
不同品质类型小麦淀粉积累动态及其面包、面条加工品质研究
2024-07-13 10:06:27郭宪峰邓红成马栋申浩巨伟王灿国訾妍
中国种业 2024年6期
郭宪峰 邓红成 马栋 申浩 巨伟 王灿国 訾妍

摘要:面食在食品工业中占有重要地位,面包和面条主要由小麦粉制成,因其感官吸引力、适口性、低成本和储存稳定性而在世界各地广受欢迎。淀粉在决定小麦的理化性质和蒸煮品质方面起着非常重要的作用,其含量及积累动态对小麦的加工品质有重要影响。以6类12个不同面包和面条品质的小麦为试验材料,测定不同时期各品种籽粒中的总淀粉含量、直链淀粉含量、支链淀粉含量的变化,对11个淀粉合成酶相关基因进行表达量的测定。结果表明,开花24d后优质品种的直链淀粉含量明显低于劣质品种;面条优质小麦花后24d支链淀粉含量迅速增加,开花后36d优质品种的支链淀粉较高;开花24d后面包劣质小麦总淀粉含量最高,面包优质品种最低,其他类型品种的平均总淀粉含量差异不大;面包优质和面包面条兼优小麦的SBEI在灌浆后期表达量较高,面包优质小麦SBEIIb在灌浆中期的表达量较高,与优质小麦支链淀粉含量较高相吻合;直链淀粉含量及直支比低的小麦品种面粉的面包、面条品质较好。在面包、面条加工过程中,通过降低淀粉中直链淀粉的含量能有效提高加工品质,直链淀粉含量/支链淀粉含量比值可以作为评价面包、面条品质的指标,为今后的品质改良提供参考。
关键词:小麦;面包;面条;品质;淀粉含量;淀粉合成基因;直支比
Study on the Starch Accumulation Dynamics in Wheat of Different Quality Types and the Processing Quality of Bread and Noodles
GUO Xianfeng1,DENG Hongcheng2,MA Dong1,SHEN Hao3,JU Wei4,WANG Canguo4,ZI Yan4
(1Mudan District Development and Protection Center of Seed Resources,Heze 274000,Shandong;2College of Agronomy,Qingdao Agricultural University,Qingdao 266109,Shandong;3Shandong Medicine Technician College,Tai?an 271000,Shandong; 4Crop Research Institute,Shandong Academy of Agricultural Sciences/National Engineering Research Center for Wheat and Maize/
Key Laboratory of Wheat Biology and Genetic Breeding in Northern Huang-Huai Region,Ministry of
Agriculture and Rural Affairs/Shandong Wheat Technology Innovation Center,Jinan 250100)
小麦是世界第一大粮食作物,是人类重要的粮食资源,其种植面积广泛。亚洲是主要的小麦产区,而中国是种植面积较大的国家之一[1]。近年来,随着人口的不断增长和人民生活水平的提高,食品安全愈加受到人们的重视。面条作为我国的传统面食制品,是我国不可缺少的主食,面包因其独特的口感与便捷的食用方式被越来越多的年轻人所接受。面包、面条等面食需求量随着社会经济的发展逐渐增加,不仅加大了小麦的消费需求,同时也对其品质提出了更高的要求。
蛋白质和淀粉是小麦籽粒的主要成分,是影响面粉理化性质和食品品质的主要因素。淀粉占小麦籽粒干重的65%~70%,对加工品质具有显著的影响。淀粉由直链淀粉(20%~30%)和支链淀粉(70%~80%)组成,直链淀粉的相对含量、支链淀粉的相对含量和直链淀粉与支链淀粉含量的比值(直支比)影响淀粉糊化特性、热力学特性、凝胶特性和蒸煮品质[2-3]。直链淀粉含量较低的面粉,面条和馒头品质更好[4-5],支链淀粉能有效延长面包和面条的货架寿命[6]。直链淀粉含量越高,面包品质越差,口感较差,老化速度更快[7-9],面条与水的结合能力降低,使其失去弹性并硬化,直链淀粉含量最少的糯小麦粉制作出的面条在口感、色泽及品质上较优越。
淀粉的生物合成是一个复杂的过程,需要一系列合成酶作用才能完成,主要包括腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)、结合态淀粉合成酶(GBSS)、可溶性淀粉合成酶(SSS)和淀粉分支酶(SBE),其编码基因的表达量在一定程度上可以反映淀粉合成情况。AGPase合成基因包括SSUI、SSUII、LSUII,GBSS合成基因包括GBSSI、GBSSII,SSS合成基因包括SSSI、SSSII、SSSIII,SBE合成基因包括SBEI、SBEIIa、SBEIIb。小麦灌浆期不同时间淀粉合成酶基因的表达量不同,对其进行研究可以进一步明确小麦淀粉合成的主要过程及对品质形成产生影响的主要生长阶段,为小麦品种的品质改良奠定基础。
开展小麦品质研究,发展优质专用、优质兼用小麦的生产是我国优质、高效农业的主要研究内容之一[10]。前人对淀粉含量的积累、淀粉合成酶基因表达量的变化及对面包、面条加工品质影响的系统研究较少,因此本试验选取6类12个不同面包、面条特性的小麦品种进行研究,分析了不同品质类型的小麦淀粉含量及积累特征,以及淀粉合成酶在不同时期的基因表达量,并探讨这些特性与面条和面包加工品质之间的关系,为小麦品质育种及改良提供参考依据。
1 材料与方法
1.1 试验材料 供试品种为6类12个小麦品种,分别是面包优质小麦[济麦4072(JM4072)、淄麦12(ZM12)],面条优质小麦[济麦19(JM19)、周麦18(ZM18)],面包劣质小麦[泰农18(TN18)、青丰1号(QF1)],面条劣质小麦[鲁麦1号(LM1)、鲁麦19(LM19)],面包面条均优小麦(包条兼优)[济麦20(JM20)、郑麦366(ZM366)],面包面条均劣小麦(包条均劣)[鲁麦14(LM14)、新麦18(XM18)]。
1.2 试验设计 试验于2017-2018年在山东省农业科学院实验站(36°42′N,117°05′E)进行。土壤为壤土,大田土壤有机质含量为10.9g/kg,全氮含量1.33g/kg,速效氮含量80.5mg/kg,速效磷含量33.4mg/kg,速效钾含量118.9mg/kg。
试验采用随机区组设计,每个品种播种密度均为300万/hm2,行距25cm,小区面积6.0m2(4.0m×1.5m),3次重复。2017年10月播种,开花后灌浆期间取籽粒进行淀粉含量及淀粉酶合成基因表达量测定,2018年6月全田收获,将籽粒晾晒、储存、磨粉并制作成面包、面条后测定加工品质指标,田间水肥管理及病虫害防治均按高产田常规管理措施进行。
1.3 测定项目与方法
1.3.1 籽粒总淀粉含量、直链淀粉含量及支链淀粉含量的测定 开花期标记同一天开花的麦穗,花后每隔6d每个小区取30~50个麦穗进行籽粒取样,共取5次。将籽粒105℃杀青40min,80℃恒温烘干,过60目筛后精确称量0.1000g装入小试管。利用双波长法测定小麦籽粒总淀粉含量、直链淀粉含量及支链淀粉含量[11-12]。淀粉含量积累速率用2次测定的淀粉含量差除以天数表示。
1.3.2 小麦淀粉合成酶相关基因表达量的测定 取上述花后同一时期的籽粒作为样品,参照天根总RNA提取试剂盒(RNAsimple Total RNA Kit)操作方法,提取小麦灌浆期各个阶段籽粒的总RNA。采用Dnase I(RNase-free)消化所得RNA样本中的残余DNA,方法参照宝生物工程(大连)有限公司的试剂使用说明。参照宝生物工程(大连)有限公司(TaKaRa)的反转录试剂盒(RNA LA PCRTM Kit(AMV)Ver.1.1)操作方法进行RNA的反转录。荧光定量PCR引物设计利用Beacon Designer 7软件(表1),目的基因序列来自NCBI中小麦EST数据库,Actin基因为内参基因,引物设计扩增区段通过NCBI中Blast同源比对功能进行选择,并对设计的引物特异性进行评价,委托北京六合华大基因科技股份有限公司合成引物。荧光定量PCR反应参照TaKaRa试剂盒(SYBYR Premix Ex TaqTM)操作方法。
1.3.3 面包、面条的制作与品质评价 收获的小麦籽粒放置2个月后制粉。根据AACC26-10的方法进行润麦(15%含水量),润麦24h。籽粒磨粉采用Quadrumat Junior试验磨粉机(Brabender公司,德国)(AACC26-21A),面粉熟化2周后用于指标的测定。
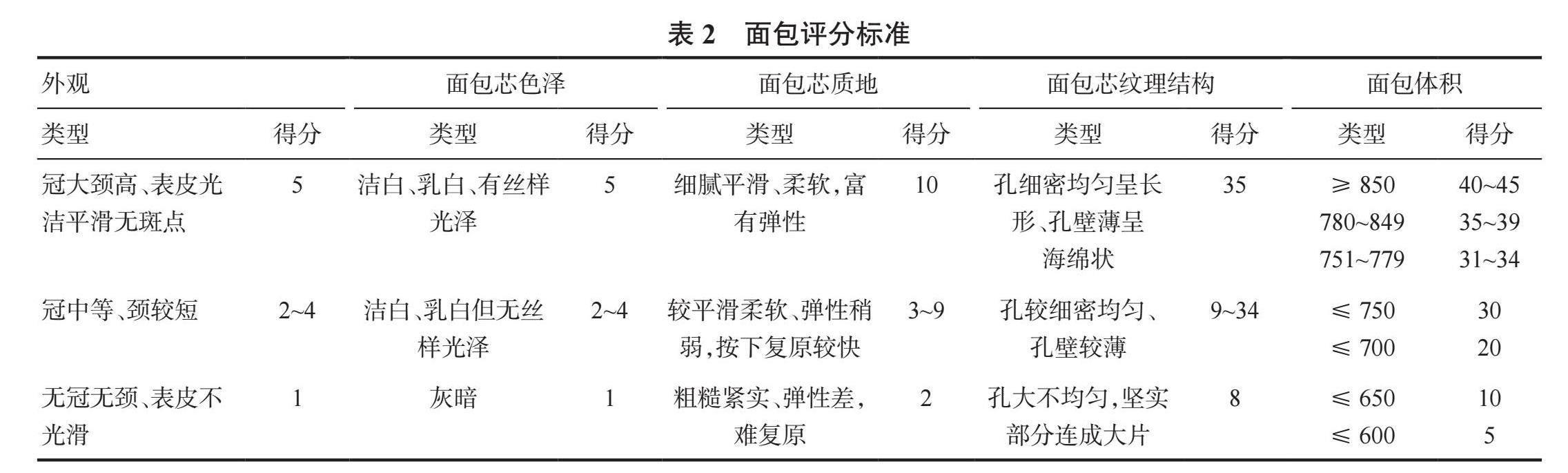
面包的制作(100g面粉,14%含水量)和品质评估按照田纪春[11]的方法略有改进。5名经过训练的小组成员品尝后进行综合评分,总分为100分,品质指标包括面包体积(45分)、外观(5分)、面包芯色泽(5分)、面包芯质地(10分)、面包芯纹理结构(35分),评价标准见表2。
面条的制作和品质评价按照田纪春[11]的方法略有改进。5名经过训练的小组成员品尝后进行综合评分,总分为100分,品质指标包括色泽(20分),表观状况(10分),适口性(20分),粘弹性(30分),光滑性(15分),食味(5分)。称取50g干面条于500mL沸水中煮3min后,每间隔0.5min品尝一次,面条保持微沸状态7min后,观察面条是否断条、是否耐煮,并按照标准逐项打分,同时取出一部分熟面条于白瓷盆中,与对照样品进行色泽和表观评价,评价标准见表3。
1.4 数据处理与分析 采用Microsoft Excel软件对数据进行统计分析,差异显著性用LSD法表示。
2 结果与分析
2.1 不同品质类型小麦淀粉含量积累动态分析 由图1可以看出,不同品种小麦开花后籽粒直链淀粉、支链淀粉以及总淀粉含量积累均呈快速增长的趋势,但不同品种相同时期的直链淀粉、支链淀粉和总淀粉含量积累存在明显差异,青丰1号开花后36d直链淀粉含量和总淀粉含量在所有品种中最高。
直链淀粉含量在开花后24d之前增长较快,24d以后基本上呈缓慢上升趋势;在开花6d后,郑麦366的直链淀粉含量在所有品种中最高,达到3.404%,最低的为济麦19,仅为1.016%;开花后36d直链淀粉含量最高的是青丰1号和鲁麦1号,达24.600%,周麦18含量最低,为18.500%。支链淀粉的增长曲线与直链淀粉相比略微平缓,增长速率较稳定,在整个积累过程中品种间差异不明显;开花后6d支链淀粉含量最高的为鲁麦19(18.227%),最低的是泰农18(5.413%),而开花后36d支链淀粉含量最高的是周麦18和济麦20,达48.500%,最低的是鲁麦1号,为43.800%。从总淀粉含量的变化来看,开花后18d以前品种之间的差异较明显,18d之后差异变小,24d前快速积累,24d后缓慢上升;开花后6d总淀粉含量最高的是鲁麦19,达到20.521%,最低的泰农18只有6.438%;开花后36d总淀粉含量最高的品种是泰农18,高达69.700%,最低的周麦18为67.000%。
根据不同面包、面条品质类型小麦总淀粉及其组成成分的含量,6种品质类型的小麦又可以分为2个大组,即优质品种(面包优质、面条优质、包条兼优)和劣质品种(面包劣质、面条劣质、包条均劣)。对6个品质类型小麦的淀粉含量积累研究表明,各类型间开花后24d之前淀粉及其组分含量差异不明显,24d后优质品种的直链淀粉含量明显低于劣质品种;面条优质小麦花后24d以后支链淀粉含量迅速增加,花后36d时成为所有类型中支链淀粉含量最高的,包条均劣小麦灌浆中期(花后18~24d)支链淀粉含量最低,开花后36d面条劣质小麦和包条均劣小麦的支链淀粉含量较低;开花后24d面包劣质小麦总淀粉含量最高,最低的是面包优质小麦,其他类型小麦的总淀粉含量差异不大(图2)。
对6个品质类型小麦的淀粉含量积累速率分析(图3),花后12d前,所有类型小麦的直链淀粉含量积累速率均明显上升,之后除面条优质和面包劣质小麦下降,其他类型均继续增加,面包劣质小麦在花后24d达到峰值;支链淀粉含量积累速率差异较大,面包优质、包条均劣及面条劣质小麦先降低后升高再降低,其他类型为先升高后降低;总淀粉含量积累速率除面包优质小麦,其他类型呈先上升后下降趋势,面条优质与包条兼优小麦在花后12d总淀粉积累速率最大,之后迅速下降。
从不同品质类型小麦直链淀粉含量与支链淀粉含量比值(直支比)可知,花后6d包条兼优小麦直支比最高,包条均劣小麦直支比最低;开花24d后,优质品种直支比比劣质品种低;成熟期(花后36d)面条优质小麦直支比最低,面条劣质小麦直支比最高(图4)。
2.2 不同品质类型小麦淀粉合成关键酶基因表达的动态分析 对6个类型小麦的淀粉合成酶基因表达量的分析表明,籽粒灌浆过程中3种AGPase合成基因(SSUI、SSUII、LUSII)相对表达量均呈单峰变化模式,基本上约在花后18d达到峰值(图5)。SSUI基因在不同品质类型小麦中均呈单峰曲线变化,在花后18d达到最高峰;包条兼优小麦在灌浆前期表达量(花后18d)居6个类型中最高,但花后18d之后呈现逐渐下降的趋势;与其他几类小麦品种不同的是,面条劣质小麦在花后24d表达量最低,之后略有增加。SSUII基因在6类小麦中的表达量均先上升后下降,面包优质小麦在灌浆前期表达量明显高于其他5类小麦品种。LSUII基因除面条劣质小麦峰值在花后24d,其他5类小麦均在花后18d达到峰值,包条均劣小麦表达量在花后12d略有下降。
2种GBSS合成基因中GBSSI基因的表达情况呈波动变化模式,在花后18d达到高峰,但6个类型小麦的GBSSI基因的表达明显不同,有的类型峰值非常明显,如面包劣质小麦,有的类型峰值不明显,如包条均劣小麦;GBSSII基因在花后18d达到高峰,包条均劣小麦在花后12d出现低峰,面条优质小麦峰值不明显。
从整体来看,3种SSS合成基因(SSSI、SSSII、SSSIII)在籽粒灌浆过程中的变化趋势明显不同。SSSI基因呈单峰变化模式,包条均劣小麦在花后12d达到峰值,其他类型小麦在花后18d达到峰值;SSSII基因有的类型峰值非常明显,面包劣质小麦在花后18d表达量明显高于其他类型,但有的类型峰值不明显,如面条劣质小麦和包条均劣小麦;SSSIII基因表达量在不同类型小麦籽粒灌浆过程中呈下降或平缓的趋势,基本上在花后12d达到峰值,随后表达量下降或略有上升。
3种SBE基因(SBEI、SBEIIa、SBEIIb)在籽粒灌浆过程中呈单峰变化趋势,并均在花后18d达到峰值。面包优质和包条兼优小麦的SBEI基因在灌浆后期表达量较高,包条均劣和面条劣质小麦表达量较低;包条均劣小麦在花后24d之前SBEIIa的表达量低于其他5种类型;包条兼优小麦在灌浆前期SBEIIb表达量最低,在花后30d增至最高。
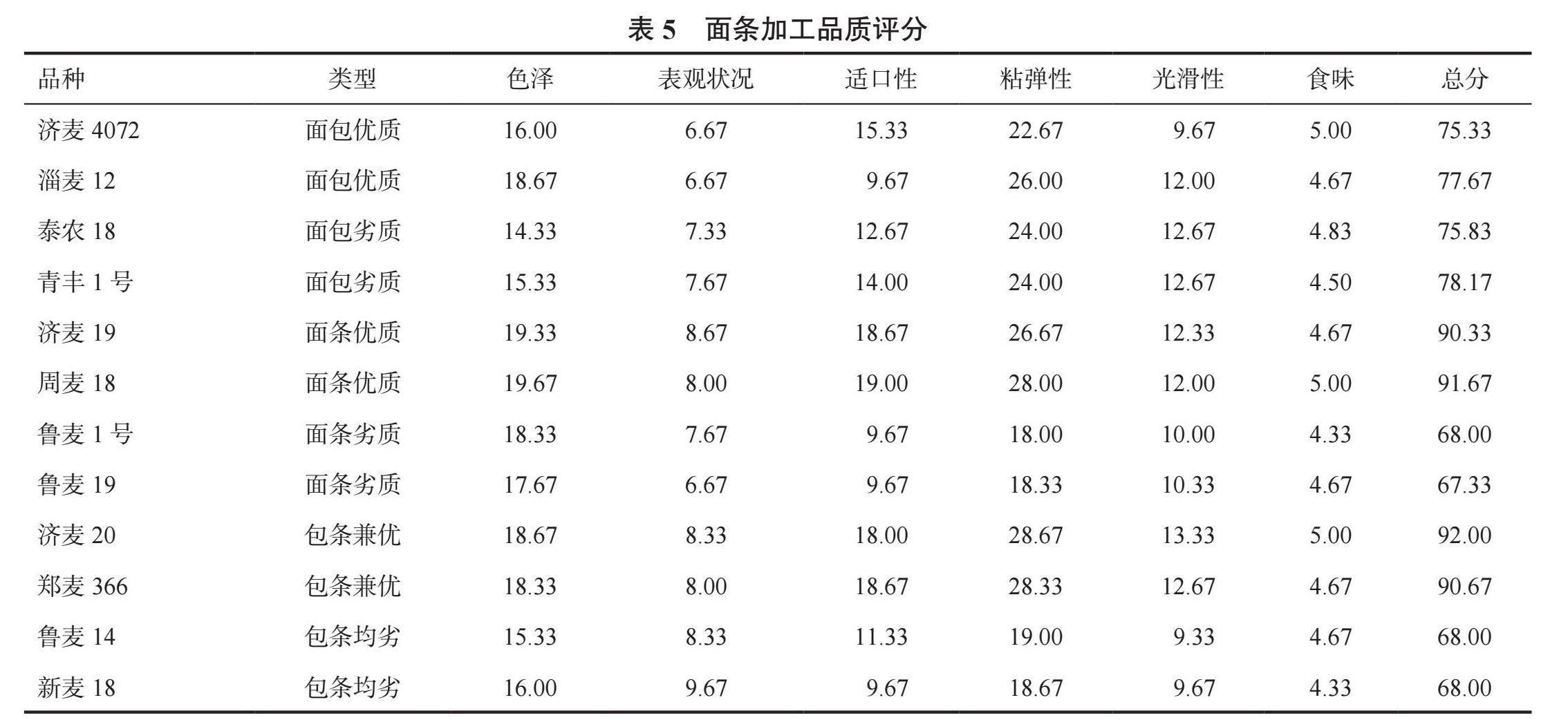
2.3 不同品质类型小麦面包、面条加工品质分析
2.3.1 不同面包、面条品质小麦品种面包品质评价 不同类型品种小麦的面包品质评价得分如表4所示,不同品种小麦在面包品质上的表现差异明显,面包优质小麦与包条兼优小麦在面包品质的评分上都处于90分以上,面包优质小麦的评分最高;面包劣质小麦与包条均劣小麦面包品质评分均低于60分,面包劣质小麦评分最低;而面条优质/劣质的品种在面包品质中的表现也相对较好,低于面包优质和包条兼优的品种,但优于面包劣质和包条均劣品种。
12个品种中,面包体积最大的是济麦20,外观较好的是济麦4072,面包芯色泽最好的是济麦4072和淄麦12,面包芯质地最好的是济麦20,面包芯纹理结构得分最高的是淄麦12,面包品质综合表现最好的是淄麦12,总分在95.00分,其次是济麦4072,总分为93.67分,均为面包优质小麦。该类型品种在花后36d的总淀粉含量最低,直链淀粉含量较低。
2.3.2 不同面包、面条品质小麦品种面条品质评价 由表5可知不同类型品种小麦的面条品质表现,面条优质小麦与包条兼优小麦的面条品质评分都在90分以上,面条优质小麦在色泽、表观状况和适口性方面优于包条兼优小麦;面条劣质和包条均劣小麦面条品质评分都低于70分,差异不明显;面包优质与面包劣质两种类型小麦面条评分均在70分以上,面包优质小麦的食味较好。
12个品种中,面条色泽最好的是周麦18,表观状况较好的是新麦18,适口性最好的是周麦18,粘弹性最好的是济麦20,光滑性得分最高的是济麦20,食味较好的是济麦4072、周麦18和济麦20,面条品质综合表现最好的是包条兼优小麦济麦20,总分在92.00分,其次是周麦18,总分为91.67分。周麦18在花后36d的直链淀粉含量及直支比最低,其次为济麦20,由此可以看出,直链淀粉含量及直支比低的小麦品种面粉的面条品质较好。
3 讨论与结论
淀粉作为小麦的重要组分之一,其含量与积累特性对面包、面条品质的影响尤为明显,本文通过对不同小麦中的直链淀粉、支链淀粉及总淀粉含量积累动态及淀粉合成酶基因表达量的研究,分析了淀粉含量及积累动态对不同类型小麦面包、面条品质的影响。
小麦淀粉的含量对面包面条的品质具有较大影响,其中直链淀粉和支链淀粉的不同比例对面包面条品质指标影响较大。直链淀粉含量过低,面团发粘,对面包的体积和质地有较大影响,一定程度上能增加面包体积,但会引起气泡不均匀和结构差等问题[13-14]。直链淀粉含量较低的面粉制成的面包在放置一段时间后仍然保持松软,加热后食用口感较好[15]。降低直链淀粉的含量可以提高面团膨胀能力,在面条制作过程中降低糊化凝胶的硬度、增强形变能力,使面条柔软并充满弹性[16],进一步影响面条的色泽、表观状况、适口性、粘弹性、光滑性和食味品质,同时也对面包体积、外观、色泽、质地、纹理结构等具有一定的影响[12,17-18]。支链淀粉含量升高可以提高淀粉膨胀势,进一步使面条质地更松
软[19]。本研究通过对比不同品种小麦的淀粉含量及积累过程,发现不同品种小麦在直链淀粉含量上的差异大于支链淀粉含量,而总淀粉含量差异较小,直链淀粉与支链淀粉的比例差异较大;同时通过观察不同类型小麦在直链淀粉、支链淀粉及总淀粉含量上的差异可以看出,直链淀粉的含量在面包、面条品质中的影响最明显,优质品种的直链淀粉含量远低于劣质品种中的含量,直支比低;而支链淀粉的积累在各类品种中相差不大,因此在面包、面条加工过程中,通过降低淀粉中直链淀粉的含量能有效提高加工品质。直链淀粉含量/支链淀粉含量的比值可以作为评价面包、面条加工品质的指标,为今后的品质改良提供参考。
基因调控淀粉酶合成淀粉是一个复杂的过程,合成底物的多少、调控基因表达量的多少、淀粉酶的多少均会对淀粉的合成产生影响,从而进一步影响小麦品质。本研究中除SSSIII相对表达量花后12d以后呈持续下降或略有上升的趋势外,其余的淀粉合成相关酶基因表达量基本上呈单峰曲线,与前人研究结果一致[20]。研究表明,抑制SBE合成基因表达,会降低支链淀粉的合成能力,直链淀粉的相对含量增加[21]。本试验中面包优质和面包面条兼优小麦的SBEI在灌浆后期表达量较高,面包优质小麦SBEIIb在灌浆中期的表达量较高,与优质小麦支链淀粉含量较高相吻合。但是仅仅从基因表达量仍无法直接分析小麦品质的优劣,仍需进一步结合品质数据,这也可能与试验样本过少有关,仍需进一步研究。
参考文献
[1]赵广才,常旭虹,王德梅,陶志强,王艳杰,杨玉双,朱英杰.小麦生产概况及其发展.作物杂志,2018(4):1-7
[2] Blazek J,Copeland L.Pasting and swelling properties of wheat flour and starch in relation to amylose content.Carbohydrate Polymers,2008,71(3):380-387
[3] Kaur A,Shevkani K,Katyal M,Singh N,Ahlawat A K,Singh A M.Physicochemical and rheological properties of starch and flour from different durum wheat varieties and their relationships with noodle quality.Journal of Food Science and Technology,2016,53(4):2127-2138
[4] Guo Q,He Z,Xia X,Qu Y,Zhang Y.Effects of wheat starch granule size distribution on qualities of Chinese steamed bread and raw white noodles.Cereal Chemistry,2014,91(6):623-630
[5] Singh S,Singh N,Isono N,Noda T.Relationship of granule size distribution and amylopectin structure with pasting,thermal,and retrogradation properties in wheat starch.Journal of Agricultural and Food Chemistry,2010,58(2):1180-1188
[6]刘爱峰,宋建民,赵振东,刘建军,吴祥云,李豪圣,刘广田.糯小麦配粉对面团流变学特性和面包烘烤品质的影响.中国农业科学,2004,37(6):902-907
[7]盛婧,郭文善,朱新开,封超年,彭永欣.不同类型专用小麦籽粒淀粉及其组分积累动态.扬州大学学报:农业与生命科学版,2006,27(2):31-35
[8] Li C Y,Zhang R Q,Fu K Y,Li C,Li C.Effects of high temperature on starch morphology and the expression of genes related to starch biosynthesis and degradation.Journal of Cereal Science,2017,73(8):25-32
[9] Wang Z B,Li W H,Qi J C,Shi P C,Yin Y G.Starch accumulation,activities of key enzyme and gene expression in starch synthesis of wheat endosperm with different starch contents.Journal of Food Science and Technology,2014,51(3):419-429
[10]纪建海,王彦霞.我国传统食品面条研究进展.粮食加工,2013,38(1):9
[11]田纪春.谷物品质测试理论与方法.北京:科学出版社,2006
[12]何照范.粮油籽粒品质及其分析技术.北京:中国农业出版社,1985
[13]付蕾,田纪春,高吉刚,孙华.抗性淀粉与小麦粉配粉对面包加工品质的影响.中国粮油学报,2010,25(12):6-9,23
[14]付蕾,田纪春,盛锋,李长城.抗性淀粉对面条加工品质的影响.中国粮油学报,2011,26(1):20-24,34
[15] Morita N,Maeda T,Miyazaki M,Yamamori M,Miura H,Ohtsuka I.Dough and baking properties of high-amylose and waxy wheat flours.Cereal Chemistry,2002,79(4):491-495
[16]王红日,王利民,戴双,刘爱峰,曹新有,宋健民.小麦淀粉理化特性遗传改良研究进展.山东农业科学,2013,45(8):137-140
[17]申浩.小麦面包面条优质兼用的淀粉理化特性研究.泰安:山东农业大学,2017
[18]刘锐,魏益民,邢亚楠,张波,张影全.小麦淀粉与面条质量关系的研究进展.麦类作物学报,2013,33(5):1058-1063
[19] Baik B K,Lee M R.Effects of starch amylose content of wheat on textural properties of white salted noodles.Cereal Chemistry,2003,80(3):304-309
[20] Stamova B.S,Laudencia-Chingcuanco D,Beckles D M.Transcriptomic analysis of starch biosynthesis in the developing grain of hexaploid wheat.International Journal of Plant Genomics,2009:407-426
[21] Jobling S A,Jarman C,Teh M M,Holmberg N,Blake C,Verhoeyen M E.Immunomodulation of enzyme function in plants by single-domain antibody fragments.Nature Biotechnology,2003,21:77-80
(收稿日期:2024-04-08)
猜你喜欢
少年博览·小学低年级(2020年10期)2020-11-06 07:32:53
时代邮刊·下半月(2020年7期)2020-07-20 06:39:48
少儿美术(快乐历史地理)(2019年5期)2019-09-10 08:34:30
儿童时代·幸福宝宝(2019年1期)2019-03-31 15:00:22
作文周刊·小学一年级版(2017年11期)2017-06-20 00:16:48
现代园艺(2016年17期)2016-10-17 06:39:29
现代园艺(2016年17期)2016-10-17 06:31:18
今传媒(2016年9期)2016-10-15 22:48:38
投资者报(2016年33期)2016-09-30 18:19:51
小学生导刊(2016年34期)2016-08-22 12:30:36
