020年2期)2020-07-25 02:14:18
中国计算机报(2017年47期)2018-01-12 16:58:56
江苏农业科学(2016年8期)2017-02-15 19:40:24
江苏农业科学(2016年8期)2017-02-15 19:39:47
湖北农业科学(2016年20期)2017-02-15 17:39:53
山东农业科学(2016年12期)2017-01-21 14:37:22
现代园艺(2016年9期)2016-07-16 10:07:06
现代园艺(2016年2期)2016-03-15 16:05:02
浙江柑橘(2016年4期)2016-03-11 20:13:01
