
含笑花雄蕊发生与发育的基本特征
2024-12-12 00:00:00刘思彤聂堂杰吴庆贤靳乐妮万小霞尹增芳
南京林业大学学报(自然科学版) 2024年6期


摘要:【目的】观察含笑花(Michelia figo)雄蕊形态建成过程,探究其雄蕊发生和发育的基本特征,为木兰科植物繁育与进化生物学的研究积累基础资料。【方法】利用扫描电镜、显微成像及石蜡切片技术,对不同生长阶段含笑花花芽中的雄蕊进行解剖观察。【结果】含笑花雄蕊原基的发生方式为螺旋状排列、向心方式发生。随着雄蕊形态建成的完成,其表观形态特征发生系列变化,最终形成较长的花药与较短的花丝。花药组织包括花药隔和4个花药室。花药室内造孢组织分化形成的小孢子母细胞减数分裂不同步,最终形成左右对称型和正四面体型小孢子四分体,之后游离的小孢子发育成二细胞型花粉和少量的三细胞型花粉。花药壁由1层表皮细胞、1层药室内壁细胞、2~3层中层细胞及1层分泌绒毡层细胞组成。成熟花粉发育时期绒毡层解体消失,但部分中层细胞宿存,直至花药壁开裂时才完全解体消失。花丝外有1层表皮细胞包裹,内部由薄壁组织填充,中央贯穿着直达药隔的维管束。发育早期的花丝薄壁组织内具有气腔的结构,成熟期消失。【结论】含笑花雄蕊发生与发育的过程基本正常,雄蕊螺旋状向心发生、小孢子母细胞减数分裂不同步、花丝气腔结构的时序变化、多数左右对称型小孢子四分体及二细胞型花粉等基本特征,体现了木兰科植物的原始性。
关键词:含笑花;雄蕊发生;花丝;花药;小孢子四分体
中图分类号:S718; Q944.33"""" 文献标志码:A开放科学(资源服务)标识码(OSID):
文章编号:1000-2006(2024)06-0034-07
The basic characteristics of stamen morphogenesis and" development in Michelia figo
LIU Sitong1, NIE Tangjie1,WU Qingxian1, JIN Leni1, WAN Xiaoxia2, YIN Zengfang1
(1.Co-Innovation Center for Sustainable Forestry in Southern China, College of Life Science, Nanjing Forestry University, Nanjing 210037, China;2. Cryo-electron Microscope Center, Zhejiang University, Hangzhou 310058, China)
Abstract: 【Objective】The fundamental characteristics of stamen morphogenesis and development were described according to the observation of stamen morphogenesis in Michelia figo. The findings provide valuable basic data for future investigations in plant breeding and evolutionary biology within the Magnoliaceae family. 【Method】Using the" scanning electron microscopy and paraffin section technology, the stamens of flower buds in M. figo at different growth stages were observed. 【Result】The formation pattern of stamen primordium in M. figo was found to be helical and centripetal. Upon the completion of stamen morphogenesis, distinct morphological changes were observed, leading to the formation of a longer anther and a shorter filament. The anther tissue consists of anther septa and four anther locules. Normally, the sporogenous tissue differentiates gradually into microspore mother cells in the anther locule. The microspore mother cell meiosis was not synchronized, and eventually the isobilateral and tertrahedroid microspore tetrads were formed. Then, the free microspore developed into two-celled pollen and a few three-celled pollen. The structure of the anther wall was composed of one layer of epidermis cells, one layer of endothecium cells, two to three layers of middle layer cells, and one layer of secretory tapetum cells. During the development of mature pollen, the tapetum cells disintegrated and disappeared, but some middle layer cells persisted until the anther wall underwent dehiscence. The filaments were surrounded by a single layer of epidermis cells on the outer surface. Internally, the filaments were composed of parenchyma tissue, and a vascular bundle ran through the center toward the connective tissue. There was the cavity structure in the parenchyma of the filaments during the earlier development stage, which later disappeared during the mature stage of the stamen.【Conclusion】The process of stamen morphorgenesis in M. figo is normal compared with other Michelia plant species. The fundamental characteristics observed, including the helical centripetal formation of stamens, asynchronous meiotic division of microspore mother cells, temporal changes of the cavity structure in filaments, predominance of isobilateral tetrads, and two-celled pollen reflected the inherent nature of the Magnoliaceae family.
Keywords:Michelia figo; stamen morphorgenesis; filament; anther; microspore tetrad
雄蕊是被子植物进行有性生殖的重要繁殖器官,包括花药和花丝两部分,花药是产生花粉及雄配子体发育的场所,而花丝则为花药发育提供所需营养物质[1]。因此,雄蕊结构与功能的正常行使对被子植物的繁殖至关重要,在雄蕊发育过程中任一环节发生异常都将对植物生殖的有效性产生显著影响,进而导致其自我更新过程受阻等问题,所以探究雄蕊发育的过程对研究植物繁殖、控制作物育种及杂交种子生产具有重要意义[2-3]。目前,已有众多学者对雄蕊的发育及近缘种间生殖发育的异同进行了探究。如Zhang等[4]为研究防己科(Menispermaceae)植物不同类型雄蕊的发育情况,详细观察了该科植物雄蕊发育过程;Ning等[5]观察了独叶草(Kingdonia uniflora)可育雄蕊和退化雄蕊的分化过程及退化雄蕊蜜腺的结构,对单花雄蕊数量和个体发育变化进行了分析。作为最古老被子植物类群之一的木兰科(Magnoliaceae)植物,其雄性生殖器官的发育也受到了广泛的重视。关于木兰属(Magnolia)[6]、玉兰属(Yulania)[7-8]、木莲属(Manglietia)[9]和含笑属(Michelia)[10]等植物已积累了较多有关小孢子发生及雄配子体发育的胚胎学资料,总体上木兰科各属植物花药的发育过程普遍表现出了较为原始的特征,但在紫玉兰(Yulania liliiflora)和二乔玉兰(Y. × soulangeana)中发现了较为进化的胞质分裂方式[11-12]。因此,木兰科植物雄性生殖器官发育的基本特性研究是一个值得关注的科学命题。
含笑花(Michelia figo)原产中国,为木兰科含笑属常绿灌木,花期较长且具有浓郁的花香,常作为观赏植物用于景观营造或园林绿化[13];其花瓣具有祛淤生新、活血止痛的功效,是重要的药用植物资源,亦可被制成花茶或香料[14]。因此,含笑花是极具发展潜力的优良观赏和香料树种,具有较高的经济价值。目前,国内外对含笑花的研究主要集中在化学成分[15]、药用作用[16]、抗寒性[17]、繁殖技术[18]、生理学[19]、孢粉学[20]及基因组学[21]等方面,其有性生殖方面的研究鲜有详细而严谨的报道,而厘清含笑花的生殖发育特征是未来开展育种工作的前提和基础。本研究利用形态解剖学的方法对含笑花雄蕊的发生及发育过程进行观察,旨在揭示其形态建成与发育的一般规律,为利用含笑花为父本开展分子精准设计杂交育种及种质资源创新积累基础数据资料。
1 材料与方法
1.1 试验材料及观察方法
以栽培于南京林业大学校园(118°48′40″E,32°4′44″N)内、长势良好的3 株9年生含笑花为试验材料,采用常规方法栽培管理。
2021年5月中旬至翌年4月中下旬,每隔15 d观察含笑花腋芽形态特征。每次随机选取3 枚腋芽,用游标卡尺测量其长度、宽度,在体视显微镜(日本,Nikon SMZ18)下进行解剖并拍照。随后常规石蜡制样、切片机(德国,Leica RM 2255)切片,切片厚度为8 μm,经番红-固绿对染,中性树胶封片后,置于显微镜(日本,Nikon 50i)下观察拍照。
2021年5月下旬至6月中旬,每隔5 d采集腋芽,用于扫描电镜制样。样品经体积分数70% FAA固定、乙醇系列脱水、乙酸异戊酯置换,在临界点干燥仪(英国,EMITECH K850)内干燥样品24 h,离子溅射仪(日本,日立 E1010)中喷金120 s,OXFORD Inca X-act(美国,FEI)能谱仪扫描雄蕊发生过程的微观形态学特征变化并拍照记录。
1.2 数据处理
应用Excel 2021、Photoshop CC 2020完成数据处理和图表制作。
2 结果与分析
2.1 含笑花雄蕊发育过程形态特征的变化
5月下旬含笑花完成成花转变,随后花芽发育;6月初,腋生花芽肉眼依稀可辨。花芽发育过程见表1。体视显微镜下解剖观察发现,在花被片原基内侧雄蕊原基开始分化(图1)。图1a显示在圆锥形的生殖顶端基部出现半球形状突起,第2枚雄蕊原基与第1枚交互排列,并呈螺旋状向顶发生(图1b)。扫描电镜下花托上雄蕊原基依序发生
(图1c),在发生后快速生长,并逐渐形成扁条形(图1d)。6月中旬雄蕊形态建成渐次完成,但表观上雄蕊花药和花丝未明显分化(图1e),此时雌蕊原基开始发生(图1d、1e)。7月上旬,在雌蕊原基发育形成条形叶片形态时,雄蕊初步分化成花药和花丝的结构(图1f)。8—11月,含笑花花芽生长缓慢,图1g显示的是11月下旬去除花被片后花芽的形态,可见雄蕊可以区分为花药和花丝两个部分。随后雄蕊伸长生长速度较快,图1h显示雌蕊被雄蕊包裹。12月下旬至翌年2月上旬,雄蕊形态分化更加明显,表观上有紫红色色素沉积(图1 i、1j)。2月初至3月初是雌蕊的快速分化期,此时可以观察到心皮逐渐发育成熟并突出雄蕊之上,雄蕊紫色色素逐渐消失,仅花药先端呈紫色,表观略带紫色条纹,花丝伸展明显,呈紫红色(图1j、1k、1l)。4月上旬,含笑花进入花期,开放的花内雄蕊花药纵裂,呈散粉状态(图1n、1o)。
2.2 含笑花雄蕊发育过程结构特征的变化
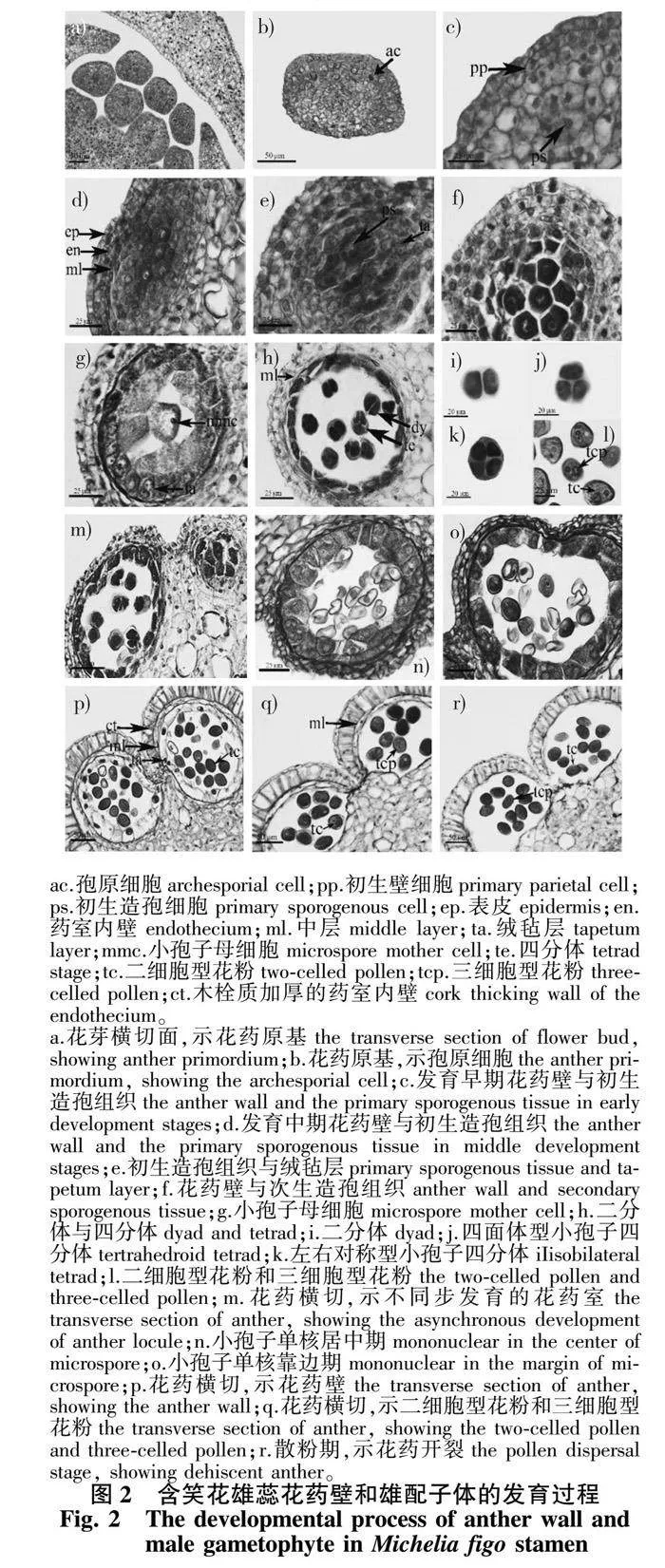
2.2.1 花药壁的发育
依据Davis[22]的分类标准,含笑花的花药壁发育类型为基本型,发育动态过程如表1所示。发育初期的花药原基,由一团细胞壁薄、细胞质浓厚的细胞组成(图2a)。图2b显示的是单个花药原基的横切面,表面的一层细胞为花药原基的表皮原,其内部孢原细胞已经形成。图2c为花药原基角隅处的半圆形突起部分,内部已形成造孢组织,周缘细胞也初步完成分裂。随后由周缘细胞分裂产生的细胞规律排列在表皮原细胞内侧,构成花药壁层,此时的花药壁层包括1层表皮细胞、1层药室内壁细胞、2~3层中层细胞,绒毡层细胞尚未分化形成(图2d)。在造孢组织向小孢子母细胞转化期,绒毡层细胞开始分化,成熟的绒毡层细胞较大、细胞质浓厚、内含多个核(图2e—2g)。图2h显示小孢子二分体、四分体时期的花药壁层细胞,表现为中层细胞未完全消失,局部区域仍保留1~2层中层细胞。随着小孢子的发育,花药壁层细胞形态变化明显,横切面观察初期表皮细胞呈扁平方形(图2n),随后表皮细胞逐渐液泡化(图2o);药室内壁细胞径向伸长,呈排列整齐的扁长方形,并形成“U”形木栓质增厚壁,但在两个花粉囊的连接处,药室内壁细胞未发生细胞壁的次生增厚,为唇细胞区。同时,绒毡层表现出分泌绒毡层的典型特征,如细胞逐渐空泡化(图2p),图2q显示绒毡层最终完全解体消失,但此时尚有未解体的中层细胞残迹位于药室内壁内侧。在成熟花粉时期,伴随唇细胞的解体,中层和绒毡层细胞完全解体消失,花药壁呈开裂状态(图2r)。
2.2.2 小孢子发生及雄配子体发育
含笑花花药内造孢组织时期持续时间较长,从花药原基开始分化的6月初直至11月下旬(表1)。在此期间,初生造孢细胞不断进行有丝分裂,形成次生造孢细胞(图2c—2f)。随后次生造孢细胞逐渐形成胼胝质的壁,分化形成小孢子母细胞(图2g)。小孢子母细胞经过减数分裂,在同一花药室内可观察到既有二分体又有四分体小孢子的发育不同步的现象(图2h)。图2i—2k显示的是二分体小孢子(图2i)与四分体小孢子,其中小孢子四分体具有两种不同发育类型,包括四面体型(图2j)和左右对称型(图2k)。此时,偶尔会观察到同一个花药两个相邻药室也存在发育不同步的现象。一个药室内小孢子母细胞正在进行减数分裂,另一个药室还处于次生造孢组织状态(图2m)。翌年2月中下旬,小孢子四分体共同的胼胝质壁降解消失,此时游离的小孢子细胞质浓,细胞核位于中央,为小孢子单核居中期(图2n)。随后小孢子液泡化明显,细胞核及细胞质被挤向细胞壁的一侧,为小孢子单核靠边期(图2o)。3月中旬,单核小孢子进行有丝分裂形成1个大的营养核和1个小的生殖核,发育为二细胞型花粉(图2p),部分生殖核会继续分裂形成两个精细胞,发育形成三细胞型花粉(图2l),因此,含笑花的同一药室内中同时存在二细胞型和三细胞型花粉(图2q、2r)。
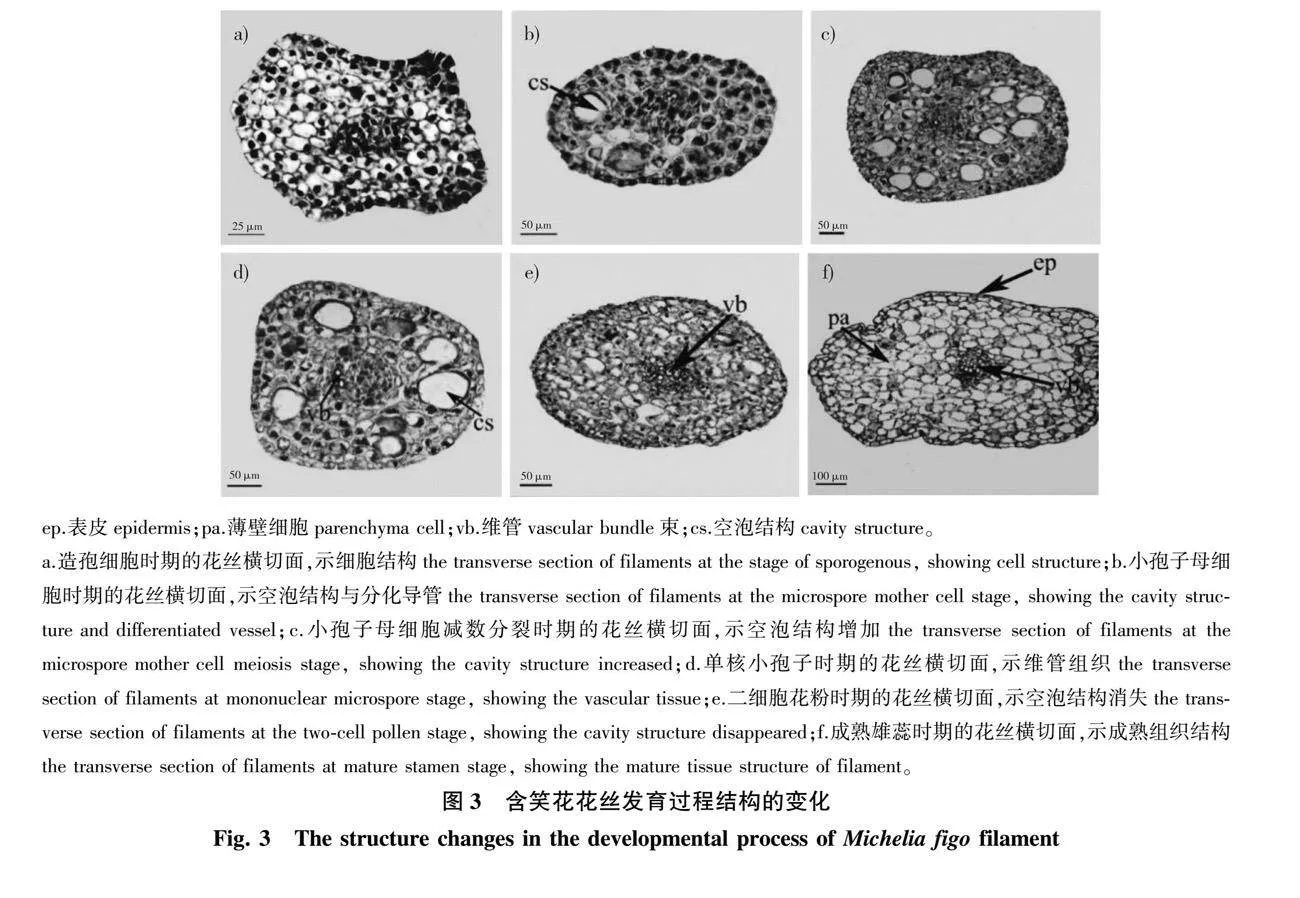
2.2.3 花丝的发育
7月上旬,雄蕊原基逐渐发育形成花药和花丝(表1)。观察横切面发现,花丝由1层表皮细胞及内部的薄壁组织组成,其中位于花丝中心部位的薄壁细胞体积较小(图3a),随后逐渐发育形成维管组织,此时薄壁组织区域出现空泡状结构(图3b)。在小孢子母细胞减数分裂时期,薄壁组织内空腔的数量达到最多(图3c);翌年1—2月,在小孢子四分体形成后,薄壁组织空腔数量逐渐减少,此时维管组织的细胞数量开始增加(图3d);3月下旬,二细胞型花粉逐渐发育成熟,可见花丝中央的维管束数量明显增多(图3e);4月初,花药纵向开裂,此时花丝的表皮细胞呈扁平的长方体状,内部的薄壁细胞体积增加,气泡状结构消失殆尽(图3f)。
3 讨 论
3.1 含笑花雄蕊发育过程的原始特征
含笑花隶属于被子植物中系统演化地位较原始的木兰科,在其雄蕊形态建成过程中,部分特征表现出了一定的原始性。现有的研究资料表明,木兰科植物桂南木莲(Manglietia conifera)[23]及观光木(Tsoongiodendron odorum)[24]的雄蕊原基均在花被片原基内侧螺旋状向顶发生,含笑花的雄蕊原基也呈现同样的发生方式,与营养枝叶序形成模式一致。在广东木莲(Manglietia" kwangtungensis)、长叶木兰(Magnolia paenetalauma)及焕镛木(Woonyoungia septentrionalis)中,雄蕊的基部宽而短,没有形态的分化,而紫花含笑(Michelia crassipes)的雄蕊则明显分化为花药和花丝[25]。本研究发现,含笑花雄蕊发生初期花药与花丝表观特征几乎难以区分,在花药形成造孢组织时,花药与花丝分化形成。对花丝的组织学特征进行观察发现,其最明显的变化是气腔结构在发育过程中从无到有的改变。万小霞[26]认为深山含笑(Michelia maudiae)花芽苞片中气腔结构的形成预示着抗寒能力的增加。与深山含笑花芽外的苞片组织结构相似,含笑花花丝组织中气腔结构的出现也表现出季节性的特征,说明花丝的组织结构与苞片的叶性特征的变化相吻合。特别地,含笑花雄蕊发育过程中的原始性可能与其生态适应性有关,这些气腔结构的出现可能是其对环境适应的需要[27]。
植物的原始特征在进化过程中会被保留下来,并在后续的演化中发生了一些变化[28]。本研究发现含笑花的绒毡层为分泌型,仅具1层多核细胞,与对红花玉兰(Y. wufengensis)[7] 的研究结果一致。但香木莲(Michelia aromatica)[29]及灰木莲(Manglietia conifera)[9]绒毡层则具有多层细胞,这种绒毡层细胞结构差异的原因可能与其进化历史有关。此外,含笑花的中层细胞至花粉完全成熟依然存在,在花药壁开裂前才彻底消失,其长时间宿存的原因、对花粉发育的影响以及是否为进化的特征还有待进一步的探究。现有的研究资料还证实,木兰科植物小孢子四分体多为原始的左右对称型[10,30],而含笑属的西畴含笑(Michelia coriacea)[31]小孢子四分体为四面体型,并将其视为一种进化的类型。含笑花具有左右对称型小孢子四分体,还可以观察到四面体型小孢子四分体,但数量上明显少于前一种类型。此外,研究资料还显示深山含笑[10]、西畴含笑[31]、广西含笑(M. guangxiensis)[32]的成熟花粉均为原始的二细胞型,而熬成齐[33]的研究则显示含笑的花粉为三细胞型花粉。根据笔者的研究结果,在含笑花中二细胞型花粉和三细胞型花粉是同时存在的,且前者数量明显多于后者。因此,笔者推测这种四分体类型的变化、二细胞型和三细胞型花粉的存在具备一定的系统演化意义,值得进一步探讨。
3.2 花粉发育不同步现象的生物学意义
在含笑花小孢子发生及雄配子体发育过程中,同一药室内花粉的发育在小孢子母细胞减数分裂前基本同步,但在减数分裂后出现了不同步的现象,即在同一花药室内,同时存在二分体及四分体,这一现象与对焕镛木报道的结果[34]类似。Hu等[35]研究发现白玉兰(Y. denudata)小孢子母细胞减数分裂前期Ⅰ之前,花药室内小孢子母细胞的分化是同步的,但随后核分裂出现了明显的不同步现象,小孢子母细胞胞质分裂方式也是双向性的,分析其作为基部被子植物类群,上述特征是其原始性的具体体现。至于不同花药间或同一花药的不同花药室内,存在小孢子的发育进程相差1~2个时期的现象在含笑花花粉发育过程中更为常见,这与百合科(Liliaceae)植物类似[36]。此外,花粉发育的不同步现象可能与植物繁殖策略有关。含笑花是一种以异交为主且异交需要传粉者的植物[37],这种发育不同步能够增加花朵的持续开放时间,从而提高异花授粉的机会;当一部分花粉还未完全成熟时,已成熟的花粉就可以被传递给传粉媒介,从而增加了花粉的传播范围和数量,以适应传粉媒介的活动规律和环境变化。
参考文献(reference):
[1]LIU X L,TAN X Y.Mutation in Arabidopsis MOR1 gene impairs endocytosis in stamen filament cells and results in anther indehiscence[J].Plant Growth Regul,2022,96(2):303-314.DOI: 10.1007/s10725-021-00777-7.
[2]GMEZ J F,TALLE B,WILSON Z A.Anther and pollen development:a conserved developmental pathway[J].J Integr Plant Biol,2015,57(11):876-891.DOI: 10.1111/jipb.12425.
[3]MARCHANT D B,WALBOT V.Anther development: the long road to making pollen[J].Plant Cell,2022,34(12):4677-4695.DOI: 10.1093/plcell/koac287.
[4]ZHANG H Y,ZHANG X Y,NING K,et al.Stamen and pollen development in Menispermaceae with contrasting androecium structure[J].Plant Syst Evol,2022,308(5):36.DOI: 10.1007/s00606-022-01828-w.
[5]NING K,ZHU Q Q,ZHANG X H.How do fertile stamens and staminodes go differently in Kingdonia uniflora?Evidence from stamen numbers,ontogeny,and structure[J].Flora,2023,303:152293.DOI: 10.1016/j.flora.2023.152293.
[6]张风娟,徐兴友,陈凤敏,等.天女木兰小孢子发生及雄配子体发育的观察[J].经济林研究,2008,26(4):71-75.ZHANG F J,XU X Y,CHEN F M,et al.Observation of microsporogenesis and male gametophyte development in Magnolia sieboldii K[J].Nonwood For Res,2008,26(4):71-75.DOI: 10.3969/j.issn.1003-8981.2008.04.016.
[7]陈丽园,桑子阳,陈发菊,等.红花玉兰大小孢子发生及雌雄配子体发育的研究[J].西北农林科技大学学报(自然科学版),2016,44(9):181-185.CHEN L Y,SANG Z Y,CHEN F J,et al.Sporogenesis and gametophytes development of Magnolia wufengensis[J].J Northwest A F Univ (Nat Sci Ed),2016,44(9):181-185.DOI: 10.13207/j.cnki.jnwafu.2016.09.024.
[8]王姗,沈永宝,鲍华鹏,等.宝华玉兰大小孢子发生和雌雄配子体发育过程中解剖结构的变化[J].植物资源与环境学报,2021,30(3):46-53.WANG S,SHEN Y B,BAO H P,et al.Change of anatomical structure in the process of mega- and microsporogenesis and female and male gametophyte development of Yulania zenii[J].J Plant Resour Environ,2021,30(3):46-53.DOI: 10.3969/j.issn.1674-7895.2021.03.06.
[9]潘丽琴,郝建,徐建民,等.灰木莲花药结构和花粉发育特征[J].林业科学研究,2021,34(6):107-113.PAN L Q,HAO J,XU J M,et al.Anther structure and pollen development characteristic of Manglietia conifera[J].For Res,2021,34(6):107-113.DOI: 10.13275/j.cnki.lykxyj.2021.06.013.
[10]熊海燕,刘志雄.深山含笑大、小孢子发生和雌、雄配子体发育研究[J].植物研究,2018,38(2):212-217.XIONG H Y,LIU Z X.Mega- and microsporogenesis and development of female and male gametophytes in Michelia maudiae Dunn[J].Bull Bot Res,2018,38(2):212-217.DOI: 10.7525/j.issn.1673-5102.2018.02.007.
[11]HAYASHI Y.On the microsporogenesis and pollen morphology in the family Magnoliaceae[J].Sci Rep Tohoku Univ,1960,26:45-52.
[12]DINIS A,SANTO D, MESQUITA J. Ultrastructure of the mature pollen of Michelia figo (Lour.) Spreng. (Magnoliaceae)[J]. J. Submicrosc Cytol Pathol, 2000, 32(4): 591-601.
[13]SRINROCH C,SAHAKITPICHAN P,CHIMNOI N,et al.Phenolic glycosides from Magnolia figo[J].Phytochem Lett,2020,37:110-115.DOI: 10.1016/j.phytol.2019.09.008.
[14]张冬莲,念波,汪志威,等.不同加工工艺对含笑花茶品质的影响[J].中国茶叶加工,2019(1):31-36.ZHANG D L,NIAN B,WANG Z W,et al.Effect of different processing techniques on the quality of Michelia figo tea[J].China Tea Process,2019(1):31-36.DOI: 10.15905/j.cnki.33-1157/ts.2019.01.009.
[15]LIN C L,KAO C L,HUANG S T,et al.Chemical constituents of the flowers of Michelia figo[J].Chem Nat Compd,2019,55(4):781-782.DOI: 10.1007/s10600-019-02811-7.
[16]CHEN J,WANG M,HAN X,et al.First report of leaf blight caused by Colletotrichum karstii on banana shrub (Michelia figo) in Sichuan,China[J].Plant Dis,2023,107(9):2868.DOI: 10.1094/PDIS-04-23-0642-PDN.
[17]亓白岩,周冬琴,於朝广,等.8种含笑属植物的抗寒性研究[J].江苏农业科学,2010,38(5):258-263.QI B Y,ZHOU D Q,YU C G,et al.Study on cold resistance of 8 species of Michelia[J].Jiangsu Agric Sci,2010,38(5):258-263.DOI: 10.15889/j.issn.1002-1302.2010.05.131.
[18]樊光毅,胡烈栋.含笑花苗培育技术探讨[J].园艺与种苗,2018,38(12):18-19.FAN G Y,HU L D. Discussion on the cultivation technology of Michelia figo (Lour.) Spreng seedlings[J]. Hortic Seed,2018,38(12):18-19.DOI: 10.16530/j.cnki.cn21-1574/s.2018.12.007.
[19]TAKEUCHI S,SHINOZAKI K,MATSUSHIMA D,et al.Calibration of the heat ratio method by direct measurements of transpiration with the weighing root-ball method for Michelia figo[J].Acta Hortic,2020(1300):21-28.DOI: 10.17660/actahortic.2020.1300.4.
[20]周瑾,柏永清,曾妍,等.含笑花花粉萌发和花粉管生长的离体培养研究[J].湖北农业科学,2022,61(19):72-77. ZHOU J,BO Y Q, ZENG Y,et al.Study on pollen germination and pollen tube growth culture in vitro of Michelia figo[J].Hubei Agric Sci,2022,61(19):72-77.DOI: 10.14088/j.cnki.issn0439-8114.2022.19.014.
[21]ZHAI M.The complete chloroplast genome sequence of Michelia figo based on landscape design,and a comparative analysis with other Michelia species[J].Mitochondrial DNA B Resour,2020,5(3):2723-2724.DOI: 10.1080/23802359.2020.1788446.
[22]DAVIS G L. Systematic embryology of the angiosperms[M]. New York: John Wiley amp; Sons,1966.
[23]HAO J,PAN L Q,JIA H Y,et al.Floral structure and breeding systems of Manglietia conifera Dandy (Magnoliaceae)[J].Forests,2019,10(9):756.DOI: 10.3390/f10090756.
[24]付琳,曾庆文,徐凤霞,等.观光木的花器官发生[J].热带亚热带植物学报,2007,15(1):30-34.FU L,ZENG Q W,XU F X, et al.Floral organogenesis of Tsoongiodendron odorum Chun[J].J Trop Subtrop Bot,2007,15(1):30-34.DOI: 10.3969/j.issn.1005-3395.2007.01.005.
[25]XU F X,CHEN D Q,SPECHT C.Comparative microsporogenesis and anther development of selected species from Magnoliaceae[J].Nord J Bot,2013,31(3):291-300.DOI: 10.1111/j.1756-1051.2012.01445.x.
[26]万小霞. 三种木兰科植物分枝成花过程及其温度适应性[D]. 南京:南京林业大学, 2021. WAN X X. Study on the process of branching and flowering in three Magnoliaceae species and their temperature adaptability[D]. Nanjing:Nanjing Forestry University, 2021.
[27]MAO P L,ZANG R Z,SHAO H B,et al.The ecological adaptability of four typical plants during the early successional stage of a tropical rainforest[J].Plant Biosyst Int J Deal Aspects Plant Biol,2014, 148(2):288-296.DOI: 10.1080/11263504.2013.770808.
[28]CRAWFORD D J,DOYLE J J,SOLTIS D E,et al.Contemporary and future studies in plant speciation,morphological/floral evolution and polyploidy:honouring the scientific contributions of Leslie D.Gottlieb to plant evolutionary biology[J].Philos Trans R Soc Lond B Biol Sci,2014,369(1648):1-10.DOI: 10.1098/rstb.2013.0341.
[29]PAN Y Z, LIANG H X, GONG X. Studies on the reproductive biology and endangerment mechanism of the endangered plant Manglietia aromatica[J]. J Integr Plant Biol, 2003, 45(3): 311.
[30]王利琳,胡江琴,庞基良,等. 凹叶厚朴大、小孢子发生和雌、雄配子体发育的研究[J].实验生物学报, 2005(6): 490-500. WANG L L, HU J Q, PANG J L, et al. Studies on the megasporogenesis and microsporogenesis and the development of their female and male gametophyte in Magnolia biloba[J]. Chinese J" Exp" Biol, 2006, 38(6): 490-500.
[31]赵兴峰,孙卫邦,杨华斌,等.极度濒危植物西畴含笑的大小孢子发生及雌雄配子体发育[J].云南植物研究,2008,30(5):549-556.ZHAO X F,SUN W B,YANG H B,et al.Mega- and microsporogenesis and development of female and male gametophytes of Michelia coriacea(Magnoliaceae),a globally critical endangered plant in south-east Yunnan of China[J].Acta Bot Yunnanica,2008,30(5):549-556.DOI: 10.3969/j.issn.2095-0845.2008.05.008.
[32]付琳,徐凤霞,曾庆文,等.广西含笑的小孢子发生及雄配子体形成的研究[J].广西植物,2011,31(3):312-317,311.FU L,XU F X,ZENG Q W,et al.Studies of microsporogenesis and male gametophyte formation of Michelia guangxiensis[J].Guihaia,2011,31(3):312-317,311.DOI: 10.3969/j.issn.1000-3142.2011.05.007.
[33]敖成齐.含笑小孢子的发生、雄配子体的发育及其系统学意义[J].广西植物,2007,27(6):836-839.AO C Q.Microsporogenesis,development of male gametophyte in Michelia figo and their systematic significance[J].Guihaia,2007,27(6):836-839.DOI: 10.3969/j.issn.1000-3142.2007.06.006.
[34]FU L,XU F X,ZENG Q W.Embryology of the dioecious Woonyoungia septentrionalis (Magnoliaceae)[J].Nord J Bot,2012,30(2):215-225.DOI: 10.1111/j.1756-1051.2011.01151.x.
[35]HU M L,BAI M,YANG M,et al.Cell polarity,asynchronous nuclear divisions,and bidirectional cytokinesis in male meiosis in Magnolia denudata[J].Protoplasma,2021,258(3):621-632.DOI: 10.1007/s00709-020-01604-y.
[36]KARTAL C.Microsporogenesis,microgametogenesis and in vitro pollen germination in the endangered species Fritillaria stribrnyi(Liliaceae)[J].Caryologia,2015,68(1):36-43.DOI: 10.1080/00087114.2014.998947.
[37]刘晨妮.玉兰与含笑杂交生物学基础研究[D]. 南京:南京林业大学, 2018. LIU C N. The basic research of the hybrid biology of Magnolia and Michelia[D]. Nanjing:Nanjing Forestry University, 2018.
(责任编辑 郑琰燚)
猜你喜欢
工会博览(2022年17期)2022-07-15 07:25:20
华北农学报(2020年1期)2020-04-16 03:37:26
蔬菜(2019年5期)2019-05-21 00:53:20
中华手工(2017年6期)2017-07-06 19:09:47
广西植物(2016年10期)2016-11-11 06:51:39
中华手工(2016年7期)2016-10-10 11:36:01
西南农业学报(2016年4期)2016-05-17 05:41:45
中国民族美术(2016年3期)2016-05-17 04:54:55
中华手工(2015年1期)2015-01-23 14:04:02
浙江农业科学(2013年6期)2013-11-08 03:40:40
