
杉阔混交和林下套种对土壤氮矿化的影响
2024-11-03 00:00:00高畅付若仙沐炜杰陈良喜吴望榕俞元春
林业科学研究 2024年5期




摘要:[目的]与阔叶树混交是提升杉木人工林土壤肥力的重要措施,探究不同杉阔混交模式下不同深度土壤氮矿化特征,对杉木人工林土壤养分循环的研究具有重要意义。[方法]选取两种经典混交模式的林分(杉-阔套种林、杉-阔混交林)与杉木纯林为研究对象,测定不同深度土壤(0~5 cm、5~20 cm、20~40 cm、40~60 cm)的理化性质、有效氮含量及氮矿化速率。[结果]0~5 cm土层的净氮矿化速率显著高于其他土层。混交和套种显著提高了0~5 cm土壤有效氮含量与净氮矿化速率。与杉木纯林相比,杉-阔套种林的土壤净氮矿化速率增加了111.19%,杉-阔混交林的土壤净氮矿化速率增加了70.73%。RDA分析表明:总碳和总氮含量是影响杉木人工林土壤氮矿化速率的主要因素,土壤含水率、碳氮比、可溶性有机碳和可溶性有机氮等对土壤氮矿化速率也有影响。相关性分析表明氮矿化速率最显著的0~5 cm土壤的碳氮比是净氮矿化速率的重要影响因素,而土壤理化性质在不同土层间对氮矿化的影响不同。[结论]土壤氮矿化与土壤深度密切相关,其主要发生在浅表层土壤且受多种土壤性质影响。杉阔混交和套种可以促进土壤氮矿化,对土壤氮矿化过程的正向影响利于维持和提升杉木人工林地的土壤肥力。
关键词:杉木人工林;杉阔混交;套种;土壤氮矿化
中图分类号:S714.6 文献标识码:A 文章编号:1001-1498(2024)05-0023-10
氮素是森林土壤营养元素最重要的组分之一。土壤中95%以上的氮以有机氮的形式存在,但有机氮只有经过氨化和硝化等矿化作用转化为无机氮才能被植物吸收利用。土壤氮矿化过程是土壤—植物养分循环过程中的重要环节。氮矿化速率是评估土壤有效氮供应能力的关键指标,研究森林土壤的氮矿化速率及其影响因素对了解土壤有效氮供应有重要意义。土壤氮矿化过程主要是由微生物主导的生物化学过程,微生物对生存环境的敏感性介导了氮矿化过程对土壤环境变化的反馈。有研究结果表明土壤氮矿化过程受环境因素的影响较大,如pH、温度、湿度、其余土壤理化性质等。并且这些因素常与土壤深度密切相关,土壤垂直尺度上的差别预计会导致氮矿化过程在不同土层中产生差异。
杉木(Cunninghamia lanceolate(Lamb.Hook))是我国重要的用材树种。但纯林连栽模式会导致土壤肥力下降、生产力和生物多样性降低等生态问题。近年来,为缓解纯林造林模式带来的不利影响,杉木与阔叶树种混交的造林模式被广泛认可。除营造同龄混交林外,在杉木纯林下套种阔叶树来营造异龄混交林也是一种混交方式,这样既能调整林分结构还能利用林分内剩余空间来提升林业生产资源的多样性。大量研究证明混交可以提升杉木人工林土壤氮养分。不同混交模式会对土壤的氮分布和转化产生不同影响,不同树种混交通过凋落物、根系分泌物和自身生长对土壤理化性质及微生物群落的调控来影响土壤氮矿化过程。但目前混交对杉木人工林土壤氮矿化影响的方向及程度并未有统一结论,目前的研究中也较少讨论混交林土壤氮矿化的垂直异质性,且土壤理化性质对不同深度土壤氮矿化的相对重要性仍未可知。为此,选取两种典型杉阔混交模式的林分与杉木纯林作为研究对象,对林下不同深度土壤(0~5 cm、5~20 cm、20~40 cm、40~60cm)的氮矿化速率、有效氮和理化性质进行测定,来探讨混交和套种对杉木人工林土壤氮矿化的影响,为维持和提升杉木人工林土壤肥力提供参考。
1 材料与方法
1.1 研究区概况
研究区域位于福建省南平市葫芦山国有林场(27°17'N,118°01'E)。该地区属南亚热带海洋性季风型气候,光照充足,气候温和,雨水充沛,年平均气温在17~19℃,年均降雨量和年均蒸发量分别为1669 mm和1413 mm,年均相对湿度81%。葫芦山林场土壤以红壤为主,土层厚度在100 cm以上。林下植被主要有芒萁(Dicranopterisdichotoma (Thunb.) Bernh)、五节芒(Miscanthusfloridulus (Lab.) Warb.)、鹧鸪草(Eriachnepallescens R.Br.)、狗脊(Woodwardia japonica(L.F.)Sm.)、黄瑞木(Adinandra millettii)等。
1.2 试验设计与样品采集
选择杉木纯林及两种典型营造杉阔混交林模式的林分为研究对象:第一种是将杉木和阔叶树种同期混合种植(杉-阔混交);第二种是对杉木纯林进行间伐后补植阔叶树种(杉-阔套种)。本文中杉-阔混交林中的阔叶树为米老排,学名壳菜果(Mytilaria laosensis Lecomte),该树种萌芽力强并且喜温喜热,是南方常见的优势阔叶树种。杉-阔套种林中的阔叶树为闽楠(Phoebe bournei(Hemsl.) Yen C.Yang),是珍贵用材树种且耐阴,适用于杉-阔套种林的营造。本研究选取的林分均为同批种植的二代杉木林,现为林龄≥30 a的成熟林。3种林分的初值密度均为2687~2985株·hm-2,纯林及杉-阔混交林经历两次间伐,杉-阔套种林经历3次间伐,每次间伐强度25%~50%。不同林分间的地理位置邻近、立地条件相似。林分基本情况信息详见表1。
本研究于2022年9月进行取样,在杉木纯林、杉-阔套种林、杉-阔混交林内分别设置3个20 m×20 m的样地,每个样地内按照“S”型采样法随机选取5个采样点用土钻取样,分别采集了0~5 cm、5~20 cm、20~40 cm、40~60 cm的土壤,并将同一样地内的土壤样品分土层混合均匀。将土壤样品置于放有冰袋的保温箱内运送至实验室。去除可见根、石砾和动物残体并过2 mm筛后,部分放入密封袋于4℃冰箱保存,部分放人晾土室风干,用于后续理化性质分析及培养实验。
1.3 指标测定
1.3.1 土壤理化性质
烘干法测量土壤含水率(SW);电极法测定pH值(水土比2.5:1);用1 mol·L-1的KCL溶液浸提新鲜土样后用连续流动分析仪(Skalar San++,荷兰)测定铵态氮(NH4+-N)、硝态氮(NO3--N);可溶性有机碳、氮(DOC、DON)用0.5 mol·L-1的K2SO4溶液浸提新鲜土样并离心后过0.45 μm孔径的滤膜,用TOC-VCPN自动分析仪(岛津,日本)测定;氯仿熏蒸-K2SO4浸提法测定新鲜土壤微生物生物量碳、氮(MBC、MBN);将风干后的土壤过0.149 mm筛后用元素分析仪(Elemental ELMAX CNS analyzer,德国)测量土壤内的总氮(TN)、总碳(TC)含量。
1.3.2 土壤氮矿化速率
土壤氮矿化速率采取好气培养法培养,取相当于10 g干土质量的新鲜土壤置于培养瓶中,调节含水量为田间最大持水量的60%。用透气的无菌封口膜封口,在25℃的培养箱中培养28 d,期间每48 h添加去离子水以维持土壤含水量。培养完成后立即测定样品内铵态氮和硝态氮的含量。根据培养前后铵态氮和硝态氮含量的差值计算净氮矿化速率(Nmin)、净氨化速率(Namm)和净硝化速率(Nnit)。如氮矿化速率为正值代表土壤有机氮释放,为负值则代表铵态氮和硝态氮被固定或消耗。计算公式如下(单位为mg·kg-l·d-l):
Nmin=(CAN-a+CNN-a-CAN-b-CNN-b)/t
Namm=(CAN-a-CAN-b)/t
Nnit=(CNN-a-CNN-b)/t
式中,CAN-a代表培养后铵态氮含量,CNN-a代表培养后硝态氮含量,CAN-b代表培养前铵态氮含量,CNN-b代表培养前硝态氮含量,t代表时间/d。
1.4 数据处理与分析
用Excel 2016对数据进行整理及计算,SPSS 26软件对数据进行单因素方差分析(One-way ANOVA)和Duncan多重比较分析不同林分类型、不同深度土层下各变量的差异性(p<0.05),利用Pearson相关分析对各指标进行相关性分析,采用R语言中的“vegan”包对土壤氮矿化速率和理化性质进行冗余分析(redundancyanalysis,RDA)。使用Origin2023b软件绘图。
2 结果与分析
2.1 杉阔套种、混交对不同深度土壤理化性质的影响
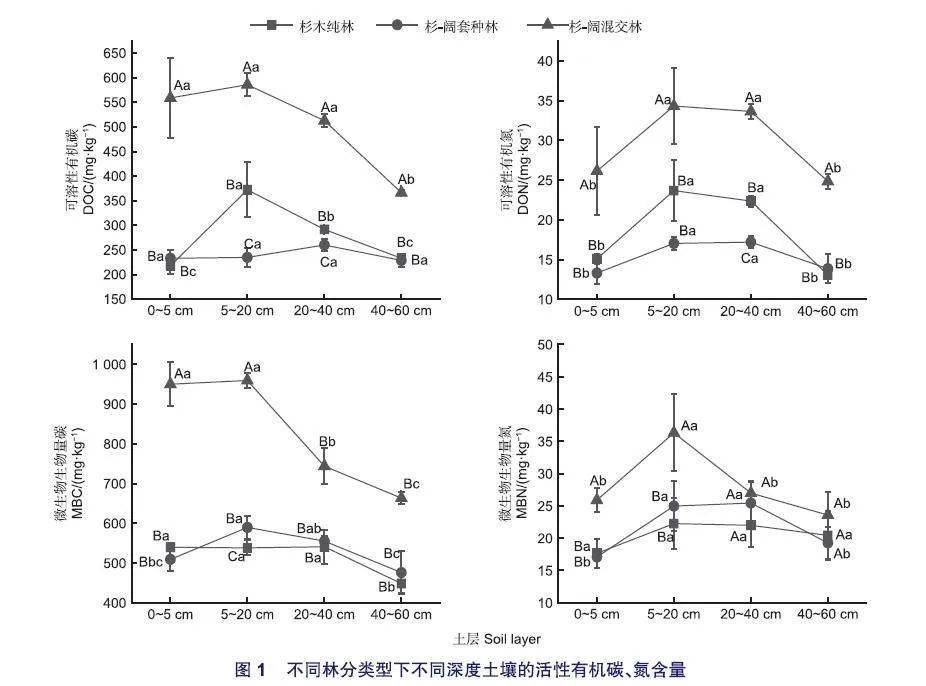
相比于纯林,杉-阔混交林的土壤pH值在0~40 cm土层内均显著降低(p<0.01),杉-阔套种林的土壤pH在0~5 cm土层显著低于杉木纯林(p<0.01)。杉-阔混交林整体土层的含水量与纯林相较更高,杉-阔套种林在0~20 cm土壤深度内的含水率(SW)显著低于纯林(p<0.01)。对比纯林,杉-阔混交林中四个土层的总碳(TC)、总氮(TN)含量均显著增加(p<0.01)。杉-阔套种林的TC含量对比纯林在0~5 cm土层显著降低,在其余土层均显著增加(p<0.01),TN含量在5~20 cm和20~40 cm土层中显著增加(p<0.01)。杉-阔混交林和杉-阔套种林相比杉木纯林在0~5 cm土层的土壤碳氮比显著降低(p<0.01),但在其它土层均显著增加(p<0.01)(表2)。
图1表明,杉-阔套种林的土壤可溶性有机碳、氮(DOC、DON)含量对比纯林在5~20 cm、20~40 cm土层均显著降低(p<0.01),而杉-阔混交林的DOC、DON含量在各土层均显著增加(p<0.05)。杉-阔混交林的土壤微生物生物量碳、氮(MBC、MBN)含量在全土层中均高于杉木纯林,在0~5 cm、5~20 cm层差异显著(p<0.01)。杉-阔套种林的MBC含量在5~20 cm土层与杉木纯林相比显著增加(p<0.01)。
2.2 杉阔混交、套种对不同深度土壤有效氮的影响
如图2所示,相比于杉木纯林,杉-阔套种林0~5 cm和5~20 cm土层的硝态氮(NO3--N)含量分别提高了74.71%和186.39%(p<0.01)。杉-阔套种林的土壤铵态氮(NH4、-N)含量在20~40 cm层显著高于纯林64.09% (p<0.01),杉-阔混交林的NH4、-N含量在0~5 cm、20~40 cm土层中对比纯林增加了114.82%和56.83%(p<0.01)。3种林分类型的NO3--N含量在0~5 cm土层均显著高于其它3层(p<0.01),NH4、-N含量随土层深度增加而减少。
2.3 杉阔混交、套种对不同深度土壤氮矿化速率的影响
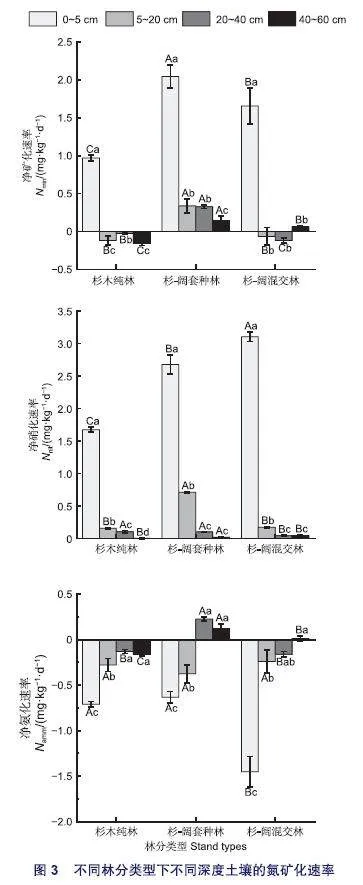
2.3.1 净氮矿化速率
3种林分内的土壤净矿化速率在土壤深度层面上均发生了显著变异(p<0.01)。由图3知,所有林分类型下土壤氮矿化过程均主要发生在0~5 cm层的土壤中。在净氮矿化速率最高的0~5 cm层中,与杉木纯林相比,杉-阔套种林的土壤净矿化速率增加了111.19% (p<0.01),杉-阔混交林的土壤净氮矿化速率增加了70.73%(p<0.01)。在杉-阔套种林中,其余土层的净氮矿化速率相比纯林均显著增加(p<0.01)。在杉-阔混交林中20~40 cm土层的净矿化速率相比纯林显著降低(p<0.01),而40~60 cm的净矿化速率显著增加(p<0.01)。
2.3.2 净硝化速率
所有林分类型中土壤净硝化速率均为0~5 cm层的土壤中最高,其它土层间的土壤净硝化速率也存在显著差异但与0~5 cm层相较数值较小(p<0.01)。在净硝化速率最高的0~5 cm层中,与杉木纯林相比,杉-阔套种林的土壤净硝化速率增加了59.92%(p<0.01),杉-阔混交林增加了83.4%(p<0.01)。在净硝化速率较低的土层中,杉-阔套种林在5~20 cm层相比纯林显著增加(p<0.01)。
2.3.3 净铵化速率
在3种林分类型中,大部分土层的净氨化速率整体为负值,且土壤净氨化速率均在0~5 cm最低(p<0.01),随着土壤深度的增加大致呈现出增加的趋势。在0~5 cm土层中,杉-阔混交林中的土壤净氨化速率显著低于杉木纯林(p<0.01)。杉-阔套种林在20~60 cm土层的土壤净氨化速率均显著高于纯林(p<0.01)。
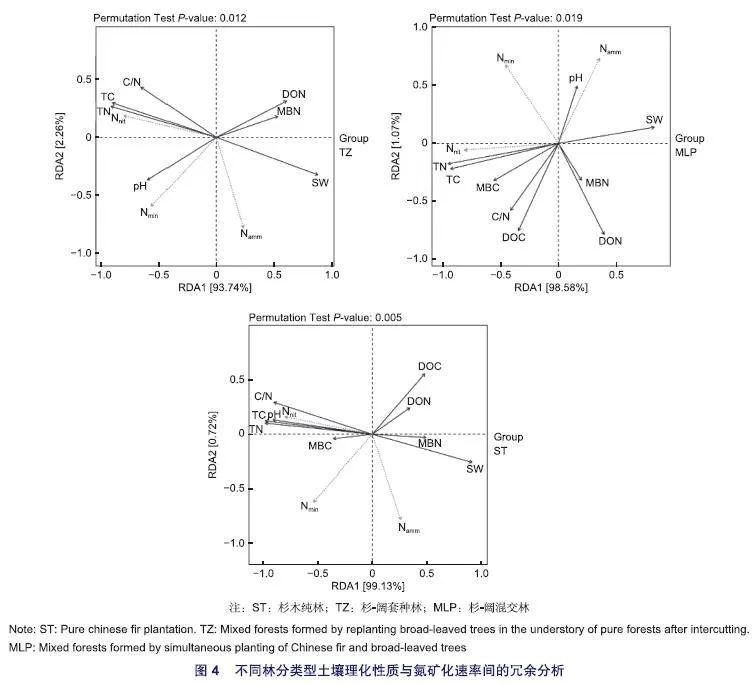
2.4 不同林分类型下土壤环境因子对氮矿化速率的影响
为了探究土壤环境因子对土壤氮矿化速率的解释率和排序特征进行了冗余分析(图4)。结果表明,3种林分类型中的土壤环境因子对氮矿化速率的总解释率均达95%以上。3种林分类型的土壤TC、TN与净氮矿化速率具有紧密的正相关关系,且对氮矿化速率具有最高的解释率,是影响土壤氮矿化的最重要因素。净氮矿化速率与SW呈负相关而与pH呈正相关。在杉-阔套种林与杉木纯林中C/N与净氮矿化速率具有较强的正相关。DOC在杉木纯林、杉-阔混交林中与净氮矿化速率存在负相关。DON在杉-阔套种林、杉-阔混交林中与净氮矿化速率存在负相关。MBC、MBN对3种类型林分土壤的氮矿化速率也有不同影响但影响程度较低。土壤环境因子与净硝化速率的关系大致同净氮矿化速率一致,而与净氨化速率则多数相反。
土壤理化性质在不同土层间对氮矿化具有不同影响(表3)。净氮矿化速率在0~5 cm土层与C/N呈显著负相关;在5~20 cm土层与DOC、DON及SW呈显著负相关;在20~40 cm土层与MBC、DOC、DON呈显著负相关,与pH呈显著正相关;在40~60 cm土层与C/N呈显著正相关。净氨化速率与净硝化速率在不同土层与理化性质的相关性也存在差异。
3 讨论
3.1 杉阔混交、套种对不同深度土壤有效氮含量的影响
土壤中的无机氮是能被植物直接吸收利用的氮,其含量是评价土壤供氮能力的重要指标,通常将土壤内的铵态氮与硝态氮视为土壤内主要的有效氮。本研究中3种林分的铵态氮、硝态氮含量均随土层加深而减少,这是因为凋落物的回归发生在土壤表层。一般来说,土壤氮素最主要的来源是凋落物,并且通常表层土壤的环境更利于微生物驱动的一系列养分循环活动。
不同林分类型主要通过凋落物、根系的输入与分解,以及地上植物的吸收利用和微生物对氮循环过程的调控来影响土壤中的有效氮含量。杉-阔套种提高了0~5 cm与5~20 cm土壤的硝态氮含量,这是由于显著提升的硝化速率促进了更多硝态氮的产生,同时低含水率阻碍了植物根系的对硝态氮的吸收消耗。杉-阔混交林的净硝化速率在多土层中显著高于纯林,但未发现土壤硝态氮显著累积,这可能是因为米老排的速生习性导致对硝态氮的吸收利用需求较大。与杉木纯林相比,杉-阔套种和杉-阔混交林20~40 cm土层中的铵态氮含量显著增加。除去混交效应带来的更多养分输入,杉木树种本身对于铵态氮具有明显的吸收偏好,因此相比混交林,杉木纯林可能加剧了铵态氮的消耗损失。
3.2 杉阔混交、套种对不同深度土壤氮矿化特征的影响
本研究表明土壤氮矿化过程中硝化作用对净氮矿化速率的贡献率更高,而净氨化速率多数为负数,这与许多研究结果一致,可能是因为氨化作用导致的铵态氮累积为硝化作用提供了反应底物。有研究表明,在微生物对无机氮的固持作用中铵态氮被明显偏爱,而当微生物分解有机氮所产生的铵态氮超过了植物和微生物所吸收的量时,铵态氮就会转变为硝态氮。且据报道亚热带酸性红壤中的异养硝化作用更强,且主导自养硝化作用的微生物在酸性红壤环境下适应良好。本研究结果显示3种林分类型的土壤净氮矿化速率均在0~5 cm土层最高,说明土壤氮矿化作用主要发生在浅表层。首先凋落物的积累和分解主要在土壤的浅表层发生,其释放的有机质既包含氮矿化作用的底物,又是微生物生命活动的能量来源。本研究中,净氮矿化速率与土壤中的总碳、总氮有显著相关性,且其含量随着土层深度显著降低,这表明有机质更丰富是0~5 cm土层矿化速率高的重要原因。其次,本研究显示净矿化速率与土壤含水量呈负相关,表层土壤相比下部土壤具有更好的透水透气性,这有利于微生物在表层聚集并保持活性进而对土壤氮矿化过程有促进作用。
在矿化作用最为显著的浅表层土壤中,杉-阔套种林和杉-阔混交林的净矿化速率与净硝化速率均显著高于杉木纯林,这证实了营造混交林会促进杉木纯林的土壤氮矿化作用。苏波等人对针叶林与针阔混交林的研究中同样认为混交经营提升了氮矿化作用和土壤供氮能力,并发现这种差异是由林下微环境以及土壤环境因子变化所导致的。首先,混交阔叶树种通常会增加凋落物的数量和质量,而凋落物是森林将养分回归给土壤的最主要来源。在本研究中,杉-阔混交林的土壤总碳、总氮的含量均显著高于杉木纯林,凋落物分解后所带来的土壤有机质增加有利于混交林土壤氮矿化速率的提高。其次,利用相关分析发现0~5 cm土层土壤的净氮矿化速率与碳氮比呈显著负相关。通常认为在一定范围内,土壤碳氮比低代表着有机质更易被分解,也就是有机质的矿化速率更快。由于杉木的凋落物含有较多的木质素、纤维素、单宁等物质,不利于微生物的分解利用,而阔叶树凋落物木质素、纤维素含量较少更易被微生物分解。所以0~5 cm土层碳氮比低也是造成套种和混交林的净氮矿化速率高于杉木纯林的原因之一。
氮转化速率取决于土壤中微生物活性的高低。本研究中显示0~5 cm和5~20 cm土层的土壤氮矿化速率与微生物量碳、氮含量的相关性较弱,这有可能是因为混交效应更多的是激发了微生物的活性而不是增加微生物数量。Holems等研究也表明,相比于土壤微生物量,微生物的活性似乎是导致土壤净氮矿化在不同季节变异的更重要因素。在本研究中,可溶性有机碳、氮在不同林分及土层中与净氮矿化速率呈负相关,代表微生物进行矿化作用时会消耗活性碳氮。相比于杉木纯林,杉-阔套种林中能够被微生物快速利用的可溶性有机碳、氮显著降低而微生物量未显著增加,表明微生物对活性碳源的消耗更多地用于维持生产活动而不是繁殖,这成为混交效应下有机氮矿化提升的潜在原因。
4 结论
杉阔混交和林下套种可以促进杉木人工林土壤氮矿化过程,土壤总碳和总氮的含量升高是导致混交林的土壤氮矿化速率高于纯林的重要因素。不同土壤深度的杉木人工林土壤氮矿化差异显著,0~5 cm土层土壤氮矿化速率显著高于下层土壤,混交和套种显著降低了0~5 cm土层的碳氮比进而促进了土壤氮矿化。混交和套种对土壤氮矿化的正向影响保证了土壤对植物供应更多的有效氮,为维持和提升杉木人工林土壤肥力和林分生产力提供养分保障。
(责任编辑:崔贝)
基金项目:国家“十四五”重点研发计划项目(2021YFD2201304);江苏高校优势学科建设工程资助项目(PAPD)
