
中国近海渔业资源的结构与质量研究
2024-07-13 04:48:59张瑛李忠辉张力
中国人口·资源与环境 2024年5期
张瑛 李忠辉 张力


摘要 海洋渔业资源对于保障食品安全、维护生态系统稳定、促进经济发展等方面起着关键作用,科学管理和可持续利用近海渔业资源,对于推动经济与环境协调发展至关重要。为了探究中国近海渔业资源的结构与质量变化情况,该研究基于1990—2020年中国近海捕捞统计数据和海表温度(sea surface temperature, SST)变化数据,通过计算分析中国近海各渔区(渤黄海渔区、东海渔区和南海渔区)渔获物的平均营养级(mean trophic level,MTL)、各渔区渔业均衡指数(fishing?in?balance index,FIB指数)与人为捕捞活动、SST变化之间的关联性,探究人为捕捞活动、SST变化对中国近海渔业资源结构与质量变化、渔业生态系统健康的影响。结果表明:过去30年间,中国近海各渔区的渔获量和FIB指数均呈现先上升后下降的趋势,渤黄海和东海渔区的MTL呈现显著下降趋势,南海渔区的MTL呈现微弱增长趋势。人为过度捕捞已影响到渤黄海和东海渔区的渔业资源结构与质量,导致低营养级鱼类占比逐渐升高,渔业资源质量显著降低,但对南海渔区影响不大。受到气候变化影响,在渤黄海渔区5—6月和东海渔区9—11月,海温增暖抑制高营养级鱼类生长,导致渔业资源出现结构向低营养级鱼类变化、质量大幅下降的趋势,但在南海渔区,海温变化对MTL没有影响。人为捕捞对中国近海渔业生态系统健康影响不大,主要是由于渔获量的增加起到了一定的平衡作用。海温变化对渤黄海、东海渔区的渔业生态系统健康影响也较小,南海秋季海温增暖则有利于其渔业生态系统健康发展。基于研究结果,作者提出了改善中国近海各渔区渔业资源结构,提高渔业资源质量,提升渔业生态系统健康的相关建议,为中国近海渔业资源的可持续利用提供了管理借鉴。
关键词 近海;渔业资源;人为捕捞;气候变化
中图分类号 P74;S931 文献标志码 A 文章编号 1002-2104(2024)05-0058-11 DOI:10. 12062/cpre. 20231021
海洋渔业资源是一种重要的自然资源,为人类提供了高质量的食物来源,并且为将近70%的世界人口提供了40%的蛋白质[1]。海洋渔业资源种类繁多,包括鱼类、甲壳类、贝类、藻类、足类和其他类,其中鱼类和甲壳类的品种众多,数量大。海洋渔业资源的合理利用对维护海洋资源的生态平衡、推动海洋经济朝着健康可持续的方向发展有着举足轻重的意义[2-3]。中国大陆海岸线绵长,同时拥有渤海、黄海、东海和南海等边缘海,海洋渔业资源丰富。但从20世纪中期开始,由于市场上对高质量渔业资源的需求不断扩大,许多先进的捕捞技术也就应运而生,捕捞力度不断加大,加之气候变化、海洋环境污染、管理制度等原因,中国近海渔业资源可持续利用面临巨大的挑战[4-9]。因此,对海洋渔业资源动态变化的有效掌握,将有利于及时发现其开发利用过程中出现的问题,推动中国海洋渔业健康发展[10]。本研究聚焦人为捕捞和气候变化双重影响,对中国近海渔业资源结构与质量变化进行对比分析与归因研究,以期为中国近海渔业资源高质量管理提供决策参考。
1 文献综述
近海渔业资源是海洋渔业资源重要的组成部分。近10年,中国近海捕捞渔业迅速发展的同时,也导致了其与资源环境冲突问题愈来愈严重。一方面,由于人为影响,随着捕捞技术的提升和捕捞政策的开放,捕捞强度呈上升趋势,且居高不下,致使近海捕捞长期处于饱和的危险状态。同时,捕捞强度增加使得近海渔业资源衰退严重,近海主要经济鱼类资源大幅衰减[4,7]。另一方面,气候变化对渔业资源波动也影响深远。研究表明,气候变化主要通过改变海水温度、上层洋流、海洋层结等过程,影响海水酸碱度、营养盐分布等状况,进一步对渔业资源的多时空尺度变化产生影响[8,11]。
针对目前的海洋渔业资源利用现状,国内外学者从不同的方向进行了研究[12-18]。已有研究在影响因素的考量方面,就人为捕捞因素对渔业资源可持续利用产生的影响,从宏观系统角度开展了较多研究。刘子飞[14-15]分析了中国近海捕捞渔业管理政策对渔业资源状态的影响,指出随着专业捕捞人员、捕捞渔船功率吨位及非选择性渔具的不断增加,中国近海渔业资源减少是很难避免的,不合理的渔具使用和超强度的捕捞会破坏渔业资源生存的“三场一通道”,对渔业资源的可持续利用形成负面影响。这些研究从宏观角度揭示了高强度的人为捕捞导致了中国近海渔业资源减少的现象,但尚未就人为捕捞对中国近海渔业资源结构与质量进行量化分析。
为衡量渔业资源结构与质量变化,国内外相关研究中广泛使用渔获物营养层级变化作为重要的参考指标[2]。渔获物平均营养级(mean trophic level,MTL)主要用来反映渔获物种群结构的变化情况[19-20]。当高营养级的捕食性鱼类被过度捕捞时,平均营养级会降低,可能会导致整个渔业生态系统平衡被打破。除MTL 外,Pauly等[21]又提出渔业均衡指数(Fishing?in?Balance index,FIB指数),主要聚焦食物链中底层的小型食饵鱼类的生物量。这些小型生物是大型捕食性鱼类的主要食物来源,过度捕捞会影响到上层食物链的鱼类。MTL和FIB指数结合可以更全面反映渔业生态系统中不同营养级的动态变化[22-24]。国内外学者针对特定海域渔获物营养级变化开展了卓有成效的研究。例如,吴佳颖等[12]对渤黄海渔区主要渔获物的平均营养级进行了研究,指出渤黄海MTL在近50年间整体呈下降趋势,中低营养级渔获物的渔获量增加,而高营养级渔获物的渔获量减少。李继龙等[13]根据中国渔船渔获统计数据,对中国东部海域和南部海域的渔业资源利用水平进行了研究,指出东部海域MTL在近30年间呈现下降趋势,其渔业资源利用已经呈现出一种不可持续性,而南部海域MTL无显著变化趋势,并未出现渔业资源利用的衰退现象。Milessi等[25]聚焦乌拉圭海域,通过分析乌拉圭渔场60种渔获物的营养级,发现MTL以每10年0. 28的速度下降,乌拉圭渔场渔业资源利用呈现出不可持续性。
除了人类的捕捞活动外,气候变化是影响渔业资源整体结构与质量的另一个关键因素。气候变化对MTL变化的影响存在持续性,其关键影响路径是通过改变海洋环境要素进而对渔业资源结构与质量产生影响。海温与海洋物种的分布、丰度及其变化有着显著的联系,适宜的海温有利于海洋生物的新陈代谢,促进海洋生物的生长和繁殖[26],异常的海温会影响海洋生物的生理功能,导致新陈代谢紊乱,从而影响其生长发育[27]。而海洋生物的生长发育又影响着自身的体长,一般来说,海洋生物的体长与自身营养级之间存在着联系[28-29]。因此,气候变化通过影响海温,进而对海洋生物的体长和营养级产生影响。陈爽等[30]针对东北大西洋海域的MTL和多样性指数进行了探究,分析结果表明,这两个指标与海洋环境因子的变化有着显著联系,随着海温和海水盐度不断升高,多样性指数和MTL呈现下降趋势。另外,海温变化与海洋生物的资源量变化,以及群落格局的代际变化也有联系[31-32]。基于气候变化的不断加剧,其所带来的海洋环境变化对渔业资源的影响是渔业管理重要的研究方向之一[33]。对于中国近海,海温呈现快速增暖趋势,其速率大约是全球平均的3~5倍[11],这种快速增温如何影响渔业资源结构与质量亟待评估。
本研究围绕人为捕捞和气候变化对中国近海渔业资源结构与质量变化的双重影响,利用1990—2020年的国内外相关数据系统分析中国近海3个渔区(渤黄海渔区、东海渔区、南海渔区)的MTL和FIB指数的变化趋势,重点分析捕捞渔船不同作业类型、海温变化对各渔区MTL和FIB指数的影响,进一步对各渔区的渔业资源结构与质量变化进行对比分析与归因研究。本研究中近海渔业资源主要考虑海水中具有经济价值的动物,以鱼类和甲壳类为主。
2 数据来源与研究方法
2. 1 数据来源
本研究所采用的渤黄海、东海、南海1990—2020年的渔获量数据,包括按品种和渔具分类的各地区海洋捕捞产量、按海域划分的各海域海洋捕捞产量均来自《中国渔业统计年鉴》国内海洋捕捞生产统计。结合数据的可获取性,分别利用辽宁省、河北省、天津市、山东省、江苏省的渔获统计数据对渤黄海渔区的渔获物进行分析;利用浙江省、上海市、福建省的渔获统计数据对东海渔区的渔获物进行分析;利用广东省、广西壮族自治区、海南省的渔获统计数据对南海渔区的渔获物进行分析。上述渔获物的种类主要包括鳀鱼、小黄鱼、鲐鱼、梭子蟹、鹰爪虾等26种。相关渔获物种类营养级的确定主要借鉴孙明等[34]对辽东湾海域主要渔业生物营养级的研究,白怀宇等[35]对黄海主要鱼类营养级和食性的研究,高春霞等[36]对浙江南部近海主要渔业生物营养级的研究,黄佳兴等[29]对南海中西部渔场主要渔业生物营养级的研究。
中国近海SST数据来自美国国家海洋和大气管理局(NOAA),水平分辨率为1°×1°;水平覆盖范围为105°E~131°E,4°N~40°N;覆盖时段为1990 年1 月—2020 年12月。
2. 2 研究方法
与个体小、寿命短、营养级低的渔业生物相比,个体大、寿命长、营养级高的肉食性渔业生物更容易受到过度捕捞的影响,这会使得海域内整体渔业资源的营养级水平呈下降趋势,而MTL指标可以用来确定某渔区内高营养级渔业资源的衰退情况。同时,FIB指数可以与MTL指标相结合,从而更加全面地评估渔业生态系统的均衡状况。另外,为了确定人为捕捞活动和海温变化对MTL和FIB指数的具体影响,本研究采取一元相关分析方法进行计算分析。
2. 2. 1 计算渔获物平均营养级
渔获物平均营养级概念是由Pauly等[20]首先提出的,主要用来反映渔获物种群结构的变化情况,判断多物种渔业生物的承受力,以衡量渔业生物多样性和可持续性水平,其计算公式为:
另外,借鉴邓景耀等[37]对中国近海渔获物营养等级划分的研究,本研究依据营养级指数将渔获物营养等级分为3 级,即低营养级种类(2. 0~2. 8)、中营养级种类(2. 9~3. 4)和高营养级种类(3. 5~4. 0)。这种划分有助于观测海洋生物营养层级构成变化,高营养级种类渔获量比重的下降,可以帮助确定某海域是否出现了捕捞对象沿着海洋食物网向下移动的现象。
2. 2. 2 计算渔业均衡指数
Pauly等[21]认为,捕捞活动对于渔业生态系统的影响是多方面的,只计算MTL这一个指标的变化,可能会存在“刻意降低食物网”的现象。因此,他又提出了另一个指标,即FIB指数,该指数是用来衡量相对于基准年的总捕捞量与MTL的抵消情况。FIB指数意在考虑渔业转向捕捞低营养级物种时可能出现的捕捞量增加的情况,其计算公式为:
2. 2. 3 一元相关分析
一元相关分析是一种用于研究两个连续型变量之间关系的统计方法,以便于了解变量之间的趋势和相关程度。在这个分析中,主要关注皮尔逊相关系数,该系数的取值范围介于-1到1之间,为正值时表示正相关,负值时表示负相关,而接近于0则意味着较弱的相关性。具体计算公式如下:
为研究人为捕捞活动对渔业资源变化的影响,本研究将统计范围内捕捞渔船作业类型划分为5种,包括选择性渔具(围网、钓具和流刺网)和非选择性渔具(拖网和定置网),将捕捞渔船不同作业类型的渔获量分别与MTL、各营养等级渔获量占比、FIB指数进行一元相关分析。为研究气候变化对渔业资源变化的影响,本研究着重分析中国近海各渔区每个月SST变化趋势对MTL和FIB指数的影响。
3 结果分析
3. 1 中国近海各渔区渔获量、渔获物平均营养级与渔业均衡指数的变化
就渔获量而言,1990—2020年渤黄海和东海渔区的年渔获量呈现基本相同的变化趋势,具体表现为在1990—1998年期间增长迅速,达到历史最高渔获量,之后基本保持稳定,2017年开始下降。南海渔区的渔获量在1990—2005年持续增长,2006—2016年渔获量稳定,之后逐年下降。综合30年间的渔获总量,东海最高,渤黄海次之,南海最低(图1(a))。
就MTL而言,渤黄海渔区的MTL在1990—2002年呈现整体下降趋势,2003—2007年则呈现上升趋势,2007—2010年又逐渐下降,之后稳定在3. 04左右。东海渔区的MTL 在1990—2002 年呈现整体下降趋势,之后稳定在3. 27左右。南海渔区的MTL在1990—2003年呈现微弱增长趋势,2004—2006年逐渐上升,之后稳定在3. 2左右(图1(b))。
就FIB指数而言,渤黄海和东海渔区的FIB指数整体上呈现出了先上升后下降的趋势。具体来说,1990—1998年FIB指数呈现上升趋势,1999—2020年FIB指数呈现下降趋势。在南海渔区,FIB指数总体上呈现上升趋势,2017年后有所下降(图1(c))。
从各营养等级渔获量占比来看,在渤黄海渔区,低营养级渔获量占比最高,基本稳定在40%~50%;中营养级渔获量占比整体呈现上升趋势,在2006年后稳定在33%左右;高营养级渔获量占比呈现下降趋势,由1990年的34%下降至2020年的20%(图2(a))。在东海渔区,高营养级渔获量占比最高,呈现显著的下降趋势,由1900—2001年的52%迅速下降到2002—2020年的40%;中营养级渔获量占比整体呈现显著的上升趋势,由1990—2000年的30%上升到2003—2020年的41%;低营养级渔获量占比最小,基本上稳定在20%左右(图2(b))。在南海渔区,中营养级渔获量占比最高,在1998年前呈现下降趋势,之后稳定在40%;低营养级渔获量占比整体稳定在20%左右;高营养级渔获量占比呈现上升趋势,由1990年的20%上升到2020年的35%(图2(c))。
为进一步分析渔获量与MTL、渔获量与FIB指数之间的关系,本研究分别计算了不同渔区渔获量与MTL、FIB指数的相关系数(表1)。结果显示,渤黄海渔区的渔获量与MTL相关系数为-0. 75,呈现显著的负相关关系;东海渔区的渔获量与MTL相关系数为-0. 34,统计学上依然显著。以上结果表明,在渤黄海和东海渔区,人为捕捞对渔业资源的结构与质量变化有显著的影响,过度捕捞会带来营养级降低。南海渔区的渔获量与MTL无相关关系,这表明人为捕捞未对渔业资源的结构与质量产生显著的影响。从FIB指数来看,3个渔区的渔获量与FIB指数均呈现显著的正相关关系,这表明渔获量的增加能够抵消MTL降低带来的影响,从而导致FIB指数上升。渔获量与MTL、FIB指数之间的相互影响,使得渔业生态系统在结构和功能上保持着相对良好的健康状态。
3. 2 中国近海捕捞渔船不同作业类型对MTL及FIB指数的影响
图3 展示了1990—2020 年中国近海不同作业类型的渔获量变化情况。由图3可以看出,在3个渔区,不同作业类型中拖网的渔获量均是最多,但渤黄海和东海渔区拖网的渔获量要远远大于南海渔区拖网的渔获量。在渤黄海渔区,流刺网和定置网的渔获量大致相当,围网和钓具的渔获量则相对较少;在东海渔区,定置网的渔获量紧随在拖网的渔获量之后,流刺网、围网和钓具的渔获量则相对较低;在南海渔区,流刺网的渔获量仅次于拖网的渔获量,并且流刺网的渔获量占比在3个渔区中最高。
各作业类型的渔获量主要是由渔船的数量、吨位、功率所决定。本研究以2002、2011、2020年为例,分析上述因素对不同作业类型渔获量的影响程度。由图4可以看出,在渤黄海渔区,渔船的数量主要以流刺网和拖网为主,其中流刺网占比大于拖网;在渔船的吨位和功率方面,拖网占比远超过流刺网。在东海渔区,渔船的数量以拖网、流刺网和定置网为主,三者所占比例相差不大;在渔船的吨位和功率方面,拖网占比远大于其他4种作业类型占比,约占55%。在南海渔区,流刺网在渔船的数量中占比最大,高达70%左右;在渔船的吨位和功率方面,主要以拖网和流刺网为主,其中拖网占比稍高于流刺网。
为分析各渔区不同作业类型对渔业资源结构与质量的影响,本研究分别计算了不同作业类型的渔获量与MTL、各营养等级渔获量占比、FIB指数的相关系数(表2—表4。由表2可知,在渤黄海渔区,拖网渔船渔获量与低营养级渔获量占比呈现显著的正相关关系,与高营养级渔获量占比呈现显著的负相关关系。这表明,随着拖网渔船作业类型的渔获量增加,渤黄海渔区的高营养级渔获量占比将减少,而低营养级渔获量占比将增加,这是渤黄海渔区的MTL呈现下降趋势的原因之一(图1(b))。由表3可知,在东海渔区,拖网渔船渔获量与中营养级渔获量占比呈现显著的正相关关系,与高营养级渔获量占比呈现显著的负相关关系。这表明,随着拖网渔船作业类型的渔获量增加,东海渔区的中营养级渔获量占比将增加,而高营养级渔获量占比将会减少,这是东海渔区的MTL呈现下降趋势的原因之一(图1(b))。由表4可知,在南海渔区,拖网的渔获量虽然最高,但与高营养级渔获量占比下降的相关关系不显著;流刺网的渔获量与高营养级渔获量占比呈现显著的正相关关系。这表明,随着流刺网作业类型的渔获量增加,南海渔区的中、高营养级渔获量占比将增加,这是南海渔区的MTL呈现上升趋势的原因之一(图1(b))。
就FIB指数而言,渤黄海与东海渔区的渔获量中占比最大的拖网渔船与FIB指数呈现正相关关系,但南海渔区两者的相关关系却不显著。在南海渔区,流刺网的渔获量与FIB指数呈现显著的正相关关系。这表明,3个渔区主要作业类型的渔获量增加能抵消MTL 下降的负面效应,以维持健康的渔业生态系统结构与功能。
3. 3 海温变化对MTL及FIB指数的影响
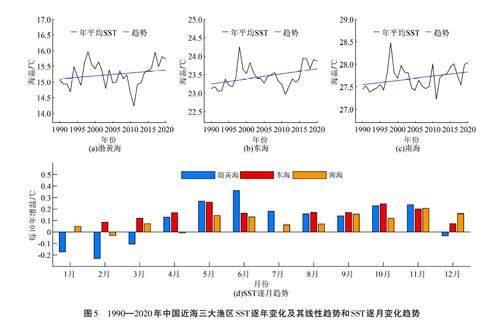
由图5可以看出,中国近海各渔区SST整体均呈上升趋势,其中东海增温最为显著,南海较弱。1998年,3个渔区的SST均达到最高值,是由于1997/1998年厄尔尼诺现象异常强烈,厄尔尼诺现象发生期间会导致中国近海的SST异常增暖,尤其是在厄尔尼诺现象发生的冬季和春季,对中国近海的SST升高影响最为显著[39]。2011—2012年,3个渔区的SST均达到最低值,其原因在于2011年拉尼娜现象强度较往年大[40],拉尼娜现象发生期间会导致中国近海的SST异常下降。2011年之后,中国近海各渔区SST上升趋势增加。南海渔区SST变化相对较弱是由于南海渔区海水温度和盐度的垂向结构变化非常复杂[41],同时受到南海季风[42]、黑潮入侵[43]、印度洋偶极子[44]等多因素复杂影响,因此南海渔区SST变化有一定的独特性[45]。
SST变化会影响到海洋生物的生长和发育[46-48],进而影响到营养级的变化[ 28]。在渤黄海渔区,SST与MTL整体上呈现负相关关系,这种关系主要由5—6月的SST与高营养级鱼类呈现显著的负相关关系所决定(图6(a))。就增温趋势而言,渤黄海渔区增温趋势最为显著的也是在5—6月,分别达到每10年0. 27 oC和0. 36 oC(图5(d)),从而抑制了同期高营养级鱼类的生长,这是渤黄海渔区的MTL呈现下降趋势的原因之一。在东海渔区,SST与MTL整体上呈现负相关关系,主要是由于秋季海温下降抑制了高营养级鱼类的生长,以9—11月最为显著;同时,SST与中营养级鱼类呈现正相关关系,一定程度上削弱了MTL的下降趋势(图6(b)),也表明东海渔区有中营养级鱼类的生长期在秋季。从图5(d)可以看出,东海渔区秋季增暖趋势也比较明显,基本上在每10年0. 17 oC~0. 24oC,表明东海渔区秋季增暖趋势抑制了高营养级鱼类的生长,从而对其MTL下降趋势有一定的贡献度。在南海渔区,SST变化与MTL整体上没有呈现显著的相关关系,但各营养级鱼类呈现不同的响应,尤其是在7—12月(图6(c)),SST与中营养级鱼类呈现显著的负相关关系,与高、低营养级鱼类呈正相关关系。这表明,SST异常增暖抑制了中营养级鱼类的生长,但促进了高、低营养级鱼类的生长,两种效应相互抵消,导致SST与MTL无显著的相关关系。由图5(d)可以看出,虽然南海渔区7—12月海温增暖也比较明显,基本上在每10年0. 06 oC~0. 2 oC范围内波动,秋季最强,但对南海渔区MTL 上升趋势没有贡献。
与MTL不同,SST对渤黄海和东海渔区的FIB指数没有显著影响,但对南海渔区的影响较为显著,主要是秋季SST与FIB指数呈现显著的正相关关系(图7)。因此,南海渔区FIB指数的增加与秋季海温增暖有关,即南海秋季海温增暖对于局地渔业生态系统持续健康有一定的贡献。
4 结论与建议
4. 1 主要结论
本研究聚焦中国近海渔业资源结构与质量变化,以渔获物平均营养级和渔业均衡指数为指标,较为系统地研究了人为捕捞和局地海温变化对渤黄海、东海及南海渔业资源结构与质量的影响,得出如下主要结论。
(1)渤黄海和东海渔区,分别以低营养级和高营养级鱼类占比最高,MTL呈下降趋势;南海渔区以中营养级鱼类占比最高,MTL无明显变化趋势。渤黄海和东海渔区的渔获量与MTL呈现显著的负相关关系,这表明人为过度捕捞已影响到渔业资源结构与质量。在南海渔区,渔获量与MTL之间并没有显著相关性,这表明人为捕捞活动未对南海渔区渔业资源的结构与质量产生显著影响。
(2)FIB指数在渤黄海和东海渔区的总体趋势均是先上升后下降,在南海则表现出微弱的上升趋势。3个渔区的渔获量与FIB指数均呈现显著的正相关关系,这表明渔获量的增长可以抵消MTL降低带来的影响,进而导致FIB指数上升,这种上升有助于维持渔业生态系统结构与功能的健康状态。
(3)在渤黄海和东海渔区,拖网渔船作业是导致MTL下降的一个重要原因,但在南海渔区,拖网渔船作业对MTL影响不大。就FIB指数而言,在渤黄海和东海渔区,拖网的渔获量占比最大,均与FIB指数呈现显著的正相关关系,但在南海渔区,两者的相关关系却不显著,南海渔区主要是流刺网的渔获量与FIB指数呈现显著的正相关关系,这表明3个渔区主要作业类型的渔获量抵消了MTL下降的负面效应,使得渔业生态系统结构与功能得以维持。
(4)在渤黄海渔区,MTL 的下降趋势是由于该区域5—6月份海温的快速增暖,这种现象对高营养级鱼类的生长产生了抑制作用。在东海渔区,秋季海温增暖对MTL下降趋势有一定的贡献度。但在南海渔区,MTL与SST变化没有呈现显著的相关关系。渤黄海和东海渔区的FIB指数受SST变化影响较小,但南海渔区FIB指数的增加与秋季海温增暖有关。
4. 2 建议
本研究着重分析了人为捕捞和气候变化对中国近海渔业资源结构与质量的影响,因此,各渔区在开展渔业活动时要综合考量这两个因素。①对于渤黄海渔区来说,建议合理控制捕捞量,根据海洋渔业资源最大产量的可持续性,确定适宜的捕捞渔船数量和功率,继续落实渔船“双控”政策;加强对拖网等非选择性渔具的管理和控制,积极发展流刺网、钓具等作业方式,加大政策扶持,促进拖改流工作取得良好成效。②对于东海渔区来说,要严格控制近海捕捞强度,压缩机动渔船,特别是小功率渔船的数量,采取合理的价格收购居民渔船并给予一定补偿,使部分专业渔民从海洋捕捞中退出;同时减少非选择性渔具(如拖网)的使用,优化渔具的选择,实施负责任的捕捞方式。③基于渤黄海和东海渔区海温增暖对MTL下降的显著作用,应采用遥感技术、GIS技术等,加强对海温的动态监测和评估,增进观测资料的共享;同时通过驯化手段提高一些经济鱼类的耐高温能力,以更好地适应外部环境的变化。④对于南海渔区来说,虽然渔区内整体高营养级渔获物的渔获量并没有出现下降现象,但是北部湾等近海大陆架内的渔获物由于受到过度捕捞的影响,已经出现了寿命长、高营养级的鱼类数量不断下降,而寿命短、低营养级的鱼类数量上升的现象,因此,在南海渔区,可适当拓展捕捞作业区,发展远洋捕捞,调整完善外海渔业结构,减轻近海捕捞压力。
本研究使用渔业统计数据对中国近海渔业资源的结构与质量进行探究,但由于渔区数据有限,故采用沿海不同省份的渔业统计数据表征各渔区数据,这可能会使分析结果的精确度降低。同时,技术进步、政策法规等因素也会对中国近海渔业资源的结构与质量产生影响,在以后的研究中需要综合多种因素进行评价。
参考文献
[1] 陈新军. 海洋渔业资源可持续利用评价[D]. 南京:南京农业大
学,2001.
[2] 鲁泉,苏雪,方舟,等. 基于渔获量平均营养级的东印度洋渔业
资源可持续利用评价[J]. 海洋学报,2021,43(8):118-127.
[3] 孙吉亭. 中国海洋渔业可持续发展研究[D]. 青岛:中国海洋大
学,2003.
[4] 朱玉贵,赵丽丽,刘燕飞. 海洋渔业资源可持续利用研究[J]. 中
国人口·资源与环境,2009,19(2):166-169.
[5] 李红艳,王颖,刘天红,等. 基于生态足迹的我国海洋水产资源
可持续利用研究[J]. 渔业研究,2021,43(5):509-516.
[6] 刘瑞玉. 关于我国海洋生物资源的可持续利用[J]. 科技导报,
2004,22(11):28-31.
[7] 卢秀容. 中国海洋渔业资源可持续利用和有效管理研究[D]. 武
汉:华中农业大学,2005.
[8] 蔡榕硕,刘克修,谭红建. 气候变化对中国海洋和海岸带的影
响、风险与适应对策[J]. 中国人口·资源与环境,2020,30(9):
1-8.
[9] 徐连章. 新制度经济学视角下的我国海洋渔业资源可持续利用
研究[D]. 青岛:中国海洋大学,2010.
[10] 孙康,周晓静,苏子晓,等. 中国海洋渔业资源可持续利用的动
态评价与空间分异[J]. 地理科学,2016,36(8):1172-1179.
[11] WU L X,CAI W J,ZHANG L P,et al. Enhanced warming over the
global subtropical western boundary currents[J]. Nature climate
change,2012,2(3):161-166.
[12] 吴佳颖,薛莹,刘笑笑,等. 黄、渤海渔业生物平均营养级的长
期变动[J]. 中国海洋大学学报(自然科学版),2017,47(11):
53-60.
[13] 李继龙,曹坤,丁放,等. 基于渔获物统计的中国近海鱼类营养
级结构变换及其与捕捞作业的关系[J]. 中国水产科学,2017,
24(1):109-119.
[14] 刘子飞. 我国近海捕捞渔业管理政策困境、逻辑与取向[J]. 生
态经济,2018,34(11):47-53.
[15] 刘子飞. 中国近海渔场荒漠化:评价、原因与治理[J]. 农业经
济问题,2019,40(6):105-116.
[16] 闫文彦,蒋杨徽,蔡立煌,等. 舟山市不同作业类型渔船捕捞产
量的灰色分析[J]. 浙江海洋大学学报(自然科学版),2018,37
(5):468-474.
[17] 宋伦,王年斌,董婧,等. 捕捞对长山群岛海域渔业生态系统的
影响[J]. 生态学杂志,2010,29(8):1578-1584.
[18] LIANG C,PAULY D. Fisheries impacts on China's coastal ecosystems:
unmasking a pervasive‘ fishing down effect[J]. PLoS one,
2017,12(3):e0173296.
[19] LINDEMAN R L. The trophic?dynamic aspect of ecology[J]. Bulletin
of mathematical biology,1991,53(1):167-191.
[20] PAULY D, CHRISTENSEN V, DALSGAARD J, et al. Fishing
down marine food webs[J]. Science,1998,279(5352):860-863.
[21] PAULY D, CHRISTENSEN V, WALTERS C. Ecopath, ecosim,
and ecospace as tools for evaluating ecosystem impact of fisheries
[J]. ICES journal of marine science, 2000, 57(3): 697-706.
[22] DUNLOP E S, GOTO D, JACKSON D A. Fishing down then up
the food web of an invaded Lake[J]. Proceedings of the National
Academy of Sciences of the United States of America, 2019, 116
(40): 19995-20001.
[23] 张忠,杨文波,陈新军,等. 基于渔获量平均营养级的东南大西
洋渔业资源状况分析[J]. 海洋渔业,2015,37(3):197-207.
[24] 丁琪,陈新军,李纲,等. 基于渔获统计的西北太平洋渔业资源
可持续利用评价[J]. 资源科学,2013,35(10):2032-2040.
[25] MILESSI A C, ARANCIBIA H. The mean trophic level of Uruguayan
landings during the period 1990-2001[J]. Fisheries research,
2005, 74(1-3): 223-231.
[26] 张瑛,李大海,耿涛. 气候变化背景下我国深蓝渔业的发展战
略研究[J]. 山东大学学报(哲学社会科学版),2018(6):
121-129.
[27] ASSIS J, TYBERGHEIN L, BOSCH S, et al. Bio?ORACLE
v2. 0: extending marine data layers for bioclimatic modelling[J].
Global ecology and biogeography, 2018, 27(3): 277-284.
[28] 许友卿,曹占旺,丁兆坤,等. 高温对鱼类的影响及其预防研究
[J]. 水产科学,2010,29(4):235-242.
[29] 黄佳兴,龚玉艳,徐姗楠,等. 南海中西部渔场主要渔业生物碳
氮稳定同位素特征[J]. 热带海洋学报,2019,38(1):76-84.
[30] 陈爽,陈新军. 气候变化对东北大西洋渔获物组成、多样性和
营养级的影响[J]. 海洋学报,2020,42(10):100-109.
[31] 刘尊雷,陈诚,袁兴伟,等. 基于调查数据的东海小黄鱼资源变
化模式及评价[J]. 中国水产科学,2018,25(3):632-641.
[32] 吴绍洪,赵东升. 中国气候变化影响、风险与适应研究新进展
[J]. 中国人口·资源与环境,2020,30(6):1-9.
[33] 肖启华,黄硕琳. 气候变化对东南太平洋智利竹筴鱼渔获量的
影响[J]. 中国水产科学,2021,28(8):1020-1029.
[34] 孙明,刘修泽,李轶平,等. 应用氮稳定同位素技术研究辽东湾
海域主要渔业生物的营养级[J]. 中国水产科学,2013,20(1):
190-198.
[35] 白怀宇,王玉堃,张廷廷,等. 基于碳、氮稳定同位素研究黄海
及东海北部主要鱼类的营养级和食性[J]. 渔业科学进展,
2021,42(2):10-17.
[36] 高春霞,戴小杰,田思泉,等. 基于稳定同位素技术的浙江南部
近海主要渔业生物营养级[J]. 中国水产科学,2020,27(4):
438-453.
[37] 邓景耀,姜卫民,杨纪明,等. 渤海主要生物种间关系及食物网
的研究[J]. 中国水产科学,1997,4(4):1-7.
[38] PAULY D,WATSON R. Background and interpretation of the
‘Marine Trophic Index as a measure of biodiversity[J]. Philosophical
transactions of the Royal Society B:biological sciences,
2005,360(1454):415-423.
[39] 鲍颖,兰健,王毅. 南海在1997/1998年El Ni?o事件后的异常变
化[J]. 地球科学进展,2008,23(10):1027-1036.
[40] 陈明诚,黄玉蓉. 典型拉尼娜事件与1988—1989年拉尼娜事件
生命史演变差异性成因分析[J]. 大气科学学报,2021,44(3):
428-440.
[41]WANG G H, XIE S P, QU T D, et al. Deep South China Sea circulation
[J]. Geophysical research letters, 2011, 38(5): L05601.
[42]WANG B, HUANG F, WU Z W, et al. Multi?scale climate variability
of the South China Sea monsoon: a review[J]. Dynamics of
atmospheres and oceans, 2009, 47(1-3): 15-37.
[43]ZHAO Y H, LIANG X S, YANG Y. The Kuroshio intrusion into
the South China Sea at Luzon Strait can be remotely influenced by
the downstream intrusion into the East China Sea[J]. Journal of
geophysical research?oceans, 2023, 128(8): e2023JC019868.
[44]ZHANG L, REN X Y, CAI W J, et al. Weakened western Indian
Ocean dominance on Antarctic sea ice variability in a changing climate
[J]. Nature communications, 2024, 15(1): 3261.
[45] 徐策,张力,余静,等. 气候变化对中国近海捕捞渔业的影响:
以太平洋年代际涛动为例[J]. 资源科学,2022,44(2):
386-400.
[46]MANTUA N J, HARE S R, ZHANG Y, et al. A Pacific interdecadal
climate oscillation with impacts on salmon production[J].
Bulletin of the American Meteorological Society, 1997, 78(6):
1069-1079.
[47]王万良,张忭忭,周建设,等. 不同水温对亚东鲑幼鱼生长及存
活的影响[J]. 水产科技情报,2019,46(1):24-27.
[48]POLOVINA J J, MITCHUM G T, GRAHAM N E, et al. Physical
and biological consequences of a climate event in the Central North
Pacific[J]. Fisheries oceanography, 2010, 3(1): 15-21.
(责任编辑:田红)
猜你喜欢
英语世界(2023年10期)2023-11-17 09:19:12
当代水产(2022年7期)2022-09-20 06:22:42
资源节约与环保(2022年12期)2022-02-02 02:39:16
中华环境(2021年9期)2021-10-14 07:51:16
海洋信息技术与应用(2020年2期)2020-07-27 01:42:04
今日农业(2019年14期)2019-09-18 01:21:36
当代水产(2019年3期)2019-05-14 05:43:18
生态文明新时代(2018年4期)2018-04-25 11:57:12
生态文明新时代(2018年1期)2018-03-21 05:16:48
海洋渔业(2016年6期)2016-04-16 03:00:27
