
基于Maxent模型的青藏高原优势种草分布格局模拟研究
2024-12-20 00:00:00聂学敏李佳慧
安徽农业科学 2024年23期









摘要 以青藏高原嵩草、苔草、针茅3种优势种草为研究对象,通过植物标本库查询得到草地标本点位数据,选取气候、土壤质地、地形为环境因子,运用Maxent模型模拟出每种优势种草的分布格局,并运用刀切法筛选出主导环境因子,探讨主导环境因子对不同优势种草分布的影响与内在机制。结果表明,最冷季降水量、温度季节性变化标准差、年降水量、海拔是嵩草分布的主导环境因子(累计贡献率>80%);最冷季降水量、最暖季均温、年降水量、海拔对苔草分布贡献较大(累计贡献率82.55%);最暖季降水量、年均温、坡度、海拔对针茅分布影响较大(累计贡献率73.47%)。土壤质地对3种草分布影响很小(累计贡献率<5%)。总体上,生长季水热综合条件是制约青藏高原优势物种分布的最主要气候因子。
关键词 优势种草;Maxent模型;分布格局;环境因子;青藏高原
中图分类号 X 173 文献标识码 A 文章编号 0517-6611(2024)23-0069-06
doi:10.3969/j.issn.0517-6611.2024.23.016
Simulation of Distribution Pattern of Dominant Grass Species in Tibetan Plateau Based on the Maxent Model
NIE Xue-min1, LI Jia-hui2
(1. Qinghai Eco-Environment Monitoring Center,Xining,Qinghai 810007;2.School of Architecture and Urban Planning,Chongqing Jiaotong University,Chongqing 400074)
Abstract Taking Kobresia, Carex and Stipa as the research objects. The grassland specimen point data were obtained by querying the herbology database and the climate, soil texture and topography were selected as environmental factors to explore the influence and internal mechanism of the dominant environmental factors on the distribution of different grasslands, the Maxent model was used to simulate the distribution pattern of each dominant grass and the Jackknife method was used to screen out the dominant environmental factors. The results showed that the precipitation in the coldest season, the standard deviation of the seasonal variation of temperature, the average annual precipitation and the altitude were the dominant environmental factors for the distribution of Kobresia (accumulated contribution rate > 80%);the precipitation in the coldest season, the average temperature in the warmest season, the average annual precipitation, and the altitude had a greater contribution to the distribution of Carex (accumulated contribution rate was 82.55%);the warmest season precipitation, the annual average temperature, the slope and altitude had the greatest influence on the geographical distribution of Stipa (accumulated contribution rate was 73.47%).Soil texture had little effect on this three grassland species (accumulated contribution rate < 5%).In general, the hydrothermal conditions in the growing season were the most important climatic factors that restrict the distribution of dominant species on the Tibetan Plateau.
Key words Dominant grass species;Maxent model;Distribution pattern;Environmental factor;Tibetan Plateau
基金项目 青海省科学技术厅项目(2022-ZJ-718);重庆市科学技术局项目自然科学基金面上项目(cstc2021jcyj-msxmX0384)。
作者简介 聂学敏(1980—),男,青海民和人,高级工程师,硕士,从事生态环境监测与评价工作。
收稿日期 2024-01-19
物种地理分布格局研究是分析物种的空间分异规律以及对环境因子响应的生物地理学研究的重要领域,也是保护生态系统多样性的基础[1]。在植被地理分布研究中,物种分布模型有着广泛的应用,该模型对输入数据要求不高、操作简便、模拟效果较好,在物种、群落和生态系统分布的模拟等方面都有着重要的应用[2]。该模型需要环境因子数据以及研究对象的分布点位数据作为输入数据,利用研究对象的不完全样本点位数据,以环境因子预测值为该因子的经验平均值,模拟出满足熵值最大的最佳分布,并反映物种已知分布区的环境因子与研究区之间的关系。Maxent(最大熵)模型因其不受有限的样本分布点位以及空间偏差数据影响,且模拟准确性较其他物种分布模型高,在模拟物种分布领域应用广泛。青藏高原是我国天然草地分布最广泛的区域,也是全球气候变化的敏感区与脆弱地带[3],在气候变化影响下,全球范围内温度持续升高,降水格局改变,导致草地的地理分布与生产力发生了变化。笔者拟在有限的数据基础上,通过物种分布模型分析青藏高原草地优势种草的分布情况及其对主导环境因子的响应,以期为青藏高原草地资源的评价、保护、持续利用以及植物资源分布的研究提供参考。
1 材料与方法
1.1 研究区概况
青藏高原被称为“地球第三极”,是世界海拔最高、全球气候变化最敏感的区域之一[4]。青藏高原植被类型齐全,草地面积广阔,主要包括高寒草甸、高寒草原、温性草原3种类型,其中,针茅、苔草、蒿草为主要的优势物种。青海省与西藏自治区是青藏高原主体,约占高原总面积的80%,草地样本点位数据也集中分布于这2个地区所在区域,该研究以青海省和西藏自治区为研究区域。
1.2 数据来源
利用资源环境数据云平台(http://www.resdc.cn)获取1∶100万的研究区植被类型空间分布栅格数据和土壤质地数据;通过中国数字植物标本馆(http://www.cvh.org)获取草地植被分布样本数据,共获取1 922条标本记录;通过中国气象科学数据共享服务网(http://cdc.cma.gov.cn)获取青藏高原及其周边51个气象站点月度的温度与降水量数据,然后在ArcGIS 10.2中采用Kriging法将气象变量插值成1 000 m的栅格数据;通过地理空间数据云平台(http://www.gscloud.cn/)获取研究区分带的30 m分辨率DEM数据;通过国家基础地理信息中心(http://www.ngcc.cn)获取1∶400万研究区的行政矢量边界数据。其中所有环境因子及其代码如表1所示。
1.3 模型模拟。
1.3.1 模型原理。Maxent模型的原理是根据已有的不完全
数据来模拟和预测未知信息。以已知信息为约束条件,模拟
出使熵值最大化的最优条件。在构建模型时,不需要做任何倾向性假设,在保证已有数据完好前提下,当满足熵值最大化条件时,删除冗余信息,使得未知信息的确定性升高[5]。设β为随机变量,B1,B2,B3,...,Bn是β变量的n种可能,则随机变量的熵值为:
R(β)=-ni=1D1logDi
式中:R(β)为随机变量的熵值;D1,D2,D3,…,Di为每种变量可能发生的概率。
当已知信息为物种分布数据时,构建Maxent模型,输出得到熵值最大的最优分布,即该物种的潜在适宜分布区。
1.3.2
模型运行。将环境因子数据通过重采样转化成相同的栅格大小(30 m×30 m),将坐标统一为WGS1984地理坐标系,并输出为二进制*.asc格式,与草地点位数据一起输入Maxent 3.4.1中进行分布模拟及验证。随机选取草地分布样本总数的75%作为训练子集构建模型[6],25%作为测试集验证模型,重复运行次数设定为20次。模型输出结果类型为Logistic值,表示物种分布概率,介于0~1,“0”表示不存在,“1”表示存在,越接近1则存在概率越大,物种适应度越高。
1.3.3
模型验证。利用接受者操作特征曲线(receiver operating characteristic curve,简称ROC曲线)对模型预测效果进行评价,曲线下的面积(Area Under ROC Curve,简称AUC值)表示模型精度,取值范围为0.5~1.0,当0.5≤AUC<0.6时,表示预测结果准确度较差;当0.6≤AUC<0.7时,表示准确度一般;当0.7≤AUC<0.8时,表示较准确;当0.8≤AUC<0.9时,表示很准确;当0.9≤AUC≤1.0时,表示极准确。该研究中模拟20次重复的AUC值为0.873左右,表明预测准确度很好,ROC曲线如图1所示。
2 结果与分析
2.1 主导环境因子筛选
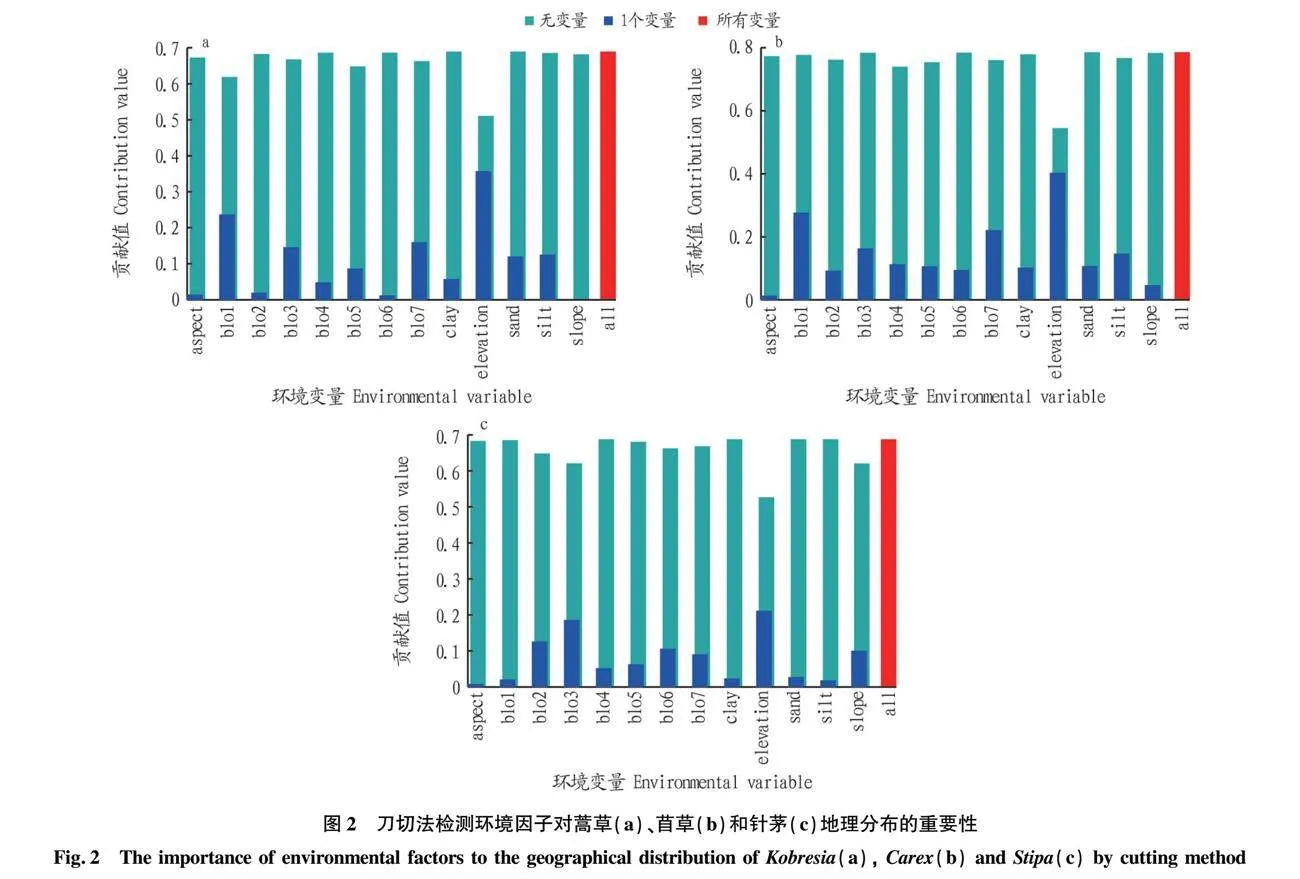
通过刀切法对环境因子贡献值进行评估,结果如图2所示,图中浅蓝色条带表示去除某一变量后环境因子对物种地理分布的贡献(简称无变量),深蓝色条带表示仅保留某一变量时环境因子对物种地理分布的贡献(简称1个变量),红色条带表示所有环境因子综合作用于物种地理分布的贡献(简称所有变量),条带越长表示贡献值越大。浅蓝色条带与红色条带长度相差越大,表明去除该变量对于物种地理分布模拟结果负面影响越大,则该变量的不可替代性越高,重要性也相对越高[7]。结果表明,对于3种主要优势种草,仅保留海拔这一变量时,其对于地理分布增益贡献值最大,而去除该变量后余下的变量与所有变量综合作用贡献值相比,减少的得分值最多,说明海拔是影响优势种草分布的主导环境因子。在所有环境因子中,最冷季降水量(bio1)、温度季节性变化标准差(bio5)、年降水量(bio7)、海拔(elevation)对嵩草分布的贡献较大,累计贡献率均大于80%;最冷季降水量(bio1)、最暖季均温(bio4)、年降水量(bio7)、海拔(elevation)是对苔草分布影响较大的环境因子,累计贡献率达到82.55%;最暖季降水量(bio3)、年均温(bio6)、坡度(slope)、海拔(elevation)对针茅分布影响较大,累计贡献率达到73.47%。对于3种优势种草,砂土(sand)、粉砂土(silt)与黏土(clay)含量对其分布的累计贡献率不超过5%,说明土壤质地对草地物种地理分布的影响不大,因为青藏高原高寒严峻的生长条件,土壤在一年中大部分时间(9月到次年5月)为冻结状态,气象因子成为制约植物生长的主导因素[8]。
2.2 不同优势种草潜在适生区面积及其空间分布
统计不同优势种草潜在适生区面积(表2)可知,每种优势种草高适生区(存在概率0.50~1.00)面积最小,低适生区(存在概率0.05~0.30)面积最大,说明潜在低适生区在研究区占主导,约占草地总面积的50%。苔草的低适生区面积最大(113.62万km2),3种优势种草的高适生区面积相差不大,其中嵩草最大(29.52万km2)。3种优势种草高适生区表现出一定的一致性。藏东北、青海湖周边地区以及唐古拉山脉以南地区是3种优势种草分布的高适生区,与李旭谦[9]的研究结果一致;青海南部以及西藏西部的昌都、南部的山南是嵩草和苔草的高适生区。此外,少部分的嵩草、苔草和针茅高适生区分布于阿里地区以南。苔草和嵩草的不适生区主要分布于藏北高原东南部以及羌塘无人区;与其他2种优势种草不同的是,针茅不适生区主要分布在藏西南的山南和林芝。
2.3 优势种草分布格局与主导环境因子的关系
2.3.1 嵩草对主导环境因子的响应。
根据嵩草分布对主导环境因子响应的Logistic曲线(图3)发现,嵩草分布对最冷季降水量(bio1)、温度季节性变化标准差(bio5)和海拔(elevation)的响应相似,随着环境变量的增加,存在概率呈先增加后减少的趋势。当最冷季降水量为26 mm左右时,存在概率达到峰值;温度季节性变化标准差和海拔分别达到8.5、3 500~4 000 m时,嵩草存在概率最高,达到其适生区范围。当温度季节性变化标准差>10时,嵩草存在概率小于0.05,表明温度季节性变化过大时会对嵩草生长产生不利影响;当海拔高于5 500 m时,嵩草存在概率小于0.05,因为海拔越高,温度越低,青藏高原海拔大于5 000 m的地区多被冰雪覆盖,植被生长极其困难。年降水量(bio7)超过240 mm后,嵩草存在概率随年降水量的增加而增加。已有研究表明降水是制约青藏高原许多物种分布的主要因素[8]。由于青藏高原蒸散量很大(年蒸散量700~1 200 mm)[10],且降水是青藏高原植物生长的重要水分来源,因此,降水成为植被覆盖年际变化的主要影响因素[11]。
2.3.2 苔草对主导环境因子的响应。
从图4可以看出,苔草分布随着对年降水量(bio7)与海拔(elevation)的增加呈现先增加后减少的趋势。年降水量为630~650 mm最适宜苔草生长;海拔为3 500~4 000 m的区域最适宜苔草分布。当年降水量小于100 mm,或海拔大于6 000 m时,苔草的存在概率降至0.05以下,表明苔草耐受区域的最高海拔为6 000 m。苔草分布对于最冷季降水量(bio1)与最暖季均温(bio4)的响应表现出先减后增的趋势。当最冷季降水量为3~5 mm时,苔草存在概率为0.10~0.15,为苔草低适生区范围;当最暖季均温为10~11 ℃时,苔草存在概率降至0.42左右,为苔草中适生区范围。最适宜苔草生长的最冷季降水量与最暖季均温分别为40 mm、19 ℃。结果表明,苔草的地理分布受温度与降水量的综合作用。
2.3.3 针茅对主导环境因子的响应。
针茅分布对主导环境因子响应的Logistic曲线(图5)显示,针茅分布对最暖季降水量(bio3)呈波峰波谷交错的趋势,当降水量为60~70、300 mm时,出现2个波峰,两者的值均大于0.50(高适生区);当降水量为100~260 mm时,存在概率稳定在0.35左右,属于针茅中适生区。针茅分布对年均温(bio6)响应曲线呈现先增后减的趋势,在-2~2 ℃时,针茅的存在概率稳定在0.55左右,随后缓慢下降,在年均温大于3 ℃后下降速度加快,直到11 ℃时到达最低值(小于0.05)。与其他2种优势种草相似,随着海拔的升高,针茅的存在概率呈先增加后减少的趋势,当海拔为3 500~4 500 m时,存在概率接近0.6,较适宜针茅生长;当海拔高于6 500 m时,针茅存在概率降到最低。针茅对于坡度的响应呈缓慢上升再急剧下降的趋势。结果表明,针茅对生长季节降水量依赖性较大,而年降水量对针茅分布贡献率很小,累计不超过6%,所以尽管阿里地区南部虽然年降水量稀少,冬季温度较低,但降水集中在5—10月,故该地区针茅广泛分布。
综上所述,3种优势种草分布对海拔的响应均呈现先增加后减少的趋势,最适宜的海拔均在4 000 m左右。最暖季均温(bio4)仅对苔草分布有较大影响,表明相对其他2种优势种草来说,苔草对生长季温度要求更高,在一定范围内(10~19 ℃),最暖季均温越高,越适宜苔草生长。当温度季节性变化标准差(bio5)大于8.5时,嵩草存在概率均开始下降,表明温度季节性变化过大不利于其生长。
2.4 不同优势种草耐受主导环境因子阈值分析
根据3种优势种草对主导环境因子响应曲线,分析其对部分主导环境因子耐受的最低或最高阈值,并进行横向比较(表3)。针茅耐受的海拔阈值最大(6 500 m),其他2种优势种草都不高于6 000 m。表明针茅对寒冷与多风环境的耐受性较高,对生长季降水量的需求较大,与胡忠俊等[12]的研究结果一致。由不同优势种草对温度季节性变化标准差(bio5)的响应可知,针茅容忍度最大(10.0),嵩草容忍度最低(8.5),从另一个方面反映了针茅对温度容忍度较大的特点。
2.5 环境因子影响优势种草分布的内在机制
为了进一步分析水热因子的空间分异对优势草种分布的影响,绘制了4种主导气候因子(最暖季均温、最冷季降水量、最暖季降水量、年降水量)空间分布图(图6)。结果表明,3种优势种草共同的适生区藏东北青海湖周边地区以及唐古拉山脉以南地区有着共同点:年降水量充沛(487.31~681.78 mm)、最暖季降水量充沛(207.40~329.43 mm);且藏东北地区最冷季降水量也较丰富(22.41~31.30 mm),而唐古拉山脉以南地区比较匮乏(4.62~17.39 mm),说明最暖季降水量对处于生长季的植物来说非常重要,而最冷季降水量对植物制约不大。喜马拉雅山中断北坡存在小范围的适生区,尽管该地区最暖季降水量较为匮乏(54.85~85.36 mm),但是该地区位于雅鲁藏布江源地,冰川广布,最暖季冰川融水与解冻的土壤中的水为生长期的草地提供足够的水分,弥补了降水的不足。青海省北部以及藏东南地区最暖季均温较高(12.84~19.27 ℃),但是青海省西北部是3种优势种草的低适生区,分析原因,该地区最暖季降水量较少(54.85~176.88 mm),说明生长季水热综合条件是制约青藏高原优势物种分布的最主要气候因子。冈底斯山脉北侧为羌塘高原内流区(羌塘无人区),气候极度严寒干燥,为3种优势种草的不适宜分布区域。
3 结论
基于Maxent模型,对嵩草、苔草与针茅3种优势种草空间分布、影响因子、耐受阈值等进行综合分析,认为海拔是影响优势种草分布的主导环境因子,而土壤质地对地理分布影响较小。每种优势种草潜在低适生区在研究区占主导,而高适生区面积相差不大,分布区表现出一定的一致性。对优势种草耐受气候因子阈值分析表明,针茅对温度、海拔等因素容忍度较大,对生长季降水量的需求较大。进一步分析水热因子的空间分异对优势草种分布的影响,结果显示3种优势种草共同的适生区是藏东北青海湖周边地区以及唐古拉山脉以南地区,青海省西北部是3种优势种草的低适生区,冈底斯山脉北侧为羌塘高原内流区(羌塘无人区)且是3种优势种草的不适宜分布区域,说明生长季水热综合条件是制约青藏高原优势物种分布的最主要气候因子。
参考文献
[1] GUISAN A,THUILLER W.Predicting species distribution:Offering more than simple habitat models[J].Ecology letters,2005,8(9):993-1009.
[2] 李国庆,刘长成,刘玉国,等.物种分布模型理论研究进展[J].生态学报,2013,33(16):4827-4835.
[3] 孙鸿烈,郑度,姚檀栋,等.青藏高原国家生态安全屏障保护与建设[J].地理学报,2012,67(1):3-12.
[4] PAULI H,GOTTFRIED M,REITER K,et al.Signals of range expansions and contractions of vascular plants in the high Alps:Observations(1994-2004)at the GLORIA master site Schrankogel,Tyrol,Austria[J].Global change biology,2007,13(1):147-156.
[5] 李灿,刘贤安,王娟,等.基于MaxEnt模型的四川省红豆杉潜在分布区分析及适宜性评价[J].林业调查规划,2018,43(1):22-29.
[6] 刘文胜,游简舲,曾文斌,等.气候变化下青藏苔草地理分布的预测[J].中国草地学报,2018,40(5):43-49.
[7] 孙敬松,周广胜.利用最大熵法(MaxEnt)模拟中国冬小麦分布区的年代际动态变化[J].中国农业气象,2012,33(4):481-487.
[8] WU L,WANG M,OUYANG H,et al.Spatial distribution modelling of Kobresia pygmaea(Cyperaceae)on the Qinghai-Tibetan Plateau[J].Journal of resources and ecology,2017,8(1):20-29.
[9] 李旭谦.青海湖流域草地类型及其分布[J].青海草业,2009,18(4):20-23,19.
[10] ZHANG X Q,REN Y,YIN Z Y,et al.Spatial and temporal variation patterns of reference evapotranspiration across the Qinghai-Tibetan Plateau during 1971-2004[J].Journal of geophysical research,2009,114:1-14.
[11] XU X K,CHEN H,LEVY J K.Spatiotemporal vegetation cover variations in the Qinghai-Tibet Plateau under global climate change[J].Chinese science bulletin,2008,53(6):915-922.
[12] 胡忠俊,张镱锂,于海彬.基于MaxEnt模型和GIS的青藏高原紫花针茅分布格局模拟[J].应用生态学报,2015,26(2):505-511.
