
基于鸟类多样性保护的武汉都市圈生态网络构建探索
2024-10-11 00:00:00郭诗怡田雪刘媛李丹哲贺慧
上海城市规划 2024年4期
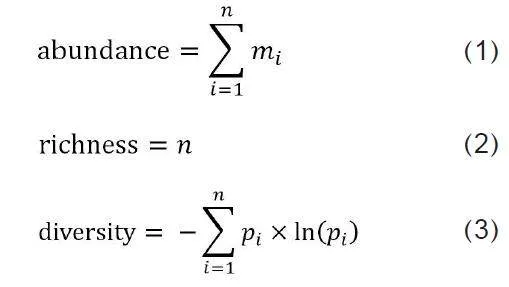
摘 要 生态网络有助于提高区域景观连通性,增加斑块之间的物质能量和基因交流,对生物多样性保护具有重要意义。既往研究侧重于从经验主义或基于图论等理论探索景观模型构建方法,较少针对具体物种的长时序观测数据建立“物种—环境”相关性模型从而构建生态网络。随着《武汉都市圈发展规划》获国家发改委正式批复,武汉都市圈成为第7个获批的国家级都市圈。以武汉都市圈为例,基于长达15年(2007—2021年)的鸟类长期观测数据和城乡环境数据,探究城乡环境特征对鸟类多样性的影响机制,识别重要鸟类栖息地的空间分布,构建以鸟类为指征的生态源地—生态廊道—生态网络保护体系。此外,基于城市扩张模型分析生态网络被侵占风险,对其进行重要性和敏感性分级,识别鸟类多样性保护的难点和痛点。有望为面向鸟类多样性保护的都市圈层面生态网络建设提供实证支撑,为国土空间规划中面向生物多样性保护的都市圈层面重要生态空间划定与生态修复提供理论参考。
关 键 词 鸟类多样性;生态网络;公众科学;城市扩张;武汉都市圈
文章编号 1673-8985(2024)04-0123-09 中图分类号 TU984 文献标志码 A DOI 10.11982/j.supr.20240417
生物多样性保护关乎生态系统的韧性和人类福祉[1-2],它是指来自陆地、海洋和其他水生生态系统及其所属生态复合体的生物体间的多样性,包括物种内部、物种之间和生态系统的多样性[3-4]。世界自然基金会(WWF)发布的《地球生命力报告2020》指出,1970—2016年间全球野生动物种群数量下降了68%,且22%的植物有灭绝风险。生物多样性丧失将严重影响生态过程和生态系统的韧性,阻碍其在调节气候、降低污染、授粉、固碳等方面发挥关键作用,进而对人类获得食物、纤维、水、能源、药物和其他遗传物质造成威胁[5]。生物多样性丧失已不单是环境问题,更关乎人类福祉和全球安全。
城市中常见的野生动物——鸟类,是目前国际公认指示生物多样性的物种[6],它们对于城市环境变化的响应具有典型性与代表性。一方面由于鸟类相对容易识别和获取,鸟类生态学发源较早,学界已具备相对丰富的鸟类相关信息积累[7];另一方面由于鸟类是高度可移动的物种,对小尺度的栖息地环境与大尺度的区域环境变化均有响应。尽管城市为鸟类提供了水源、食物来源和居所,城市建设中对绿地与水体的破坏压缩了鸟类的生存空间,对鸟类多样性产生了巨大威胁。
随着《武汉都市圈发展规划》获国家发改委正式批复,武汉都市圈成为第7个获批的国家级都市圈。江河密布、湿地众多的武汉都市圈位于全球重要候鸟迁徙路线之一的“东亚—澳大利亚候鸟迁徙路线”上,既是候鸟迁徙的重要中转站和越冬区,也是青头潜鸭等珍稀鸟类的繁殖地。因此,作为我国中部最具影响力的城市发展集群,有必要厘清武汉都市圈城乡环境变化对鸟类多样性的影响,科学划定重要生态源地和廊道并构建生态网络,为面向鸟类多样性保护的都市圈层面生态网络构建树立典范。
1 相关研究
从基于景观生态学模型的生态网络规划,到制定生物多样性保护的专项计划并出台相关支撑文件,最后落到生境修复和营造具体措施以确保城市生物多样性保护规划的落地实施——城市生物多样性保护的规划研究正逐步精细化[8]。然而,具体物种保护规划研究进展相对缓慢。一方面由于物种保护规划需要物种观测和田野调查作为支撑,比较耗费人力物力。另一方面,城市建成环境与鸟类多样性的关联性研究向实践应用的转化较少;既有研究多采用经验主义,依据物种分布记录,以区域内物种特有性和受威胁程度确定保护措施,或采用模型估计区域内生物多样性,包括植被覆盖率、景观斑块连接度、综合物种多样性指数等指标[9],但上述方法大多围绕栖息地或生境质量的提升,而较少从城市乃至城市群层面探索面向鸟类多样性保护的重要栖息地判别及保护方法。
随着城市化的快速推进,基于鸟类对城市生态环境和居民福祉的重要性开展城市背景下的生物多样性保护研究显得尤为重要,我国针对鸟类多样性影响机制的研究仍主要在自然栖息地开展,以不同生境鸟类群落结构及其动态变化为主[10],而从城市建成环境入手,结合地域代表性鸟类的生态学习性和栖息地偏好对鸟类多样性的时空影响机制的研究则不足。城市建成环境对鸟类多样性的影响机制是识别重要鸟类栖息地的关键切入点。生物多样性研究视角下,城市建成环境包括人工环境(如建筑、道路、车辆、人流、噪声、污染物)和自然基底(如林地、水体、草地、湿地等)两部分[11]。既往研究多从栖息地尺度和区域/景观尺度分别探讨城市建成环境对鸟类多样性的影响[12]。栖息地尺度下往往关注小气候(热环境、风速、温度)、水(含沙量、PH值、微生物含量)、植物(与动物多样性相关)、噪声水平等微观环境或当地特有的环境特征。区域/景观尺度的显著环境要素可分为:(1)土地覆盖,即绿地面积、水域面积、不透水表面面积等[13-14]到特定类型区域的距离,如距离城市中心的距离、距离绿地的距离、距离水体的距离等[15]。(2)景观生态学指标,即景观连接度、斑块密度等[16-18]。这一尺度下既往研究较多关注城市灰绿空间比例及景观格局,仅有个别研究探讨了城市社会要素对生物多样性的影响,包括土地利用变化、人类活动等[19-20]。
公众科学(citizen science)通常指公众参与科学研究并产生新知识的有意识的合作过程,其参与者是不分地域的非专家群体[21],通过借助大量来自普通公众的非职业科学家、科学爱好者或志愿者,进行数据采集、分析、应用等活动。公众科学形式的鸟类调查与传统鸟类调查相比,扩大了调查和评估的范围,增强了数据采集能力,为鸟类研究提供了大量数据,弥补了传统调查的不足,能有效完善生物多样性本底资料,或可为区域视角下的城市鸟类多样性时空影响机制提供支撑[22-23]。
生态网络起源于岛屿生物地理理论,指在空间中利用线性廊道将破碎的景观斑块进行有机联系,维持区域内生物多样性、景观完整性的网络体系[24]。当前生态网络构建研究侧重于利用景观生态学相关模型对生物栖息地和迁徙廊道进行定量分析,包括采用描述要素或区域景观总体特征的景观格局指数法[25-26],以及遵循“生态源地识别—综合阻力面构建—生态网络生成”的基本模式的图论算法、电路理论、最小累计阻力模型、斑块重力模型等具体模型手段构建区域生态网络[27]。其中最小累计阻力模型通过计算物种在不同生态源地间运动所需要耗费的代价,反映物种运动的潜在可能性及趋势,能较好地模拟较大尺度内景观要素间的相互关系,被广泛应用于生态网络构建与优化研究与实践中[28-29]。
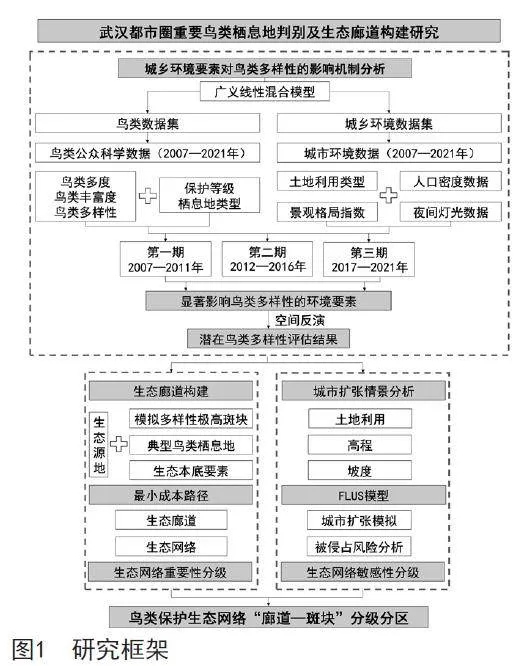
本研究以武汉都市圈为研究对象,根据2007—2021年鸟类长期观测数据和2009、2014、2019年城乡环境数据,识别鸟类多样性和城乡环境演变特征,探究武汉都市圈城乡环境特征对鸟类多样性的影响机制,评估武汉都市圈潜在鸟类多样性,利用最小累计阻力模型构建由鸟类栖息地和鸟类迁移廊道组成的生态网络,进而以城市扩张对鸟类栖息地和生态廊道的侵占风险为依据,对生态网络进行重要性和敏感性分级,识别鸟类多样性保护的难点和痛点,为武汉都市圈生态网络建设提供科学依据,为面向生物多样性保护的国土空间规划和重要生态空间保护与修复工程提供参考,研究框架如图1所示。
2 研究区概况
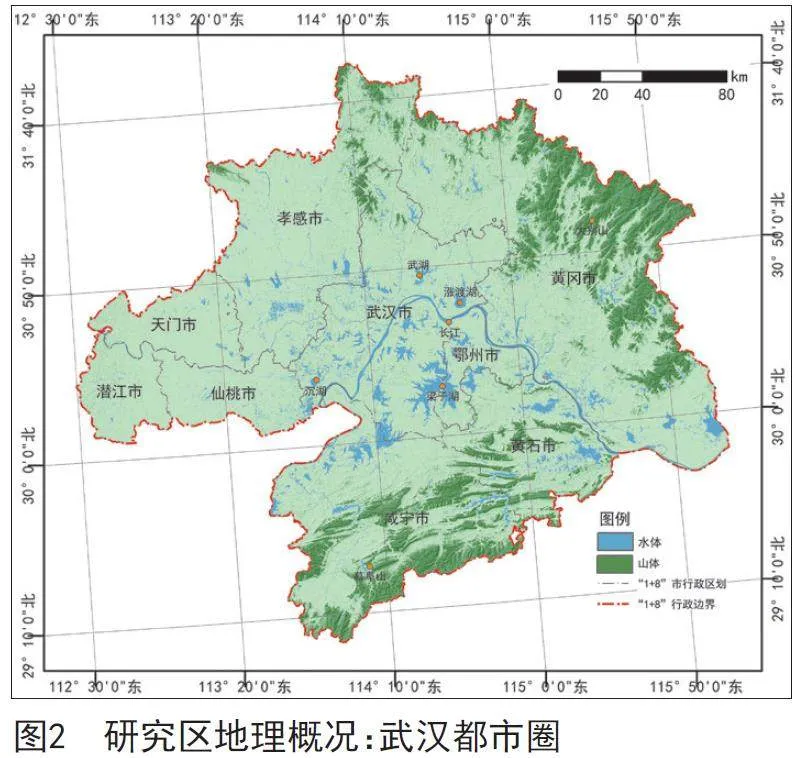
武汉都市圈(见图2)是以我国中部最大城市武汉为中心,覆盖周边100 km范围内的8个大中型城市(黄冈、鄂州、黄石、孝感、咸宁、仙桃、天门、潜江)而构成的城市联合体,是湖北省经济要素最为密集、最具活力的地区。武汉都市圈生态本底条件优越,生态要素齐全,生境类型丰富,分布有山脉、森林、河流、湖泊、草原等多种土地覆盖类型。南北环绕有以幕阜山、大别山余脉为主的低山丘陵,中部分布有梁子湖、汤逊湖、鲁湖、东湖、沉湖等重要湖泊,形成了以山脉、水系为骨干,以山、林、江、湖为基本要素的多层次、多功能、复合型、网络化的区域生态体系。武汉都市圈鸟类资源丰富,目前已观测到的鸟类共417种,占全国鸟类总数的28%。其中,国家一级保护鸟类12种,如东方白鹳、黑鹳等;国家二级保护鸟类67种,如白额雁、鸳鸯、灰鹤等。
3 研究方法与数据
3.1 数据来源与预处理
3.1.1 鸟类分布数据
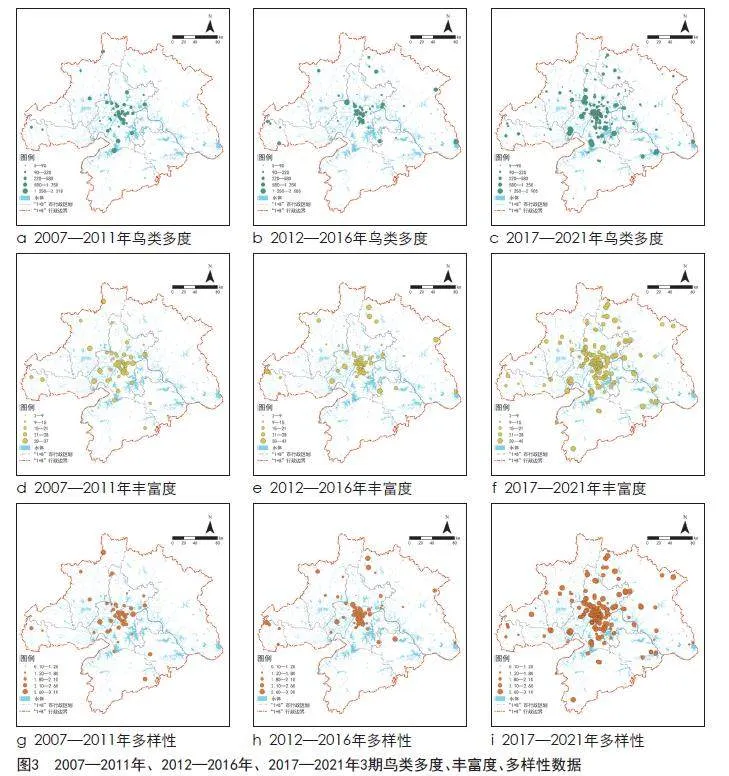
从“中国观鸟记录中心”网站(见表1)检索2007—2021年武汉都市圈的观鸟记录共5 831条,观鸟报告内信息包括鸟类名称、种、目、科、数量、观测地点、观测时间等。筛选2007—2011年、2012—2016年、2017—2021年3期内每年重复观测次数≥3次的观鸟记录,分别得到鸟类观测点62个、95个、308个,观测到386种、108 410只鸟类。分别计算3期鸟类观测点的鸟类多度(abundance)、丰富度(richness)、多样性(diversity)3类指标(见图3)。
式中:n表示每次观测中观测到的鸟类物种数;mi表示该次观测中第i个物种的个体数;pi表示第i个物种占该次观测到的物种总个体数的比例。
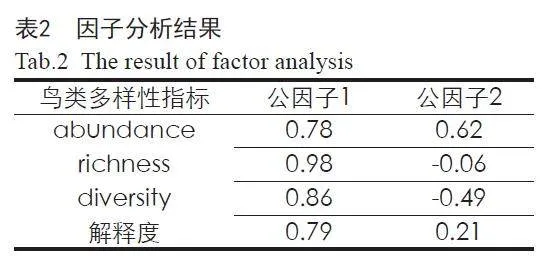
通过自然对数法将鸟类多度、鸟类丰富度、鸟类多样性3类指标进行归一化处理,通过R语言psych包采用因子分析法对3类指标进行降维处理。其中,fa函数进行主成分分析,经过正交旋转后得到武汉都市圈的总体鸟类多样性。因子分析结果如表2所示,公因子1对武汉都市圈总体鸟类多样性的解释度达0.79,满足本研究的精度需求。
3.1.2 城市环境数据
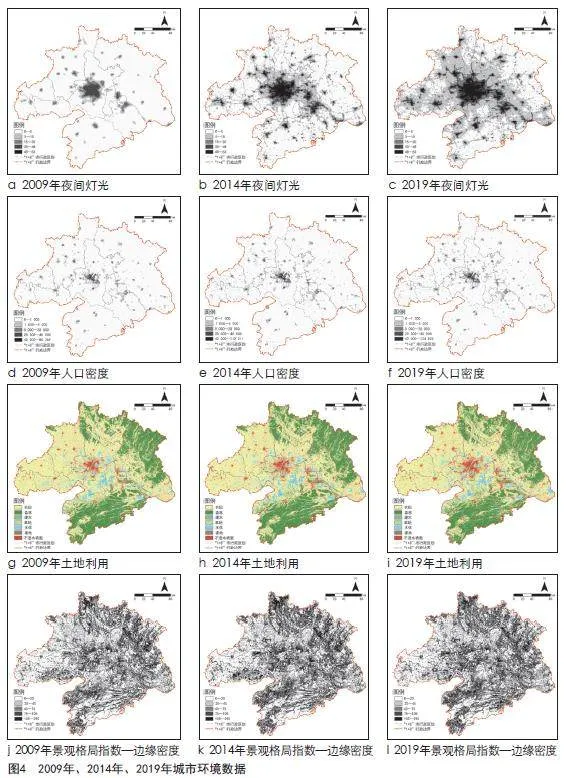
(1)土地利用数据:根据Zenodo数据知识库获取相应3期30 m精度的武汉都市圈土地利用数据,将武汉都市圈土地利用类型划分为农田、森林、灌木、草地、水体、裸地和不透水表面7类,得到武汉都市圈2009年、2014年、2019年的土地利用数据。
(2)夜间灯光数据:本研究选择2009年第4版的DMSP/OLS全球夜间灯光影像及2014年和2019年NPP/VIIRS全球夜间灯光影像,精度为1 000 m。由于DMSP/OLS全球夜间灯光影像和NPP/VIIRS全球夜间灯光影像分别由不同传感器检测完成,因此这3期数据需要进行预处理后才能使用。首先,为消除DMSP/OLS夜间灯光数据内部之间存在的差异,对DMSP/OLS夜间灯光数据进行逐步校正和饱和校正,并进行年内融合。然后,采用像元模拟法建立DMSP/OLS和NPP/VIIRS两种夜间灯光数据之间的相互校正模型,得到可比较的2009年、2014年、2019年的夜间灯光数据。
(3)人口密度数据:Worldpop人口密度数据是基于各国人口普查和官方人口估计数据,通过随机森林模型(Random Forest)生成的年度人口密度数据集,精度为1 000 m。根据武汉都市圈2009年、2014年和2019年人口统计年鉴,对该数据进行校正,得到可比较的武汉都市圈2009年、2014年、2019年的人口密度数据。
(4)景观格局指数:通过Fragstats软件,利用武汉都市圈2009年、2014年、2019年土地利用数据,提取森林、灌木、草地,评估武汉都市圈3期景观格局动态变化。本研究从景观破碎度、景观斑块形状和景观多样性3方面进行景观格局变化分析,选取斑块密度(PD)、最大斑块指数(LPI)、边缘密度(ED)、斑块形状指数(LSI)、聚集度指数(AI)、香农多样性指数(SHDI)、香农均匀度指数(SHEI)7种景观格局指数进行景观格局变化分析。
上述数据名称和来源见表1和图4。
3.2 研究方法
3.2.1 广义线性混合模型
首先通过共线性研判对自变量进行筛选,利用R语言Imer4程序包通过广义线性混合模型(Generalized Linear Mixed Model)探究武汉都市圈城乡环境特征对鸟类生物多样性的影响机制,并利用MuMin函数计算模型解释度,提取显著影响的环境要素和相关性系数,通过空间反演预测武汉市鸟类多样性空间分布。
3.2.2 最小累计阻力模型
最小累计阻力模型(Minimum Cumulative Resistance,MCR)通过模拟出发地与目标地之间的最短成本距离以确定物种迁移的最佳路径,是模拟生态廊道的常用方法之一。本研究利用反函数对潜在武汉都市圈生物多样性评估结果进行变换得到阻力面,通过ArcGIS中Spatial Analyst—距离分析—成本距离及成本路径工具生成最短成本路径并结合自然本底条件继续筛选,得到生态廊道,从而构建武汉都市圈生态网络。
3.2.3 FLUS模型
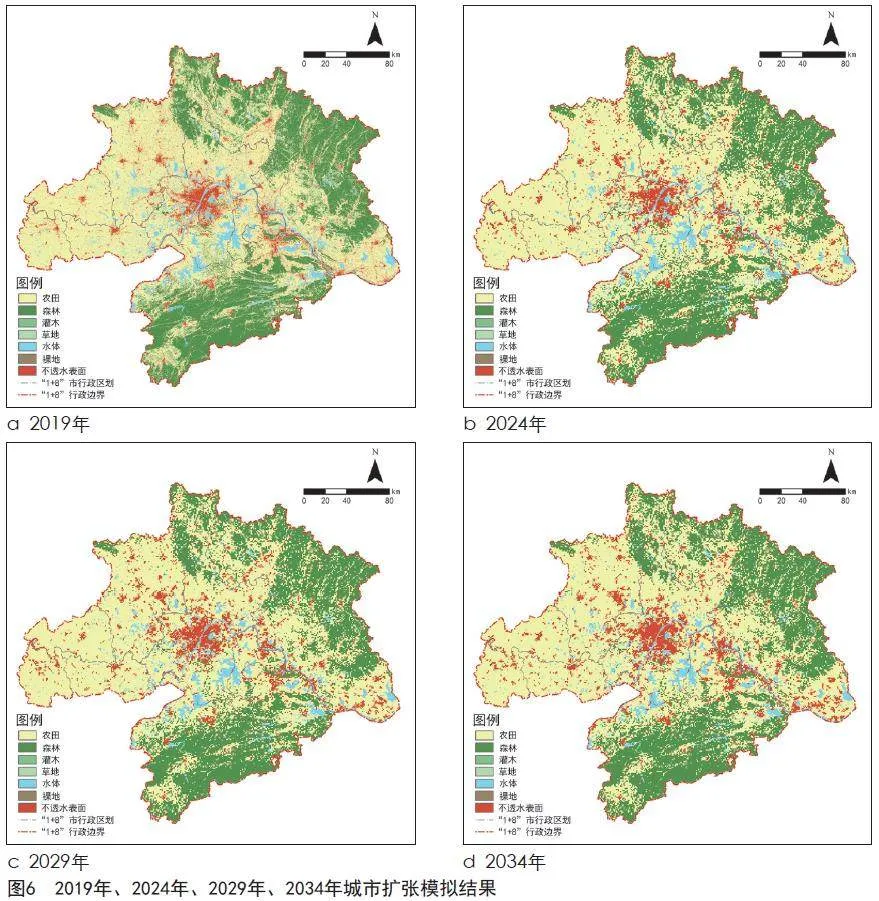
FLUS模型由基于神经网络的适宜性概率计算模块(BP-ANN)和基于自适应惯性机制的元胞自动机模块(CA)组成。BP-ANN模块需输入土地利用变化驱动因子,本研究通过武汉都市圈30 m分辨率DEM数据得到的高程和坡度作为决定土地利用的主要因素。设置神经网络训练采样比例(本研究设置为2%),并选用随机采样模式进行各类用地的训练样本采样(本文训练隐藏层数设置为12),实现神经网络训练。结合标准化处理后各驱动因子分布状况,计算得到各土地利用类型转换的适宜性概率结果。然后确定不同土地利用类型相互转换难易程度矩阵(0表示不允许转换,1表示可自由转换),预设各土地利用类型变化数量的目标,再通过自适应惯性机制的元胞自动机模块进行模拟,模拟迭代目标次数为300,邻域大小为3×3,最终实现武汉都市圈2024年、2029年、2034年城市扩张模拟结果[30]。
4 结果与分析
4.1 显著影响鸟类多样性的环境要素
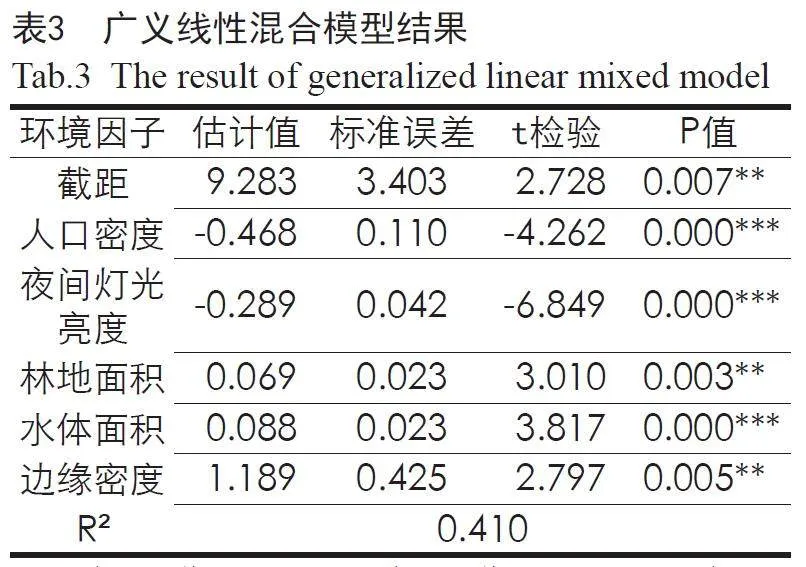
通过广义线性混合模型,构建“鸟类多样性—城乡环境要素”回归模型(见表3)。结果表明,人口密度和夜间灯光亮度与鸟类多样性呈显著负相关,而土地利用数据中的林地面积、水体面积和反映总体景观格局的边缘密度指数与鸟类多样性呈显著正相关,且人口密度、夜间灯光亮度对鸟类多样性的负面影响强于土地利用,其他城乡环境要素则对鸟类多样性的影响不明显。根据回归结果构建武汉都市圈潜在鸟类多样性分布模型,模型解释度达41%,模型预测结果精度较高。
4.2 重要鸟类栖息地判别和生态廊道构建
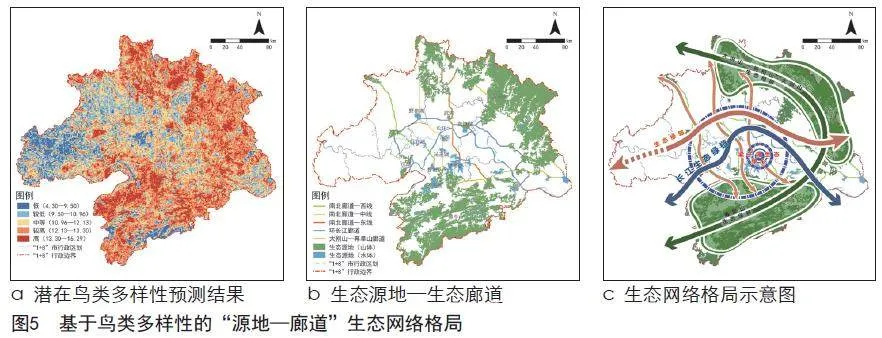
潜在鸟类多样性模拟结果(见图5)表明,在东北部大别山、南部幕阜山等山区,以及涨渡湖、沉湖等重要水体区域鸟类多样性水平较高,而在武汉市三环线以内等城市建设水平较高、人口密度较大的城市中心区鸟类多样性水平较低,整体呈现城区低郊区高的特点。
在ArcGIS中将潜在鸟类多样性模拟结果分级,并结合武汉都市圈“三区三线”划定及重要自然本底要素,提取26处重要鸟类栖息地作为生态源地,总面积18 841.52 km²,占研究区总面积的32.08%。通过最小成本路径模型,结合生态源地空间分布,筛选确定得到5条生态廊道,廊道总长度为85.33 km。
由生态源地及生态廊道构成的生态网络可整体概括为“一心一链两屏四轴”:梁子湖生态绿心是武汉都市圈内面积最大、鸟类多样性最高、最为完整的湿地斑块,以梁子湖为中心,沿长江展开的生态绿链和以南北两侧山林为主的两道生态屏障形成了武汉都市圈生态网络的基本骨架,再由跨越长江、联通南北的4条生态绿轴串联各湖泊、河流、山体等生态源地,构建了以保护生物多样性为导向的武汉都市圈生态网络安全格局。
4.3 基于FLUS模型的生境敏感性评价
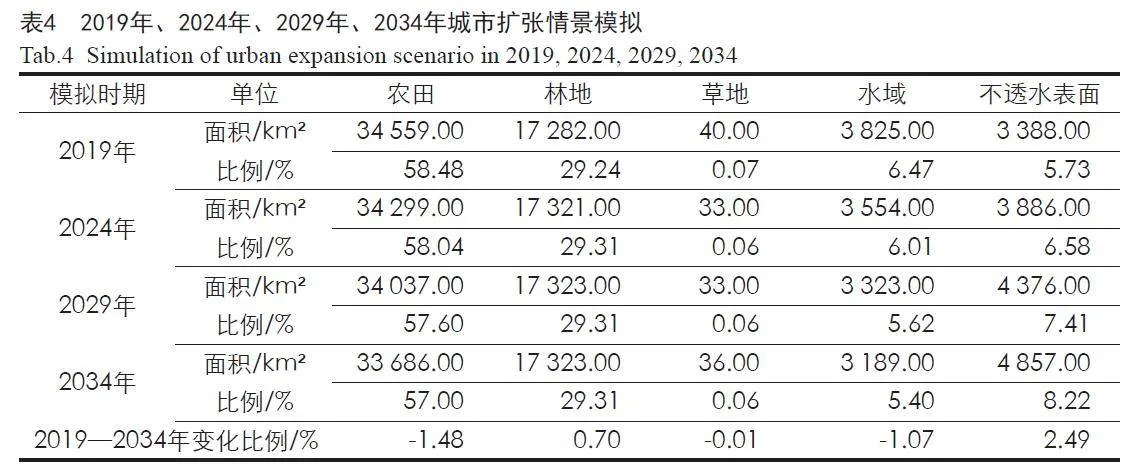
以2014年武汉都市圈土地利用数据为基础,利用FLUS模型,对2019年武汉都市圈土地利用进行模拟预测(见图6,表4),模拟精度Kappa系数达0.8,满足本研究的精度需求。在此基础上,以2019年土地利用数据分别对2024年、2029年、2034年土地利用变化进行预测。到2034年,武汉都市圈不透水表面的面积增幅最大,相比2019年增大了2.49%,城市化水平显著提高,林地面积预计增长0.7%,水体面积预计下降1.07%。这表明武汉都市圈生态保护措施有一定成效,但水体保护措施仍有待加强。
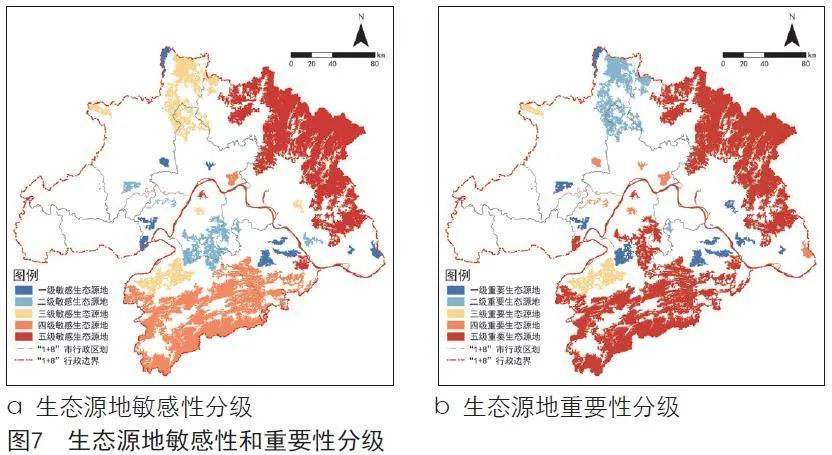
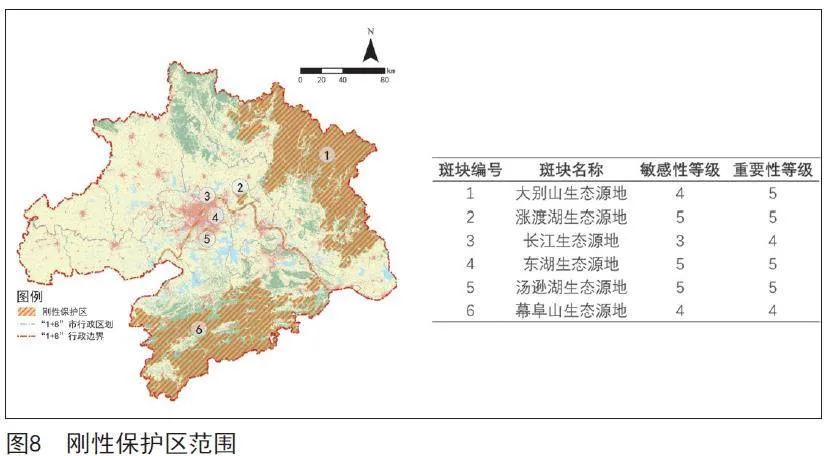
根据生态源地被侵占面积对比,对生态源地进行敏感性等级划分(见图7)。大别山等林地生态源地和长江、东湖等水体生态源地被侵占面积极大,划定为五级敏感生态源地,总面积达8 284.22 km²。幕阜山、涨渡湖区域被侵占面积较大,划定为四级敏感生态源地,总面积达6 014.48 km²。其余生态源地被侵占面积较少,因此划定为三级、二级、一级敏感生态源地,总面积依次为2 487.89 km²、1 226.08 km²、828.85 km²。
结合武汉都市圈常见林鸟、常见水鸟及珍稀鸟类在生态源地的空间分布,对生态源地进行重要性等级划分。幕阜山、大别山、长江、沉湖、梁子湖、东湖区域鸟类多样性评估结果极高,且有较多珍稀鸟类(黄胸鹀、东方白鹳、青头潜鸭、黑鹳、白鹤、白胸翡翠等)分布,划定为五级重要生态源地,总面积达14 991.00 km²。武湖、龙感湖、汤逊湖、后官湖、涨渡湖、野猪湖鸟类多样性评估结果较高,有一定珍稀鸟类分布,划定为四级重要生态源地,总面积达368.46 km²。其余生态源地鸟类多样性评估结果较低,同时源地自身面积较小,因此划定为三级、二级、一级重要生态源地,总面积依次为796.15 km²、1 633.02 km²、1 052.89 km²。幕阜山、大别山、长江、东湖、涨渡湖区域同时具有较高等级的重要性和敏感性,在后续规划及管理保护中需重点保护。
根据生态源地敏感性与重要性分级结果,提取敏感性与重要性分级均在四级及以上的生态源地斑块作为刚性保护区(见图8)。其中,大别山、幕阜山为武汉都市圈南北两侧山体屏障,涨渡湖、东湖、汤逊湖长期以来是武汉都市圈湿地鸟类的重要栖息地和越冬区,长江流域湿地更是世界生物多样性最丰富的区域之一,在确保上述区域鸟类多样性保护关键斑块得到有效保障的同时,对其余生态源地斑块进行弹性引导,构建刚性控制与弹性保护相结合的可持续发展的武汉都市圈生态网络格局。
5 结论与讨论
本研究以武汉都市圈为研究对象,基于长达15年的鸟类长时序观测数据和多源城乡环境数据,建立二者之间的广义线性混合模型,探究城乡环境变化对鸟类多样性的影响机制,提取显著影响鸟类多样性的环境要素构建预测鸟类多样性分布的评价指标。结果显示,夜间灯光亮度、人口密度对鸟类多样性造成的负面影响较大,而林地、水体面积增加则对鸟类多样性有显著正面影响,同时,景观边缘密度对鸟类多样性也有一定促进作用。因此在生物多样性保护规划中增加林地水体生境时应尽量减少人为干扰的影响,同时在生境修复过程中重点关注斑块形状的复杂度,增加栖息地的异质性和复杂性。
此外,本研究根据模拟潜在鸟类多样性结果判别重要鸟类栖息地并构建武汉都市圈生态网络,通过城市扩张模拟获取鸟类栖息地敏感性评价结果,对生态空间敏感性与重要性评价以及构建刚弹结合的城市生态安全格局具有重要作用。结果显示,武汉都市圈重要鸟类栖息地主要分布于南北两侧山体及长江两岸重要湖泊湿地,形成“一心一链两屏四轴”的生态网络格局,但幕阜山、大别山、长江、东湖、涨渡湖等鸟类栖息地生态敏感性较高,未来城市扩张进程中被侵占的风险较大,需重点保护。
5.1 城市建成环境对鸟类多样性的影响机制
武汉都市圈鸟类多样性空间分布同时受人为因素和自然因素的影响。由结果可知,反映城市化水平的人口密度、夜间灯光亮度与鸟类多样性呈显著负相关,与既往研究相符。这是由于城市化水平较高的区域受人为干扰的影响较大,缺乏足够的栖息地,加上食物匮乏导致大量鸟类难以适应城市环境。此外,有研究指出,夜间灯光对鸟类的进食、睡眠、繁殖等生理行为有广泛影响[31],因此在城市建成区形成了鸟类多样性的洼地。土地利用数据中林地面积[32-33]、水体面积[34-36]与鸟类多样性呈显著正相关,这是由于林地及水体生境中多样的植被组成在提供更多隐蔽空间的同时,还提高了生境异质性[37],不仅能够满足包括树鹨、黑脸噪鹛、白鹭、灰头麦鸡等常见林鸟、水鸟栖息,也吸引了青头潜鸭、白鹤、黑鹳等珍稀濒危鸟类[38]。而不透水表面、农田等其他土地利用类型对鸟类多样性的影响不明显,可能由于本研究所采用的数据精度有限,无法反映城市建成区内部土地利用变化对鸟类多样性的影响。在景观水平上,边缘密度是景观形状复杂度的直观反映,边缘密度与鸟类多样性呈显著正相关,表明景观被边界分割的程度越高,鸟类多样性越大。这可能由于生境形状越复杂,产生的边缘效应越强,而水鸟基于食物的密度而非食物本身的营养或能量来觅食,因此形成了具有丰富食物资源的边缘区域,给大量鸟类提供了良好的栖息环境[39-40]。
5.2 重要鸟类栖息地分布与生态网络构建
武汉都市圈重要鸟类栖息地空间分布结果反映了不同城市McYBsLoSNpwmsMGZIAmXhY4CcSVP91GQNQiuNh6XL1w=环境对鸟类多样性的友好程度,林地和水体构成了鸟类栖息地的主要土地利用类型,与依托重要鸟类栖息地提取的生态廊道一起形成了外部由山体组成生态屏障,内部由水系组成水网,由廊道串连起外部山体和内部水网形成山水相依的生态网络格局。但武汉都市圈西部仙桃市、天门市、潜江市的生态空间较少,与生态网络的联系较为薄弱,东西部的景观联系有待加强。可以通过修复生态断裂点、建设踏脚石和生态廊道差异化保护提升都市圈生态网络的稳定性,更好地发挥生态系统的服务价值[41-42]。此外,要准确评估当前城市生物多样性保护规划的进程,确定每一阶段保护措施的重点,并实施全生命周期管理与保护。随着城市的发展定期监测与反馈,对当下的干预措施进行调整,确保生物多样性保护规划的良性效益[43]。
5.3 城市扩张对生态网络的风险影响
未来城市建设用地仍处于增长趋势,2019—2024年增长速度较快,2024—2034年有所放缓。到2034年,建设用地预计将增加2.49%,主要向北部幕阜山方向及东部延长江发展,长江两岸建设用地扩张较为明显,孝感、鄂州、黄石周边零星碎片状建设用地显著增多。同时,随着建设用地的扩张,水体和林地受到大面积侵占,其中幕阜山北部被侵占面积将达到28.44 km²,大别山南部被侵占面积将达到82.41 km²,林地斑块破碎化程度日益加重。水体生态源地中,长江被侵占面积将达到108.31 km²,东湖被侵占面积将达到3.49 km²,涨渡湖被侵占面积将达到77.19 km²,严重影响重要河湖自然形态及其流动性和连通性[44]。在未来城镇开发建设中既要保护生态安全,又要为城市的社会经济发展保留必要的刚性保护空间,规范管控对生态功能不造成破坏的有限人为活动,严格控制土地管理与相关政策法规,坚持保护有限、自然恢复为主,与人工修复相结合的原则,并结合实际明确生态修复时序安排,定期开展鸟类多样性调查检测,加强关键栖息地保护和遗传资源保存,确保生态安全和社会稳定,协调好生物多样性保护和城市扩张的平衡,践行绿色发展战略[45]。
本研究采用鸟类公众科学数据以探究城市建成环境对鸟类多样性的时空影响机制,但亦存在如鸟类观测点空间分布不均匀等缺陷。在后续研究中,应考虑完善数据来源如通过组织周边城市进行更为详实的鸟类多样性调查,并结合地域性鸟类的分布,为面向鸟类多样性保护的武汉都市圈生态网络建设提供实证支撑,为国土空间规划中面向生物多样性保护的重要生态空间划定与生态修复提供理论参考。
参考文献 References
[1]HAALAND C, BOSCH C. Challenges and strategies for urban green-space planning in cities undergoing densification: a review[J]. Urban Forestry & Urban Greening, 2015, 14(4): 760-771.
[2]HUMPHREY J W, WATTS K, FUENTES-MONTEMAYOR E, et al. What can studies of woodland fragmentation and creation tell us about ecological networks? A literature review and synthesis[J]. Landscape Ecology, 2015, 30(1): 21-50.
[3]GASTON K J. Global patterns in biodiversity[J]. Nature, 2000, 405(6783): 220-227.
[4]蒋志刚,马克平,韩兴国. 保护生物学[M]. 杭州:浙江科学技术出版社,1997.
JIANG Zhigang, MA Keping, HAN Xingguo. Conservation biology[M]. Hangzhou: Zhejiang Science and Technology Publishing House, 1997.
[5]干靓. 城市建成环境对生物多样性的影响要素与优化路径[J]. 国际城市规划,2018,33(4):67-73.
GAN Jing. Built environment factors affecting urban biodiversity and planning optimization approaches[J]. Urban Planning International, 2018, 33(4): 67-73.
[6]ALEXANDRINO E R, BUECHLEY E R, PIRATELLI A J, et al. Bird sensitivity to disturbance as an indicator of forest patch conditions: an issue in environmental assessments[J]. Ecological Indicators, 2016, 66: 369-381.
[7]MORELLI F, BENEDETTI Y, SU T, et al. Taxonomic diversity, functional diversity and evolutionary uniqueness in bird communities of Beijing's urban parks: effects of land use and vegetation structure[J]. Urban Forestry & Urban Greening, 2017, 23: 84-92.
[8]肖华斌,张慧莹,刘莹,等. 自然资源整合视角下泰山区域生态网络构建研究[J]. 上海城市规划,2020(1):42-47.
XIAO Huabin, ZHANG Huiying, LIU Ying, et al. Study on the construction of ecological network of the Taishan Area based on natural resources integration[J]. Shanghai Urban Planning Review, 2020(1): 42-47.
[9]王海洋,王浩琪,陈禧悦,等. 国内外城市生物多样性的评价与提升研究综述[J]. 生态学报,2023(8):2995-3006.
WANG Haiyang, WANG Haoqi, CHEN Xiyue, et al. Review on evaluation and enhancement of urban biodiversity[J]. Acta Ecologica Sinica, 2023(8): 2995-3006.
[10]张征恺,黄甘霖. 中国城市鸟类学研究进展[J]. 生态学报,2018,38(10):3357-3367.
ZHANG Zhengkai, HUANG Ganlin. Urban ornithological research in China: a review[J]. Acta Ecologica Sinica, 2018, 38(10): 3357-3367.
[11]赵伊琳,王成,白梓彤,等. 城市化鸟类群落变化及其与城市植被的关系[J]. 生态学报,2021,41(2):479-489.
ZHAO Yilin, WANG Cheng, BAI Zitong, et al. Changes of bird community under urbanization and its relationship with urban vegetation[J]. Acta Ecologica Sinica, 2021, 41(2): 479-489.
[12]郭诗怡,斋藤馨,夏原由博,等. 城市建成环境对鸟类多样性的影响机制研究评述及展望[J]. 中国园林,2022,38(2):71-76.
GUO Shiyi, SAITO Kaoru, NATUHARA Yoshihiro, et al. The mechanism of urban built environment impact on avian diversity: a systematic review[J]. Chinese Landscape Architecture, 2022, 38(2): 71-76.
[13]VILLASEÑOR N R, ESCOBAR M. Cemeteries and biodiversity conservation in cities: how do landscape and patch-level attributes influence bird diversity in urban park cemeteries?[J]. Urban Ecosystems, 2019, 22(6): 1037-1046.
[14]PLUMMER K E, GILLINGS S, SIRIWARDENA G M. Evaluating the potential for bird‐habitat models to support biodiversity‐friendly urban planning[J]. Journal of Applied Ecology, 2020, 57(10): 1902-1914.
[15]DALE S. Urban bird community composition influenced by size of urban green spaces, presence of native forest, and urbanization[J]. Urban Ecosystems, 2018, 21(1): 1-14.
[16]ALEXANDER J, SMITH D, SMITH Y, et al. Eco-estates: diversity hotspots or isolated developments? Connectivity of eco-estates in the Indian Ocean Coastal Belt, KwaZulu-Natal, South Africa[J]. Ecological Indicators, 2019, 103: 425-433.
[17]ERSOY E, JORGENSEN A, WARREN P H. Identifying multispecies connectivity corridors and the spatial pattern of the landscape[J]. Urban Forestry & Urban Greening, 2019, 40: 308-322.
[18]MIMET A, KERBIRIOU C, SIMON L, et al. Contribution of private gardens to habitat availability, connectivity and conservation of the common pipistrelle in Paris[J]. Landscape and Urban Planning, 2020, 193: 103671.
[19]XUE X, LIN Y, ZHENG Q, et al. Mapping the fine-scale spatial pattern of artificial light pollution at night in urban environments from the perspective of bird habitats[J]. Science of the Total Environment, 2020, 702: 134725.
[20]马远,李锋,杨锐. 城市化对生物多样性的影响与调控对策[J]. 中国园林,2021,37(5):6-13.
MA Yuan, LI Feng, YANG Rui. The impact of urbanization on biodiversity and its regulation countermeasures[J]. Chinese Landscape Archi-tecture, 2021, 37(5): 6-13.
[21]SHIRK J, BALLARD H L, JORDAN R C, et al. Public participation in scientific research: a framework for intentional design[C]//96th ESA Annual Convention. 2011.
[22]SILVERTOWN J. A new dawn for citizen science[J]. Trends in Ecology & Evolution, 2009, 24(9): 467-471.
[23]刘萌萌,张曼玉,韩茜,等. 公众观鸟和传统样线法调查应用于鸟类多样性监测的比较:以南京老山为例[J]. 生态与农村环境学报,2023,39(9):1196-1204.
LIU Mengmeng, ZHANG Manyu, HAN Qian, et al. Comparing the effectiveness of birdwatching to line transect for biodiversity surveys: a case study of Laoshan Nanjing[J]. Journal of Ecology and Rural Environment, 2023, 39(9): 1196-1204.
[24]俞孔坚. 生物保护的景观生态安全格局[J]. 生态学报,1999(1):10-17.
YU Kongjian. Landscape ecological security patterns in biological conservation[J]. Acta Ecologica Sinica, 1999(1): 10-17.
[25]刘世梁,侯笑云,尹艺洁,等. 景观生态网络研究进展[J]. 生态学报,2017,37(12):3947-3956.
LIU Shiliang, HOU Xiaoyun, YIN Yijie, et al. Research progress on landscape ecological networks[J]. Acta Ecologica Sinica, 2017, 37(12): 3947-3956.
[26]阎凯,王宝强,沈清基. 上海市生态网络体系评价方法研究[J]. 上海城市规划,2017(2):82-89.
YAN Kai, WANG Baoqiang, SHEN Qingji. Research on evaluation and construction of ecological network system in Shanghai[J]. Shanghai Urban Planning Review, 2017(2): 82-89.
[27]李权荃,金晓斌,张晓琳,等. 基于景观生态学原理的生态网络构建方法比较与评价[J]. 生态学报,2023,43(4):1461-1473.
LI Quanquan, JIN Xiaobin, ZHANG Xiaolin, et al. Comparison and evaluation of the ecological network construction method based on principles of landscape ecology[J]. Acta Ecologica Sinica, 2023, 43(4): 1461-1473.
[28]杨凯,曹银贵,冯喆,等. 基于最小累积阻力模型的生态安全格局构建研究进展[J]. 生态与农村环境学报,2021,37(5):555-565.
YANG Kai, CAO Yingui, FENG Zhe, et al. Research progress of security pattern construction based on minimum cumulative resistance model[J]. Journal of Ecology and Rural Environment, 2021, 37(5): 555-565.
[29]田雅楠,张梦晗,许荡飞,等. 基于“源—汇”理论的生态型市域景观生态安全格局构建[J]. 生态学报,2019,39(7):2311-2321.
TIAN Ya'nan, ZHANG Menghan, XU Dangfei, et al. Landscape ecological security patterns in an ecological city based on source-sink theory[J]. Acta Ecologica Sinica, 2019, 39(7): 2311-2321.
[30]吕乐婷,张杰,彭秋志,等. 东江流域景观格局演变分析及变化预测[J]. 生态学报,2019,39(18):6850-6859.
LYU Leting, ZHANG Jie, PENG Qiuzhi, et al. Landscape pattern analysis and prediction in the Dongjiang River Basin[J]. Acta Ecologica Sinica, 2019, 39(18): 6850-6859.
[31]汪婷,周立志. 合肥市小微湿地鸟类多样性的时空格局及其影响因素[J]. 生物多样性,2022,30(7):146-158.
WANG Ting, ZHOU Lizhi. The spatial-temporal of bird diversity and its determinants in the small wetlands in Hefei City[J]. Biodiversity Science, 2022, 30(7): 146-158.
[32]郑进凤,唐蓉,贺霜,等. 贵州花溪大学城破碎化林地鸟类多样性与嵌套分布格局[J]. 生物多样性,2021,29(5):661-667.
ZHENG Jinfeng, TANG Rong, HE Shuang, et al. Bird diversity and nestedness on fragmented woodlots in Huaxi University Town, Guizhou Province[J]. Biodiversity Science, 2021, 29(5): 661-667.
[33]杨刚,许洁,王勇,等. 城市公园植被特征对陆生鸟类集团的影响[J]. 生态学报,2015,35(14):4824-4835.
YANG Gang, XU Jie, WANG Yong, et al. The influence of vegetation structure on bird guilds in an urban park[J]. Acta Ecologica Sinica, 2015, 35(14): 4824-4835.
[34]张颖,黄婷婷,胡骞,等. 基于鸟类多样性提升的社区公园生境营造策略探析[J]. 中国园林,2022,38(3):106-111.
ZHANG Ying, HUANG Tingting, HU Qian, et al. Analysis on the habitat strategy of community park based on the improvement of bird diversity[J]. Chinese Landscape Architecture, 2022, 38(3): 106-111.
[35]宫蕾,张黎黎,周立志,等. 长江中下游安庆沿江湖泊湿地夏季鸟类多样性调查[J]. 湖泊科学,2013,25(6):872-882.
GONG Lei, ZHANG Lili, ZHOU Lizhi, et al. Bird diversity in summer in Anqing floodplain wetlands, middle-lower reaches of the Yangtze River[J]. Journal of Lake Sciences, 2013, 25(6): 872-882.
[36]刘大钊,周立志. 安徽安庆菜子湖国家湿地公园景观格局变化对鸟类多样性的影响[J]. 生态学杂志,2021,40(7):2201-2212.
LIU Dazhao, ZHOU Lizhi. Effects of landscape pattern change on bird diversity in Anhui Anqing Caizi Lake National Wetland Park[J]. Chinese Journal of Ecology, 2021, 40(7): 2201-2212.
[37]张鹏骞,张志明,白加德,等. 北京南海子麋鹿苑鸟类多样性研究[J]. 生态学报,2020,40(5):1740-1749.
ZHANG Pengqian, ZHANG Zhiming, BAI Jiade, et al. Bird diversity in Nan-Haizi Milu Park of Beijing, China[J]. Acta Ecologica Sinica, 2020, 40(5): 1740-1749.
[38]ZHOU D, CHU L M. How would size, age, human disturbance, and vegetation structure affect bird communities of urban parks in different seasons?[J]. Journal of Ornithology, 2012, 153: 1101-1112.
[39]李祎可,王强,李星醇,等. 边缘效应对湿地中鸟类的影响机制研究进展[J]. 湿地科学,2022,20(5):613-621.
LI Yike, WANG Qiang, LI Xingchun, et al. Progress on the impact mechanism of edge effect on birds in wetlands[J]. Wetland Science, 2022, 20(5): 613-621.
[40]刘骏杰,陈璟如,来燕妮,等. 基于景观格局和连接度评价的生态网络方法优化与应用[J]. 应用生态学报,2019,30(9):3108-3118.
LIU Junjie, CHEN Jingru, LAI Yanni, et al. Improvement and application for ecological networks using landscape pattern and connectivity methods[J]. Chinese Journal of Applied Ecology, 2019, 30(9): 3108-3118.
[41]肖华斌,施俊婕,盛硕,等. 生态系统服务优化导向下城市绿色基础设施构建研究——以济南市西部新城为例[J]. 上海城市规划,2019(1):45-50.
XIAO Huabin, SHI Junjie, SHENG Shuo, et al. Research on the construction of green infrastructure under the guidance of ecosystem service optimization: a case study of Ji'nan Western New District[J]. Shanghai Urban Planning Review, 2019(1): 45-50.
[42]李红波,黄悦,高艳丽. 武汉城市圈生态网络时空演变及管控分析[J]. 生态学报,2021,41(22):9008-9019.
LI Hongbo, HUANG Yue, GAO Yanli. Analysis on the spatio-temporal evolution and regulation of the ecological network in Wuhan Metropolitan Area[J]. Acta Ecologica Sinica, 2021, 41(22): 9008-9019.
[43]王思元,李慧. 基于景观生态学原理的城市边缘区绿色空间系统构建探讨[J]. 城市发展研究,2015,22(10):20-24.
WANG Siyuan, LI Hui. Analysis of shaping the urban fringe greenspace system based on landscape ecology principles[J]. Urban Development Studies, 2015, 22(10): 20-24.
[44]吴榛,张凯云,王浩. 城市扩张情景模拟下绿地生态网络构建与优化研究——以南京市部分区域为例[J]. 中国园林,2022,38(4):56-61.
WU Zhen, ZHANG Kaiyun, WANG Hao. Construction and optimization of green space ecological network under urban expansion scenario simulation: a case study in some areas of Nanjing[J]. Chinese Landscape Architecture, 2022, 38(4): 56-61.
[45]彭宏杰,花磊,张雪松,等. 基于生态敏感性评价的城市用地扩张模拟研究[J]. 长江流域资源与环境,2022,31(1):83-92.
PENG Hongjie, HUA Lei, ZHANG Xuesong, et al. Simulation research on urban land expansion based on ecological sensitivity evaluation[J]. Resources and Environment in the Yangtze Basin, 2022, 31(1): 83-92.
基金项目:国家自然科学基金青年基金“平原水网城市建成环境对鸟类多样性的影响机制与规划响应”(编号52208083);第74批中国博士后科学基金面上资助项目“长江流域生态空间对鸟类多样性的影响机制与空间优化研究”(编号2023M742694);湖北省文旅厅重点项目“基于‘生态—文化’耦合评价的长江国家文化公园(湖北段)四类功能区识别与规划建议”(编号HCYK2022Z08)资助。
