
基于模拟数据分析长江调水对棘洪滩水库的影响
2024-09-03 00:00:00周守平陈立国张烨徐欣吕慎海刘本陈建锋
山东农业大学学报(自然科学版) 2024年3期
关键词:南水北调



摘 要: 目前南水北调东线调水工程水源复杂多变,显著影响了受水水体的水质与浮游藻类。为量化与评估这一影响,本研究在调水周期内采集不同季节的供水(长江水)、受水(棘洪滩水库水)水样,同时开展室内藻类生长潜力试验,比较分析了不同比例的供、受水体混合处理后的理化因子、浮游藻类细胞密度与群落结构的变化情况。结果显示,春、秋季长江水的引入一定程度上削弱了棘洪滩水库的营养负荷;pH在季节间均表现为下降的趋势;秋季长江水的引入导致棘洪滩水体浮游藻类密度峰值显著增加(Plt;0.05),而在春季,仅当长江调水比例高于40%时,藻类密度峰值才随调水量的增加而显著增大(Plt;0.05)。群落结构方面,春、秋季长江与棘洪滩水体浮的游藻类均以蓝藻为主要优势,不同的是,春季以色球藻属(Chroococcus)的种类占主要优势,秋季则以假鱼腥藻属(Pseudanabaena)、尖头藻属(Raphidiopsis)与细鞘丝藻属(Leptolyngbya)的种类为优势。随长江水引入比例的增加,春季调水进一步增加了色球藻属的优势度;秋季调水则削弱了尖头藻属优势度同时增加了假鱼腥藻属的优势度。综合表明,不同季节长江调水对棘洪滩水库的浮游藻类生长影响存在差异,浮游藻类群落结构显著受到供水水体与季节的综合影响。
关键词: 南水北调;棘洪滩水库;藻类生长潜力;浮游藻类群落结构
中图法分类号: X835 文献标识码: A 文章编号: 1000-2324(2024)03-0466-09
南水北调是为缓解我国北方水资源严重短缺而实施的重大战略性工程。鉴于不同地区水资源配置与实际情况,南水北调东线和胶东调水工程联合为胶东半岛供水,其主要水源包括长江水、黄河水与本地湖库水。已有研究表明,长江调水显著改善了沿线受水区的水生态状况[1-3]。引江济巢调水工程对巢湖的水质和水动力水质条件改善明显,且流量越大,效果越显著[4]。引江济太工程利用长江水有效缓解了太湖浮游植物聚集的情况,但对太湖生态系统稳定性造成了一定的影响[5]。南水北调中线工程的实施导致丹江口库区浮游植物群落结构发生变化[6]。实际上,任何调水工程都可能对受纳系统造成复杂的物理、化学、水文和生物影响[7-8]。例如中国的南水北调工程和长江-淮河调水工程虽然在一定程度上缓解了受水区的用水危机,但也带来了跨流域的环境问题[9-12]。巴西的引水工程造成了与生物入侵和同质化相关的生物多样性影响[13-14]。许多研究提出了评估调水工程影响的方法[15-19],但多集中在数值模拟上,而实际调水过程中环境因子的复杂动态变化使得模型预测存在较大误差,这可能导致对调水的影响评估不准确。浮游藻类作为水生生态系统的重要初级生产者,对水体环境变化更为敏感,能在短时间内做出快速响应[20]。Song 等使用藻类生长潜力试验评估了由金沙江和巴清河调水对程海湖藻类群落结构的影响,金沙江调水将导致程海湖浮游藻类群落结构的优势种由鱼腥藻转变为鱼腥藻与小环藻共存,而巴清河调水将进一步提升鱼腥藻的优势地位[21]。也有研究认为调水将导致调蓄湖泊蓝藻的相对丰度的下降,对优势种的影响更为强烈[22],由调水引起浮游藻类群落结构的变化尚有待研究。
东平湖作为东线长江水输水线路上的最后一个调蓄湖泊,而棘洪滩水库作为青岛市重要的饮用水源地,两者重要的战略地位是显而易见的。长江调水洪滩水库,其水质的变化已成为公众关注的焦点,也关乎到调水工程的成败。利用藻类生长情况对调水情况进行预测具有巨大潜力,这一方法已在调水评估中得到很好的应用[23-24]。鉴于上述原因,本研究利用藻类生长潜力试验探讨了在不同季节的调水周期内,长江调水对棘洪滩水库理化因子、浮游藻类细胞密度与群落结构的影响。旨在为保障水质安全提供数据支持,为跨流域调水中存在的共性问题提供解决思路,以及为调水部门提供基于生态系统水平的管理建议。
1 材料和方法
1.1 样品采集与理化因子测定
样品的采集:于2022 年10 月与2023 年4 月东平湖调水周期内,采集东平湖(长江水)中部和棘洪滩水库出水口表层水5 L,经过100 μm筛绢滤去大型浮游动物,水样被收集在酸洗的聚乙烯塑料瓶中,每个样点重复采样两次,低温微光环境保存,24 h 内开展试验。采用手持式pH仪(中国雷磁,PHB-4)测定水体pH。部分水样添加硫酸至pH 为1~2,常温保存,用于测定总氮(TN),总磷(TP),总可溶性氮(TDN),总可溶性磷(TDP),分析参考标准方法[25]。
1.2 浮游藻类生长潜力试验
试验方案:根据实际调水情况与研究目的,将经过处理后的水样按照不同比例混合,模拟实际调水情况。具体采样季节和试验方案见表1。
培养条件:温度为24±1 ℃,光源为白色日光灯,光照强度控制在5 000±10% lx,光暗比设置为14∶10。培养容器为容积250 mL锥形瓶,试验溶液体积设置为150 mL,于恒温光照培养箱中进行培养,每隔3 h 进行人工摇动,并随机交换位置。
藻类鉴定与计数:每隔3 d 取样,每个三角烧瓶取15 mL水样,以3 000 rpm 的转速离心,浓缩体积至2 mL,上清液返还至原三角烧瓶中。将底部藻液混匀,取0.1 ml 藻液在光学显微镜下利用浮游藻类计数框进行计数和鉴定,剩余藻液返还至原三角烧瓶中。浮游藻类鉴定参考《中国淡水藻类———系统、分类及生态》[26],计数参考《淡水浮游生物研究方法》[27]。
1.3 数据处理与分析
绘图与数据分析软件使用Origin 和SPSS25.0,采用One way-ANOVA 方法对数据进行显著性分析,采用最小显著差异法(LSD)检验各组间差异的显著性。
2 结果
2.1 不同季节及不同混合比例下试验水体的理化因子变化情况
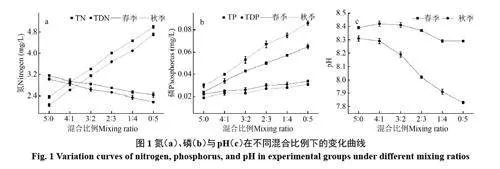
春季棘洪滩水体总氮含量为3.16 mg/L,总可溶性氮含量为3.03 mg/L,长江水源略低,总氮与总可溶性氮含量分别为2.44 和2.17 mg/L。棘洪滩水库总磷与总可溶性磷含量分别为0.024 和0.022 mg/L,长江水源则略高,含量达到0.065 和0.034 mg/L(图1)。秋季棘洪滩水体氮磷含量均低于长江水源但整体营养水平均高于春季,棘洪滩水体总氮与总可溶性氮含量分别为2.36 和2.06 mg/L,长江水源分别为4.98 和4.7 mg/L,棘洪滩水库总磷与总可溶性磷含量分别为0.03 和0.019 mg/L,长江水源分别为0.086和0.03 mg/L。
随着长江与棘洪滩水体的混合比例的增大,春季和秋季理化因子变化结果相似。受到长江水源与棘洪滩水库水体营养盐差异的影响,随着混合比例的增加,总氮与总可溶性氮含量在春季随之减小,秋季则随之增大;总磷与总可溶性磷含量在春与秋季随之增加。春季与秋季棘洪滩水库pH 均低于长江水体,春季随着混合比例的增加,pH略有上升后下降,秋季则随着混合比例的增加,混合水体pH不断下降(图1)。
2.2 不同季节及不同混合比例下试验水体的藻类生长情况
不同季节长江水的引入对棘洪滩浮游藻类生长的影响存在差异(图2),在春季,随着培养时间的延长,实验组的藻类密度呈上升趋势,在16~20 d 达到密度峰值。藻类密度峰值显示,当混合比例在5∶0 与3∶2 之间时,改变混合比例对藻类密度峰值无显著性(Pgt;0.05)差异,处理组的藻类密度峰值与棘洪滩对照组的密度相近,当混合比例超过3∶2 时,随着混合比例的增加,藻类密度峰值随之显著(Plt;0.05)增大,最终与长江水对照组藻类相近。在秋季,实验组藻类密度随着培养时间的延长而增加,在12~20 d 达到密度峰值,与春季不同,当混合比例在5∶0 与3∶2 之间时,藻类密度峰值随混合比例的增加而显著(Plt;0.05)增大,当混合比例超过过3∶2 时,藻类密度峰值达到最大值,且随调水比例增加而保持相对稳定。
春季棘洪滩水库浮游藻类组成较均匀,以蓝藻、绿藻与硅藻为主,蓝藻中色球藻属(Chroococcus)物种相对丰度最高,达到25.8%;绿藻中卵囊藻属(Oocystis)物种相对丰度最高,达到21.2%;硅藻中针杆藻属(Synedra)物种相对丰度最高,达到15.9%。长江水体浮游藻类群落结构与棘洪滩水库存在差异,蓝藻中假鱼腥藻属(Pseudanabaena)物种相对丰度最高,为18.3%;绿藻中卵囊藻属(Oocystis)物种相对丰度最高,为12%;硅藻中直链藻属(Melosira)物种的相对丰度最高,为16%。到达培养末期(20 d)时,棘洪滩水库与长江水体对照组的浮游藻类群落结构相对于培养初期(0 d)存在差异,该时期蓝藻占据了主要的优势地位,蓝藻相对丰度超过80%,构成以色球藻属为主,相对丰度超过60%。在混合处理组中,随着长江水混合比例的增加,色球藻属物种相对丰度先升高后下降,最高由棘洪滩对照组的65%升至89.7%,随后降至长江水体对照组的66.7%,而假鱼腥藻属物种的相对丰度先减小而后升高,由棘洪滩水库对照组的18.6%最低降至2.7%,随后升至27.5%,色球藻属物种相对丰度最大时,假鱼腥藻属物种相对丰度最小,此时混合比例为2∶3,针杆藻属的优势度随着长江水的混入比例升高而不断下降,在混合比例大于3∶2 时,相对丰度仅为1% 左右(图3)。
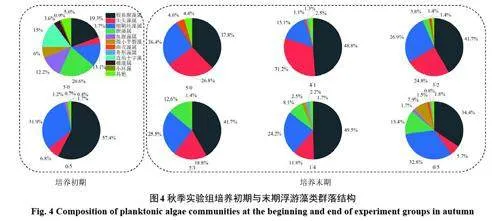
秋季长江水体与棘洪滩水库水体藻类群落结构均以蓝藻为主,初期棘洪滩水库蓝藻组成主要以假鱼腥藻属(Pseudanabaena)、细鞘丝藻属(Leptolyngbya)、颤藻属(Oscillatoria)与鱼腥藻属(Anabaena)为主,而长江水体中以假鱼腥藻属(Pseudanabaena)和细鞘丝藻属(Leptolyngbya)相对丰度最高。两水体混合之后,假鱼腥藻属的相对丰度在混合组中高于40%,且大于其在对照组中的丰度。随着混合比例的升高,尖头藻属(Raphidiopsis)的相对丰度逐渐下降,而细鞘丝藻属的相对丰度基本保持不变,维持在25%附近(图4)。
3 讨论
多数情况下,氮磷含量被认为是影响藻类生长的关键因子,而磷是湖泊藻类生长的限制性因子,但与湖泊富营养化程度相关[28,29]。东平湖(长江水)与棘洪滩水库春、秋季总氮浓度均较高,处于2~4 mg/L 之间,而东平湖(长江水)春、秋季磷含量高于棘洪滩水库,东平湖(长江水)春、秋季磷含量分别为0.065 和0.086 mg/L,棘洪滩水库则处于0.03 mg/L左右。氮磷比被认为与湖泊生态系统中影响藻类氮磷限制相关[30],棘洪滩水库氮磷比在79~132 之间,东平湖(长江水)氮磷比在38~59 之间,均超过之前报导的磷限制的下限值[31,32],东平湖(长江水)与棘洪滩水库影响藻类生长的关键因子可能为磷,这一点已在之前的研究中得到证实[33]。棘洪滩水库磷含量低于东平湖(长江水),东平湖水体额外的磷输入可能是促进棘洪滩水库藻类生长的原因。但不同季节表现存在差异,春季长江混合比例低于40%时,不会引起浮游藻类细胞密度峰值显著(Plt;0.05)增加;而秋季长江水的引入会引起浮游藻类密度峰值的显著(Plt;0.05)增加,并在混合比例大于等于40%时,浮游藻类密度峰值达到最大值。这可能与浮游藻类的生长特性相关,藻类生物量与营养盐浓度间的关系类似Logistic 增长曲线,只有当外界可利用的营养盐浓度超过一定限度才能引起藻类的快速生长[34]。有研究发现当水体可溶性磷浓度超过0.04 mg/L[33]时,藻类生物量才随着磷浓度的增加而显著升高,Kristiina 等认为在北方湖泊中,将总磷浓度控制在0.05 mg/L以下时,有助于控制蓝藻水华[35]。作者认为春季长江调水小于等于40%的调水量,对应总磷含量不超过0.043 mg/L时,不会引起棘洪滩水库浮游藻类密度的显著升高。秋季则与春季不同,表现在春季棘洪滩水库浮游藻类细胞密度受东平湖调水影响有限,而秋季则受东平湖调水影响明显,可能与春秋季的营养盐含量、pH的差距和浮游藻类群落结构相关。长江水的磷含量秋季高于春季,相同混合比例下,秋季长江水能为棘洪滩水库提供更多的磷,而棘洪滩水体的藻类生长限制因子为磷[33],因此秋季长江水体的引入导致了棘洪滩水库磷浓度的增加,进而引起了棘洪滩水库水体藻类密度峰值的显著升高(Plt;0.05)。虽然在混合比例达到20%时,棘洪滩总磷浓度达到0.04 mg/L,仍旧低于之前推测的棘洪滩水库引起藻类生长的磷的最小阈值浓度,但依然引起藻类密度峰值的显著增加(Plt;0.05)。这可能与春秋季棘洪滩水库水体浮游藻类群落结构相关,秋季假鱼腥藻占据主要优势地位,而假鱼腥藻受氮和磷浓度影响较大[36],秋季长江水的混入提高了棘洪滩水库的氮磷含量(图1),导致了棘洪滩水库以假鱼腥藻为主要优势的浮游藻类细胞密度的增加。此外,水体中pH 含量主要与碳酸盐缓冲体系相关,较低的pH 表明东平湖水体中承载了更多的CO2,为藻类生长提供了更多的碳源,有利于浮游藻类的生长,也是秋季长江水的引入导致棘洪滩水库浮游藻类密度峰值显著增加的原因。
考虑到水库的缓冲能力,棘洪滩水库持续调水可能不会引起营养盐浓度的显著上升[37],但由于调水短期引入的藻类与营养盐可能对藻类的生长产生影响,藻类实际生长情况可能与调水本身相关。在本研究中,东平湖(长江水)与棘洪滩水库的混合水最高可使棘洪滩水库藻类密度升高至混合前的两倍,但不会引起棘洪滩水库浮游藻类的异常增殖,因此经东平湖调长江水引起棘洪滩水库藻类水华的概率较低。春季棘洪滩水库在混合比例低于60%情况下,仍能维持在原有浮游藻类密度附近,秋季东平湖水体的混入则会显著引起棘洪滩水库浮游藻类显著升高,因此,基于藻类生长潜力测试,考虑藻类丰度指标,由东平湖调水至棘洪滩水库春季更合适。
在藻类群落结构方面,不同种类的浮游藻类生存策略存在差异,一般包含C 策略(Competitive)、S 策略(Stress-tolerant)、R 策略(Ruderal)三种不同类型的生存策略[38]。当存在低强度的干扰和环境压力时,C策略物种倾向于占主导地位。在低干扰强度和高压力条件下,作为抗胁迫物种的S)策略种往往在竞争中胜出,R策略种在低压力和高干扰环境中有望成为优势[39]。色球藻属(Chroococcus)为S 型[40],面对不同比例混合下的非稳态环境可能占据一定的优势地位。在春季实验中,当长江水比例在40%和60%时,由于水体混合程度与培养压力的增加,色球藻属相对丰度增加,最高达到90%;而混合比例在0、20%或80%、100%,其混合程度低,色球藻属相对丰度与对照组相近。根据浮游植物的形态、生理和生存策略,Reynolds 等[41]在生态系统中引入功能群(functional group, FG)概念,进一步细化和分类。尖头藻属(Raphidiopsis)属于SN功能群[42],其生境特点是混合和温暖,一定程度上解释了秋季相对春季尖头藻属相对丰度增加的原因。棘洪滩水库受到外来水源混合影响,可能导致尖头藻属持续占优。但在秋季,当长江水比例超过40%,其相对丰度随着混合比例的增大而减小,最终与长江水对照组类似。假鱼腥藻属(Pseudanabaena)属于S1 功能群[42],生境特点为浑浊的混合环境,虽然培养环境水体透明度较高,但假鱼腥藻被认为与氮磷浓度正相关[43],秋季东平水体的混入显著增加了氮磷含量,引起了假鱼腥藻属的相对丰度的增加。春季东平湖(长江水)的输入导致了棘洪滩水库色球藻属相对丰度的增加,并在混合比例为40%与60%下达到绝对优势,相对丰度超过80%,表明色球藻属是棘洪滩水库春季调水过程中潜在的优势藻种,适宜环境条件下可能成为棘洪滩水库春季的绝对优势种。秋季调水导致假鱼腥藻属相对丰度增加,表明持续调水可能增加假鱼腥藻的优势且持续占优势,这主要受到水源地的浮游藻类与季节双重因素驱动。
值得注意的是,假鱼腥藻属已经成为山东境内多个调蓄湖泊潜在的水华种[44]。对山东省第一大水库峡山水库的浮游植物优势种生态位及种间联结性动态分析显示,假鱼腥藻在多个季节具有最大的生态位宽度及较强的适应多样化生境的能力,可在冬季迅速发展的优势蓝藻[45]。此外,假鱼腥藻属某些种类可能产异味物质,例如2- 甲基异莰醇[46],且已在青岛居民用水中检出[47]。因此调水管理部门需密切关注在调水过程中假鱼腥藻对受水水库的影响。
藻类生长潜力试验是在较为适合的环境,记录藻类群落结构的演替规律,找出单次调水情况下具有较高生长潜力的藻种,借以预测实际调水过程中藻类群落结构的变化,以此为调水管理部门的决策提供科学的理论基础数据。本研究通过藻类生长潜力试验模拟了长江经东平湖调水,棘洪滩水库浮游藻类的变化情况,初步给出了调水量阈值和分析了优势蓝藻可能带来的风险。客观的讲,这一模拟与现实调水情况可能存在一定的出入,其原因是现实调水浮游植物变化还可能受多个物理、化学和生物等复杂因素影响。但本研究的结果初步给出了调水浮游藻类变化的合理区间和趋势,这对调水前评估调水对受水水库水质影响具有积极重要的意义。
鉴于胶东调水工程涉及多水源调水,受水水库众多,作者建议在水源地本底情况不清的情况下,调水前很有必要进行藻类生长潜力实验,预测浮游植物群落变化,防止调水引起藻类异常增殖形成水华;此外需根据室内模拟试验的结果,选择合适的季节和调水量进行生态调水。目前作者仅关注单一水源调水棘洪滩水库的浮游植物群落变化情况,作为胶东调水工程重要的调蓄水库,多水源的混合调度汇集于棘洪滩水库对其的影响程度还需进一步的监测评估。未来还需结合野外数据,动态分析浮游藻类群落结构演替规律,合理调度水资源,保障居民用水安全。
4 结论
长江经东平湖调水至棘洪滩水库,增加了受水水库营养盐浓度,持续的调水输入改变了棘洪滩水库藻类群落结构,增加了蓝藻的相对丰度。春季调水比例超过棘洪滩库容40%和秋季调水时,将显著引起棘洪滩水库浮游藻类密度峰值的升高。基于维持受水水库浮游藻类密度保持相对稳定方面,本研究建议长江经东平湖调水棘洪滩水库应选择春季调水较好,且调水比例应不超过40%。春秋季两水体混合过程中,实验组浮游藻类密度峰值均未达到藻类水华标准,因此认为春、秋季由长江经由东平湖向棘洪滩调水的藻类水华风险较小,但秋季调水过程中假鱼腥藻属的相对丰度有所增加,需要关注该藻对水库饮水安全的影响。
参考文献
[1] Qu X, Chen Y, Liu H, et al. A holistic assessment of
water quality condition and spatiotemporal patterns
in impounded lakes along the eastern route of
China's South-to-North water diversion project[J].
Water Research, 2020, 185:116275.
[2] Guo C, Chen Y, Xia W, et al. Eutrophication and
heavy metal pollution patterns in the water suppling
lakes of China's south-to-north water diversion
project[J]. Science of the Total Environment, 2020,
711:134543.
[3] Guo C, Chen Y, Gozlan R E, et al. Patterns of fish
communities and water quality in impounded lakes
of China's south-to-north water diversion project[J].
Science of the Total Environment, 2020, 713:136515.
[4] 菅浩然,韩 涛. 引江济巢工程对改善巢湖水质的数
值模拟分析[J]. 水电能源科学,2020,38(9):53-55
[5] 殷鹏,华萍,胡晓雨.2007—2020 年引江济太环境
效益分析及工程优化调度研究[J]. 水利建设与管
理,2022,42(09): 55-59+69.
[6] 董磊,吴敏,林莉,等. 南水北调中线工程对丹江
口水库浮游植物群落结构影响[J]. 环境科学与技
术,2021,44(S1):1-7.
[7] Qiu Y, Zhang Y, Lan P, et al. Influence of
Atmospheric Phosphorus and Nitrogen Sedimentation
on Water Quality in the Middle Route Project of the
South-to-North Water Diversion in Henan Province
[J]. International Journal of Environmental Research
and Public Health. 2022, 19(21):14346.
[8] Zhang S, Pang Y, Xu H, et al. Shift of phytoplankton
assemblages in a temperate lake located on the
eastern route of the south-to-north water diversion
project[J]. Environmental research, 2023, 227:
115805.
[9] Huang H, Wang Y, Wang S, et al. Saltwater Intrusion
in the Changjiang River Estuary in Response to the
East Route of the South-to-North Water Transfer
Project in the New Period after 2003[J]. Sustainability,
2024, 16(2):14346.
[10] Sun R, Wei J, Zhang S, et al. The dynamic changes
in phytoplankton and environmental factors within
Dongping Lake (China) before and after the Southto-
North Water Diversion Project[J]. Environmental
Research, 2024, 246:118138.
[11] Yang H, Li Y, Liu H, et al. The variation of DOM
during long distance water transport by the China
South to North Water Diversion Scheme and impact
on drinking water treatment[J]. Frontiers of
Environmental Science Engineering, 2024, 18(5):59.
[12] Cao ZG, Li S, Zhao YE, et al. Spatio-Temporal
Pattern of Schistosomiasis in Anhui Province, East
China: Potential Effect of the Yangtze River-Huaihe
River Water Transfer Project[J]. Parasitology
International, 2018, 67:538-546 .
[13] Daga VS, Azevedo-Santos VM, Pelicice FM, et al.
Water Diversion in Brazil Threatens Biodiversity[J].
Ambio, 2020, 49:165-172.
[14] Yuan R, Wang M, Wang S, et al. Water Transfer
Imposes Hydrochemical Impacts on Groundwater by
Altering the Interaction of Groundwater and Surface
Water[J]. Journal of Hydrology, 2020, 583:124617.
[15] Guo S, Huang J, Zhang X, et al. Ecological
sensitivity study of the South-North Water Transfer
central line water source area based on land use
change[J]. Environmental monitoring and assessment,
2023, 195(3):384-384.
[16] Zhuang W, Ying SC, Frie AL, et al. Distribution,
pollution status, and source apportionment of trace
metals in lake sediments under the influence of the
South-to-North Water Transfer Project, China[J].
Science of The Total Environment, 2019, 671:
108-118.
[17] Zhao G, Gao X, Zhang C, et al. The effects of
turbulence on phytoplankton and implications for
energy transfer with an integrated water qualityecosystem
model in a shallow lake[J]. Journal of
Environmental Management, 2020, 256:109954.
[18] Lee HW, Yeom BM, Choi JH. Modelling water
quality improvements in a South Korean inter-basin
water transfer system [J]. Water, 2020, 12:3173.
[19] 李国辉,李莉杰,李根保. 基于EFDC 的滇池藻类模
型参数敏感性时空特征[J]. 应用与环境生物学报,
2021,27:1047-1054.
[20] Anjusha A, Jyothibabu R, Savitha KMM, et al.
Seasonal variation of phytoplankton growth and
microzooplankton grazing in a tropical coastal water
(off Kochi), southwest coast of India[J]. Continental
Shelf Research, 2018, 171:12-20.
[21] Song Y, Qi J, Deng L, et al. Selection of water source
for water transfer based on algal growth potential to
prevent algal blooms[J]. Journal of Environmental
Sciences, 2021, 10(3):246-254.
[22] Li Q, Wang G, Tan Z, et al. Succession of
phytoplankton in a shallow lake under the alternating
influence of runoff and reverse water transfer[J].
Hydrology Research, 2020, 51(5):1077-1090.
[23] Fott J, Nedbalova L, Brabec M, et al. Light as a
controlling factor of winter phytoplankton in a
monomictic reservoir[J]. Limnologica, 2022, 95:
125995.
[24] Li Q, Wang G, Tan Z, et al. Succession of
phytoplankton in a shallow lake under the alternating
influence of runoff and reverse water transfer[J].
Hydrology Research, 2020, 51:1077-1090.
[25] 国家环境保护总局«水和废水监测分析方法»编委
会. 水和废水监测分析方法 (第四版)[M]. 北京:中
国环境科学出版社,2002.
[26] 胡鸿钧,魏印心. 中国淡水藻类——系统、分类及
生态[M]. 北京:科学出版社:2006,5-1020.
[27] 章宗涉,黄祥飞,淡水浮游生物研究方法[M]. 北京:
科学出版社:1991,333-344.
[28] Mamun M, Lee S, An K. Roles of Nutrient Regime
and N: P Ratios on Algal Growth in 182 Korean
Agricultural Reservoirs. Polish Journal of Environmental
Studies. 2018, 27(3):1175-1185.
[29] Liang Z, Soranno PA, Wagner T. The role of
phosphorus and nitrogen on chlorophyll a: Evidence
from hundreds of lakes[J]. Water Research, 2020,
185:116236.
[30] Qin B, Zhou J, Elser JJ, et al. Water depth underpins
the relative roles and fates of nitrogen and
phosphorus in lakes[J]. Environmental Science amp;
Technology, 2020, 54:3191-3198.
[31] Smith VH. The nitrogen and phosphorus dependence
of algal biomass in lakes: An empirical and
theoretical analysis[J]. Limnology and Oceanography,
1982, 27:1101-1112.
[32] Kolzau S, Wiedner C, Rücker J, et al. Seasonal
patterns of nitrogen and phosphorus limitation in
four German lakes and the predictability of limitation
status from ambient nutrient concentrations[J]. PLoS
ONE, 2014, 9:e96065.
[33] Xue L, Hu J, Wang Z. et al. Assessing risks of algal
blooms in water transfer based on algal growth
potential[J]. Environmental Monitoring and Assessment,
2023, 195:871.
[34] Bolier G, Donze M. On the accuracy and
interpretation of growth curves of planktonic algae
[J]. Hydrobiologia, 1989, 188:175-179.
[35] Vuorio K, Järvinen M, Kotamäki N. Phosphorus
thresholds for bloom-forming cyanobacterial taxa in
boreal lakes[J]. Hydrobiologia, 2020, 847:4389-4400.
[36] 韦洁琳,崔玉静,李亦真,等. 南水北调东线山东境内
调蓄湖库蓝藻种群分布特征及环境驱动因子[J]. 环
境科学,2023,44(11):6137-6148.
[37] 丁 宁,杨莹莹,万年新,等. 调蓄水库细菌群落季节
变化特征与影响因素[J]. 环境科学,2023,44:1484-
1496.
[38] Marazzi L, Gaiser EE, Jones VJ, et al. Algal richness
and life ‐ history strategies are influenced by
hydrology and phosphorus in two major subtropical
wetlands[J]. Freshwater Biology, 2017, 62:274-290.
[39] Xiao Y, Li Z, Guo J, et al. Succession of
phytoplankton assemblages in response to largescale
reservoir operation: a case study in a tributary
of the Three Gorges Reservoir, China[J]. Environmental
Monitoring and Assessment, 2016, 188:153.
[40] Reynolds C. Ecology of phytoplankton[M]. Cambridge:
Cambridge University Press, 2006.
[41] Reynolds CS, Huszar V, Kruk C, et al. Towards a
functional classification of the freshwater phytoplankton
[J]. Journal of Plankton Research, 2002, 24:417-428.
[42] Padisák J, Crossetti LO, Naselli-Flores L. Use and
misuse in the application of the phytoplankton
functional classification: a critical review with
updates[J]. Hydrobiologia, 2009, 621:1-19.
[43] 周 利,王茂伟,高静思,等. 氮、磷浓度对伪鱼腥藻
生长及光合生理的影响[J]. 环境工程, 2020, 38:
110-116.
[44] 韦洁琳,崔玉静,李亦真,等. 南水北调东线山东境内
调蓄湖库蓝藻种群分布特征及环境驱动因子[J]. 环
境科学,2023,44:6137-6148.
[45] 谭好臣,王瑗媛,陈阳亮,等. 多水源水库浮游植物优
势种生态位及种间联结性动态分析:以山东峡山水
库为例[J]. 湖泊科学,2023,35:844-853.
[46] Xu H, Brookes J, Hobson P, et al. Impact of copper
sulphate, potassium permanganate, and hydrogen
peroxide on Pseudanabaena galeata cell integrity,
release and degradation of 2-methylisoborneol[J].
Water Research, 2019, 157:64-73.
[47] 陈贝怡,周 睿,观文娜,等. 青岛市自来水和水源地
自然水中嗅味物质的相关性研究[J]. 给水排水,
2023,59,523-528.
猜你喜欢
山东水利(2020年12期)2021-01-18 05:59:04
河南水利年鉴(2020年0期)2020-06-09 05:43:42
河南水利年鉴(2020年0期)2020-06-09 05:43:34
中华建设(2019年8期)2019-09-25 08:26:18
水利技术监督(2017年6期)2017-12-19 13:28:37
河南水利年鉴(2017年0期)2017-05-19 02:29:37
水科学与工程技术(2016年2期)2016-07-10 10:19:57
水科学与工程技术(2016年2期)2016-07-10 10:19:57
水利信息化(2015年5期)2015-12-21 12:54:40
汉语世界(2015年2期)2015-03-26 09:19:01
