
铬污染下丛枝菌根真菌和生物质炭对紫花苜蓿生长和土壤微环境的改良效应
2024-07-02 15:01:05贾相岳吴文强
江苏农业科学 2024年9期
贾相岳 吴文强

摘要:为探究重金属铬(Cr)胁迫下丛枝菌根真菌(arbuscular mycorrhizal fungi,AMF)和生物质炭(biochar,BC)联合促进植物生长和改善土壤微环境的作用,试验采用盆栽法研究土壤Cr浓度为0、50 mg/kg时,不同生物质炭处理[对照(CK)、3%生物炭(BC)]并接种丛枝菌根真菌(AMF)对紫花苜蓿生物量、根系形态结构、土壤微生物群落、土壤酶活性以及球囊霉素相关蛋白(glomalina-related soil protein,GRSP)含量的影响。结果表明,重金属Cr 胁迫下,紫花苜蓿生物量显著下降,土壤微生物群落中真菌、细菌和放线菌数量减少,相关土壤酶活性降低,生物质炭处理能够帮助AMF侵染紫花苜蓿根系,显著增加AMF侵染率、丛枝着生率、侵入点位数和泡囊数。Cr浓度为50 mg/kg时,AMF和/或生物质炭处理均可以改善紫花苜蓿的根系形态结构(根系总长度、根系总体积、根尖数、根分叉数),增加总生物量和根冠比,与CK处理相比,土壤微生物中真菌数量增加26.47%~92.94%,细菌数量增加57.1%~164.9%,放线菌数量增加31.25%~88.94%;土壤微生物量碳含量增加32.24%~41.38%,微生物量氮含量增加47.37%~97.37%;AMF和/或生物质炭处理下土壤中过氧化氢酶、蔗糖酶、脲酶和碱性磷酸酶活性显著高于CK处理,分别提高4.59%~23.85%、5.00%~120.00%、
15.01%~39.97%和55.05%~152.29%,土壤易提球囊霉素相关土壤蛋白含量和总球囊霉素相关土壤蛋白含量分别是不接种对照的1.6~2.6倍和2.4~3.7倍。结论认为,AMF和/或生物质炭处理能够通过改善植物根系结构和土壤微生物群落结构,增加土壤相关酶活性和球囊霉素含量等提高植物对重金属Cr的抗性,促进植物生长,与单一处理相比,生物质炭和AMF具有协同增效作用,二者配合处理效果更好。
关键词:丛枝菌根真菌;生物质炭,铬胁迫;土壤;微生物;根系
中图分类号:S541+.106;X53文献标志码:A
文章编号:1002-1302(2024)09-0263-08
农业生产中化肥、农药的不合理使用,工业废水灌溉以及金属矿山污染等使得土壤中重金属沉降,对土壤理化性质以及植物生长造成不利影响[1-2]。重金属铬(Cr)广泛存在于自然界,其自然来源主要是岩石风化,后随工业化发展,城市污水灌溉、污泥及城市垃圾正在成为Cr污染的重要来源,因污染面积广、持续时间长、难降解等特性,成为土壤环境关注的焦点[3-5]。研究发现,土壤中Cr过多时,会抑制有机物质的硝化作用,并使Cr在植物体内蓄积,同时对土壤酶活性以及微生物环境平衡造成危害,显著抑制植物的生长发育状况[6-7]。徐成斌等发现,Cr对土壤中的脲酶和过氧化氢酶活性具有不同程度的抑制作用,有效铬与微生物量氮含量呈显著负相关[8]。徐雷等发现土壤pH值和碱解氮含量随Cr污染的增加而降低,有机碳和有效磷随Cr污染的增加而增加[9]。于皓等则认为高浓度的Cr污染会降低土壤细菌群落的多样性[10]。基于以上研究,修复重金属污染土壤,改善土壤质量是亟待解决的问题。
丛枝菌根真菌(arbuscular mycorrhiza fungi,AMF)是分布最为广泛的一类球菌门类真菌。AMF侵染植物根系后,可通过寄主植物来获取营养物质,保证真菌自身生长发育,同时帮助植物增强对外界营养物质的吸收和积累,从而达到互惠共生的目的[11-12]。同时AMF也具有增加土壤肥力、提高土壤酶活性等重要作用[13-14]。AMF除了吸收矿物质养分外,还能稳定土壤结构,使植物在有毒、有害重金属土壤等胁迫条件下生存[15]。AMF侵染后能够促进植物根系细胞壁的木质化,使寄主植物根尖表皮加厚、细胞层数增多,促进根系的生长、分枝,改变根系形态结构,从而影响重金属进入根系的进程[16]。研究发现,接种AMF的玉米根系pH值显著增加,而生物有效态铜浓度显著降低[17]。重金属污染下,接种AMF能够改变牧豆树(Prsopis juliflora)根际微生物的群落结构,增加细菌、AMF等的生物多样性[18]。
生物质炭(Biochar)是植物秸秆等在特定条件下,热解产生的一种多功能材料,生物质炭作为多功能的土壤改良剂,具有存碳量大、孔隙结构疏松、含氧活性基团数量多等特点[19],且能提供丰富的营养物质,防止土壤板结,为微生物活动提供有益的繁殖和生存条件[20]。刘会等发现添加生物质炭的苹果植株土壤中有机质、碱解氮、有效磷、速效钾和根际土细菌、放线菌、真菌数量均显著高于未添加处理,土壤容重显著降低[21]。另有研究发现,生物质炭和AMF之间存在协同作用,接种AMF并施用生物质炭能显著增加土壤中细菌和放线菌的数量,提高土壤酶活性,促进甜瓜生长[22]。重金属胁迫下,与单一生物质炭或者AMF处理相比,二者互作对土壤pH值、土壤养分及望江南根系的扩展、植株生长及干物质积累的改善效果最好[23]。
目前,对重金属Cr的研究较少,少数研究集中在植物对Cr的富集、转移和修复潜力上[24-25],针对微生物、植物、有机土壤改良物三者联合修复Cr污染土壤的研究则未见报道。本试验以紫花苜蓿为研究对象,探究Cr污染条件下接种AMF并配施生物质炭对紫花苜蓿生长、土壤酶活性以及微生物群落等的影响,从而认识AMF和紫花苜蓿对重金属胁迫的响应,以期为今后应用多重修复技术(菌根+植物+有机土壤改良物)修复重金属Cr污染提供参考依据。
1 材料与方法
1.1 试验材料
试验于2022年4月在晋中信息学院林下空地进行。供试植物材料为紫花苜蓿(Medicago sativa)中苜一号种子,为太原理工大学矿业工程学院自有。供试AMF菌种为地表球囊霉(Glomus versiforme),购自中国科学院生态环境研究中心,接种物为孢子、侵染根段、外生菌丝和培养基质的混合物,经检测,菌根侵染率70%,接种物含80个孢子/g。紫花苜蓿种子用过氧化氢溶液浸泡10 min后,用无菌蒸馏水冲洗表面消毒备用。栽培基质为壤土,采自晋中信息学院校园内,经高压蒸汽灭菌(120 ℃,2 h)后备用。花盆(规格:25 cm×18 cm×18 cm)经0.5%高锰酸钾溶液消毒1 h后使用。每盆装 3 kg 壤土,风干后过1 cm筛。土壤基本理化性质:pH值为6.83,有效磷含量为57.8 mg/kg,碱解氮含量为60.9 mg/kg,速效钾含量为44.9 mg/kg,有机质含量为2.5%。
1.2 试验方法
试验设计2个Cr浓度水平(0、50 mg/kg),4个处理组(CK,对照;AMF,接种地表球囊霉;BC,添加生物质炭;AMF+BC,接种地表球囊霉并添加生物质炭),共8个处理,随机排列,重复3次。其中不同浓度Cr处理以K2Cr2O7(Cr6+)的形式添加在土壤中,室内平衡7 d后用于试验。其中,CK处理作为对照;AMF处理是接种地表球囊霉,接种数量为 100 g/盆;BC处理是添加3%浓度生物质炭,由花生壳和牲畜粪便混合高温厌氧制成,购自山西纵合生物质炭有限公司;AMF+BC处理是先将生物质炭添加到土壤中混合均匀,之后再进行接种AMF与植物移栽。其他未接种处理则接种等量灭菌接种物。加入基质覆盖,灌水浇透,将5粒紫花苜蓿种子播于花盆中,覆膜保湿,放于人工气候室(光—暗周期为12 h—12 h,温度25 ℃,湿度70%)内培养;出苗后,留3株,每周浇水1次,5~10 d浇1次Hoagland营养液,保持水分在70%左右,培养2个月取样用于各项生理指标的测定,重复3次。
1.3 指标测定
菌根侵染情况的测定:采集紫花苜蓿的须根系,经FAA固定液固定,3%氢氧化钾90 ℃水浴15~30 min,用乳酸甘油染色法计算菌根侵染率和丛枝着生率,在电镜下观察侵入点位数和泡囊数[26],计算公式如下:
菌根侵染率=∑(0×根段数+10%×根段数+20%×根段数+…+100%×根段数)/总根段数。
总生物量和根冠比的测定:植株地上部和地下部分别收获后用水洗净后擦干,在105 ℃烘箱中杀青0.5 h,再于85 ℃下烘干至恒重,记作总生物量。根冠比为植物地下部与地上部干重的比值。
根系生长特性的测定:利用根系分析系统WinRHIZO(北京华辰阳光科技)进行紫花苜蓿根系总长度、根系总体积、根尖数和根分叉数的测定。
土壤酶活性的测定:采用高锰酸钾滴定法测定土壤过氧化氢酶(S-CAT)活性,采用磷酸苯二钠比色法测定土壤碱性磷酸酶(S-AKP)活性,利用靛酚蓝比色法测定土壤脲酶(S-UE)活性,利用3,5-二硝基水杨酸比色法测定土壤蔗糖酶(S-SC)活性,微生物量碳、氮测定采用三氯甲烷熏蒸-K2SO4浸提法[27-28]。
土壤微生物群落中真菌、细菌和放线菌的测定:采用涂布平板法测定土壤中细菌、真菌和放线菌。先称取待测土壤样品10 g,放入装有90 mL无菌水中,摇床振荡20 min,静置30 s;再用1 mL 无菌吸管,吸取10-1稀释液1 mL,移入装有9 mL无菌水的试管中,混合均匀,即成10-2稀释液;以此类推,连续稀释,制成10-3、10-4、10-5、10-6等一系列稀释菌液,再用稀释平板计数。
球囊霉素相关土壤蛋白的测定:易提取球囊霉素相关土壤蛋白(easily extractable glomalin-related soil protein,EE-GRSP)、总球囊霉素相关土壤蛋白(total glomalin-related soil protein,T-GRSP)采用张静等描述的方法[29]测定。
1.4 数据处理
采用Microsoft Excel 2010软件对数据进行处理和绘图,所有统计分析均采用SPSS 16.0进行统计分析,所有数据均进行单因素方差分析(one-way ANOVA)、双因素方差分析(two-way ANOVA)以及差异显著性检验(LSD法,α=0.05)。图表中数据为平均值±标准差。
2 结果与分析
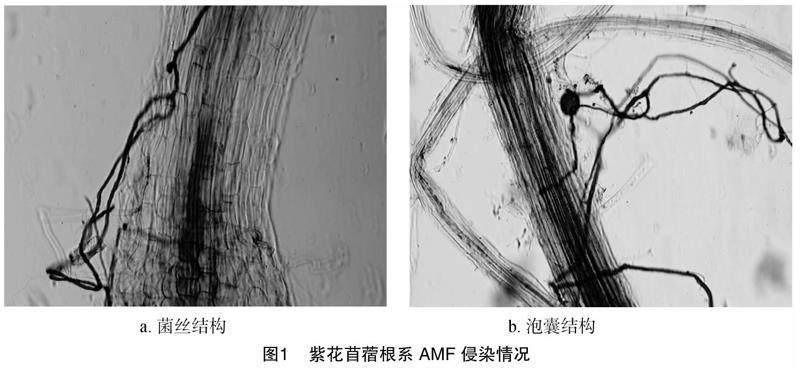
2.1 AMF和生物质炭对Cr胁迫下紫花苜蓿菌根侵染的情况
无论是否添加Cr,AMF或AMF+BC处理均可以与紫花苜蓿根系结成稳定的共生联系,形成丛枝菌根结构(图1)。未添加Cr处理下(Cr浓度为0),AMF侵染率达到最大,为56.5%,泡囊数为 15.8个/cm根长,丛枝着生率为35.2%,侵入点位数为10.2个/cm根长。Cr浓度为50 mg/kg时,AMF侵染率、泡囊数、丛枝着生率和侵入位点数显著降低,与AMF处理相比,AMF+BC处理下菌根侵染率、泡囊数、丛枝着生率和侵入位点数分别增加10.47%、9.92%、7.62%和13.33%(表1)。
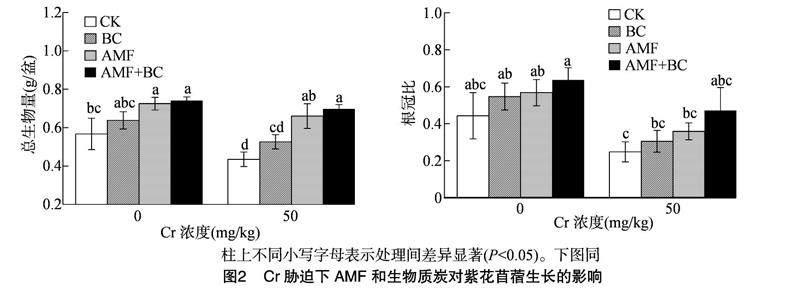
2.2 AMF和生物质炭对Cr胁迫下紫花苜蓿生物量的影响
由图2可知,Cr胁迫下,紫花苜蓿总生物量和根冠比表现为下降趋势。无论是否添加Cr,接种AMF和/或BC处理均可以增加紫花苜蓿总生物量和根冠比。未添加Cr处理下,与CK处理相比,BC处理下紫花苜蓿总生物量和根冠比分别增加12.52%和23.48%,AMF处理下分别增加27.87%和28.22%,AMF+BC处理下分别增加30.5%和43.8%。Cr浓度为50 mg/kg时,与CK处理相比,BC处理下紫花苜蓿总生物量和根冠比分别增加20.92%和22.98%,AMF处理下分别增加51.72%和44.76%,AMF+BC处理下分别增加60.23%和89.92%。
2.3 AMF和生物质炭对Cr胁迫下紫花苜蓿根系生长特性的影响
由图3可知,Cr胁迫下,紫花苜蓿地下部根尖数、根分叉数、根系总长度和根系总体积也表现为下降趋势。无论是否添加Cr,接种AMF和/或BC处理均可以增加紫花苜蓿地下部根尖数、根分叉数、根系总长度和根系总体积。未添加Cr处理下,与CK处理相比,BC处理下紫花苜蓿根尖数、根分叉数、根系总长度和根系总体积分别增加27.01%、60.00%、50.17%和10.87%,AMF处理下分别增加92.59%、78.57%、79.54%和72.83%,AMF+BC处理下分别增加98.98%、102.49%、96.61%和83.70%。Cr浓度为50 mg/kg时,与CK处理相比,BC处理下紫花苜蓿根尖数、根分叉数、根系总长度和根系总体积分别增加19.71%、30.67%、32.66%和38.71%,AMF处理下分别增加53.49%、63.05%、50.03%和100.00%,AMF+BC处理下分别增加73.59%、99.01%、117.44%和116.13%。
2.4 AMF和生物质炭对Cr胁迫下紫花苜蓿根系土壤微生物量碳、氮含量的影响由图4可知,Cr胁迫下,紫花苜蓿根系土壤微生物量碳、氮含量显著降低。无论是否添加Cr,接种AMF和/或BC处理均对紫花苜蓿根系土壤微生物量碳、氮含量有促进作用。未添加Cr处理下,与CK处理相比,BC处理下的紫花苜蓿根系土壤微生物量碳、氮含量分别增加20.50%和8.15%,AMF处理下分别增加29.77%和21.48%,AMF+BC处理下分别增加44.23%和38.52%。当Cr处理浓度为 50 mg/kg 时,与CK处理相比,BC处理下紫花苜蓿根系土壤微生物量碳、氮含量分别增加32.24%和47.37%,AMF处理下分别增加40.86%和71.05%,AMF+BC处理下分别增加41.38%和97.37%。
2.5 AMF和生物质炭对Cr胁迫下紫花苜蓿菌根侵染的情况
由表2可知,Cr胁迫下,紫花苜蓿根系土壤中真菌、细菌、放线菌数量呈现下降趋势。无论是否添加Cr,接种AMF处理对紫花苜蓿根系土壤中真菌、细菌、放线菌数量有增加作用,而BC处理下真菌数量下降,细菌和放线菌数量增加。未添加Cr处理下,与CK处理相比,BC处理下的真菌数量下降17.80%,细菌与放线菌数量增加45.71%和30.34%;AMF处理下真菌、细菌、放线菌数量分别增加37.70%、62.86%和13.86%;AMF+BC处理下分别增加96.86%、96.19%和59.18%。当Cr处理浓度为 50 mg/kg 时,与CK处理相比,BC处理下的真菌数量下降35.88%,细菌与放线菌数量增加57.14%和52.40%;AMF处理下真菌、细菌、放线菌数量分别增加26.47%、98.70%和31.25%;AMF+BC处理下分别增加92.94%、164.94%和88.94%。
2.6 AMF和生物质炭对Cr胁迫下紫花苜蓿根系酶活性的影响
由图5可知,Cr胁迫下,紫花苜蓿根系S-CAT活性受Cr刺激作用,表现为增加趋势,而S-SC、S-UE 和S-AKP活性表现为下降趋势。无论土壤是否添加Cr,接种AMF和/或BC处理均对紫花苜蓿根系土壤酶活性有促进作用。未添加Cr处理下,与CK处理相比,BC处理下的紫花苜蓿根系S-CAT、S-SC、S-UE和S-AKP活性分别增加41.43%、22.37%、7.03%和37.32%,AMF处理下分别增加55.71%、64.47%、30.87%和60.56%,AMF+BC处理下分别增加91.43%、114.47%、36.39%和135.21%。当Cr处理浓度为50 mg/kg时,与CK处理相比,BC处理下紫花苜蓿根系S-CAT、S-SC、S-UE和S-AKP活性分别增加4.59%、5.00%、15.01%和55.05%,AMF处理下分别增加12.84%、75.00%、21.21%和135.78%,AMF+BC处理下分别增加23.85%、120.00%、39.97%和152.29%。
2.7 AMF和生物质炭对Cr胁迫下紫花苜蓿根系土壤中GRSP含量的影响
由图6可知,Cr胁迫下,紫花苜蓿根系土壤中EE-GRSP、T-GRSP含量下降。无论是否添加Cr,接种AMF+BC处理对紫花苜蓿根系土壤中 EE-GRSP、T-GRSP含量有促进作用。未添加Cr处理下,与CK处理相比,BC处理下的EE-GRSP、T-GRSP含量无显著差异,AMF处理下分别增加76.00%和175.42%,AMF+BC处理下分别增加265.60%和303.91%。当Cr处理浓度为50 mg/kg时,与CK处理相比,BC处理下EE-GRSP、T-GRSP含量也无显著差异,AMF处理下分别增加65.42%和136.08%,AMF+BC处理下分别增加158.88%和265.19%。
3 讨论
环境问题特别是重金属污染已引起各类专家学者的广泛关注,重金属Cr因具有易吸附、难去除的特性,不仅危害土壤质量、植物生长,而且会随着食物链潜在威胁人畜健康[30]。土壤微生物在土壤有机质中占比较小,但土壤微生物在C、N和其他元素循环中起着关键作用。AMF是根际土壤微生物的一个重要组成部分,在陆地生态系统中广泛存在。与寄主植物形成的菌根共生体中根外菌丝为寄主植物生长发育提供相应的营养物质和水分条件,而寄主植物为真菌提供生长所需要的碳水化合物。已有多数研究报道,AMF能够通过增加土壤肥力、提高土壤酶活性等来增强对重金属的耐性[31]。而生物质炭因其独特的理化性质能够提高土壤碳氮矿化速率及改善土壤微生态环境等备受关注。本试验发现,重金属Cr胁迫下生物质炭与AMF存在协同增效作用,二者联合能够提高土壤相关酶活性,增加土壤微生物C、N含量以及真菌、细菌、放线菌数量,改善紫花苜蓿的根系形态并促进植物生长。本试验中,添加生物质炭能够促进AMF对紫花苜蓿的侵染,增加AMF的侵入点位数和泡囊数,显著促进了紫花苜蓿的生物量,改善植物根系情况。这可能是施用生物质炭调节了土壤养分含量,进而促进土壤AMF孢子繁殖和紫花苜蓿根系AMF定殖,AMF侵染紫花苜蓿根系表皮后,紫花苜蓿根系细胞增厚并分裂,显著改变寄主植物根系形态,增加根系总长度、根系总体积、根尖数和根分叉数。
土壤中一切生物变化的发生过程都离不开土壤中各类酶的参与,土壤酶活性是衡量土壤生物学活性和土壤生产力的重要标志,也是探讨重金属胁迫下土壤生态效应的有效指标[32]。金倩等研究发现,不同Cr处理对大豆根际S-UE活性有显著抑制效果,S-CAT活性呈现先升高后降低的趋势,S-SC 活性也呈现类似趋势[33] 。本试验下,Cr浓度为 50 mg/kg 时,紫花苜蓿根系S-CAT活性受Cr刺激作用,表现为增加趋势,而S-SC、S-UE和 S-AKP 活性表现为下降趋势,这与上述研究结果一致。接种AMF和/或生物质炭处理下,S-SC、S-UE、S-CAT以及S-AKP活性均表现增加趋势。冯慧琳等研究发现,生物炭施用后土壤蔗糖酶、脲酶、过氧化氢酶和中性磷酸酶活性增加,细菌多样性显著提升[34] 。王岩等发现生物炭和AMF接种能够改良辣椒连作土壤,生物炭与AMF配施处理菌根侵染效果最佳,侵染率高达59%,S-UE、S-SC 活性随生物炭添加量的增加呈增加趋势[35]。这可能是因为AMF通过侵染紫花苜蓿根系刺激根际土壤酶的产生和分泌,并促进其根系阳离子交换及活力提升,使得土壤酶活性显著增加。
土壤微生物量在一定程度上能代表参与调控土壤中能量和养分循环以及有机物质转化的对应微生物的数量[36]。本试验发现Cr胁迫下,土壤真菌、细菌和放线菌数量下降,接种AMF后土壤中微生物数量则显著增加,BC处理下真菌数量下降,而放线菌和细菌的数量增加。另有研究发现,重金属污染下稻田土壤的微生物生物量C、N含量比未明显污染的土壤显著降低约20%[37]。而生物炭提供易分解碳源并能为微生物生长发育提供场所,致使微生物量C、N含量增加。罗珍等通过两室分根装置种植玉米发现,接种AMF的菌根室中土壤微生物量C、N含量均有显著提高,并增加土壤呼吸作用,形成了明显有别于根际的微生物区系[38]。本试验也发现,重金属Cr胁迫下,土壤微生物量C、N含量显著下降,接种AMF和/或生物质炭处理下,土壤微生物量C、N含量表现为增加趋势。而其他研究中土壤微生物N的变化却是相反的,这和土壤N状况、植物N竞争都有关,具体还需进一步试验证明。AMF能分泌一种顽疏水、耐热、顽抗的糖蛋白-球囊霉素相关蛋白(Glomalina-related soil protein,GRSP),主要存在于AMF的菌丝以及孢子壁中,随菌丝脱落后沉积在土壤中[39]。GRSP有助于重金属的固定,产生强化学络合作用,降低重金属的生物可利用性和毒性[15]。本研究发现,Cr胁迫下土壤中EE-GRSP、T-GRSP含量下降,而接种AMF和/或生物质炭处理下,土壤中EE-GRSP、T-GRSP含量增加,与不接种对照和施用生物质炭处理差异显著,以接种AMF并添加生物质炭处理效果最好。这可能是因为生物质炭促进土壤微生物生长,特别是AMF的侵染与活性,从而可能增加微生物对矿物的分解及GRSP分泌。
4 结论
综上可知,重金属Cr胁迫下,AMF能够与紫花苜蓿形成菌根共生体系,添加生物质炭能够促进AMF对紫花苜蓿的侵染,提高菌根侵染率、丛枝着生率,并增加侵入点位数和泡囊数。接种AMF和/或生物质炭处理均能够促进植物生长,改善植物根系生长特性,增加根系总长度、根系总体积、根尖数和根分叉数。此外,AMF和/或生物质炭处理能够诱导产生较多的GRSP,增加土壤中微生物量碳、氮的积累,提高土壤中蔗糖酶、脲酶、过氧化氢酶和碱性磷酸酶活性,增加土壤微生物数量,显著提高对重金属Cr的抗性,相对于单一AMF或生物质炭处理,二者协同配施的效果更好。
参考文献:
[1]Ye L L,Chen Y S,Chen Y D,et al. Phytomanagement of a chromium-
contaminated soil by a high-value plant:phytostabilization of heavy metal contaminated site[J]. BioResources,2020,15(2):3545-3565.
[2]Zhang J K,Wang Z H,Ye Y. Heavy metal resistances and chromium removal of a novel Cr(Ⅵ)-reducing pseudomonad strain isolated from circulating cooling water of iron and steel plant[J]. Applied Biochemistry and Biotechnology,2016,180(7):1328-1344.
[3]武 坤,孔 潇,董 郁,等. 人工湿地植物对污水中重金属铬、镉、铅富集能力的整合分析[J]. 江苏农业学报,2022,38(6):1532-1540.
[4]姚金玲,王海燕,于云江,等. 城市污水处理厂污泥重金属污染状况及特征[J]. 环境科学研究,2010,23(6):696-702.
[5]牛 勇,余 辉,张 敏,等. 太湖流域典型河流沉积物重金属污染特征及生态风险评价[J]. 环境工程,2013,31(5):151-155,38.
[6]魏 远,郑施雯,朱建林,等. 重金属铬胁迫对土壤微生物数量及酶活性的影响[J]. 东北林业大学学报,2011,39(9):90-93.
[7]任宗玲,黄丽芸,李永涛,等. Cr3+、Ni2+单一及复合污染对水稻土酶活性的影响[J]. 华南农业大学学报,2010,31(4):16-21.
[8]徐成斌,孟雪莲,马溪平,等. 铬污染对土壤环境质量生物特征指标的影响研究[J]. 环境科学与管理,2012,37(8):1-3.
[9]于 皓,安益君,金德才,等. 铬污染对土壤细菌群落结构及其构建机制的影响[J]. 环境科学,2021,42(3):1197-1204.
[10]徐 雷,张小平,卞方圆,等. 铬污染对雷竹林土壤质量的影响[J]. 竹子学报,2022,41(4):45-50.
[11]Khandan Mirkohi A,Sheikh Asadi M,Taheri M R,et al. The effects of arbuscular mycorrhizal fungi and different phosphorus levels on some growth aspects of Lisianthus[J]. Journal of Science and Technology of Greenhouse Culture,2015,6(2):57-68.
[12]Nazari Deljou M J,Marouf A,Jaberian Hamedan H. Effect of inoculation with arbuscular mycorrhizal fungi (amf) on gerbera cut flower (Gerbera jamesonii) production in soilless cultivation[J]. Acta Horticulturae,2014(1034):417-422.
[13]毕银丽,孙江涛,王建文,等. AM真菌对采煤沉陷区黄花菜生长及根际土壤养分的影响[J]. 生态学报,2018,38(15):5315-5321.
[14]刘圆圆,张 丽,王 硕,等. 氮和土著AMF对黄瓜间作土壤酶活性及氮利用的影响[J]. 菌物学报,2019,38(11):1965-1975.
[15]游义红,李 博,李明锐,等. 重金属污染胁迫下接种AMF对玉米生长和镉铅含量的影响[J]. 山地农业生物学报,2019,38(3):38-45.
[16]张旭红,高艳玲,林爱军,等. 植物根系细胞壁在提高植物抵抗金属离子毒性中的作用[J]. 生态毒理学报,2008,3(1):9-14.
[17]申 鸿,刘 于,李晓林,等. 丛枝菌根真菌(Glomus caledonium)对铜污染土壤生物修复机理初探[J]. 植物营养与肥料学报,2005,11(2):199-204.
[18]Solís-Domínguez F A,Valentín-Vargas A,Chorover J,et al. Effect of arbuscular mycorrhizal fungi on plant biomass and the rhizosphere microbial community structure of mesquite grown in acidic lead/zinc mine tailings[J]. The Science of the Total Environment,2011,409(6):1009-1016.
[19]李 明,安忠义,王 浩,等. 生物质炭对土壤环境质量的影响概述[J]. 环境保护前沿,2018,8(4):293-301.
[20]刘杰云,邱虎森,王 聪,等. 生物质炭对双季稻田土壤反硝化功能微生物的影响[J]. 环境科学,2019,40(5):2394-2403.
[21]刘 会,朱占玲,彭 玲,等. 生物质炭改善果园土壤理化性状并促进苹果植株氮素吸收[J]. 植物营养与肥料学报,2018,24(2):454-460.
[22]王 策,郭绍霞,刘润进,等. 生物质炭和AM真菌联合施用对连作甜瓜生长和土壤理化性质的影响[J]. 中国瓜菜,2021,34(7):29-34.
[23]郭雄飞. 生物炭和AM真菌对重金属污染下土壤养分及望江南生长的影响[J]. 草业学报,2018,27(11):150-161.
[24]白利勇,季慧慧,孙文轩,等. 粉煤灰中重金属Pb/Cr/Cu在土壤-小白菜中的迁移与形态转化[J]. 土壤学报,2019,56(3):682-692.
[25]周 武,袁志辉,刘敏超. 重金属Cr(Ⅵ)和Ni对旱伞草富集能力及其生理生化指标的影响[J]. 环境工程学报,2015,9(1):171-176.
[26]刘润进,陈应龙. 菌根学[M]. 北京:科学出版社,2007.
[27]鲍士旦. 土壤农化分析. 北京:中国农业出版社,1998:56-83.
[28]关松荫. 土壤酶及其研究法[M]. 北京:农业出版社,1986.
[29]张 静,唐旭利,郑克举,等. 赤红壤地区森林土壤球囊霉素相关蛋白测定方法[J]. 生态学杂志,2014,33(1):249-258.
[30]姜 蒙,王 伟.重金属Cr6+耐受细菌的筛选研究[J]. 浙江农业学报,2016,28(2):324-329.
[31]李文彬,宁楚涵,徐 孟,等. 丛枝菌根真菌和高羊茅对压实土壤的改良效应[J]. 草业学报,2018,27(11):131-141.
[32]何学敏,吕光辉,秦 璐,等. 艾比湖地区典型乔灌群落水盐胁迫下土壤重金属对酶活性的影响[J]. 水土保持研究,2012,19(6):94-99.
[33]金 倩,刘 静,林立金,等. 锌铬复合胁迫对大豆根际土壤酶活性的影响[J]. 土壤,2015,47(6):1139-1143.
[34]冯慧琳,徐辰生,何欢辉,等. 生物炭对土壤酶活和细菌群落的影响及其作用机制[J]. 环境科学,2021,42(1):422-432.
[35]王 岩,周 鹏,白立伟,等. 生物炭和AM真菌配施对连作辣椒生长和土壤养分的影响[J]. 中国生态农业学报(中英文),2020,28(10):1600-1608.[HJ2mm]
[36]决 超. 微生物菌肥与土壤改良基质对连作马铃薯土壤性质及微生物群落的影响[J]. 江苏农业科学,2023,51(1):218-224.
[37]阎 姝,潘根兴,李恋卿. 重金属污染降低水稻土微生物商并改变PLFA群落结构:苏南某地污染稻田的案例研究[J]. 生态环境,2008,17(5):1828-1832.
[38]罗 珍,朱 敏,王晓锋,等. 分根装置中接种AM真菌对玉米秸秆降解及土壤微生物量碳、氮和酶活性的影响[J]. 中国生态农业学报,2013,21(2):149-156.
[39]王 建,周紫燕,凌婉婷. 球囊霉素相关土壤蛋白的分布及环境功能研究进展[J]. 应用生态学报,2016,27(2):634-642.
收稿日期:2023-07-17
基金项目:山东省自然科学基金(编号:ZR2018BB073)。
作者简介:贾相岳(1987—),男,内蒙古赤峰人,硕士,讲师,主要研究方向为土壤环境化学。E-mail:Zongshuiji681@yeah.net。
猜你喜欢
红领巾·探索(2021年2期)2021-08-26 15:02:02
四川劳动保障(2021年3期)2021-06-09 07:08:56
学生天地(2020年34期)2020-06-09 05:50:36
当代陕西(2019年11期)2019-06-24 03:40:26
现代园艺(2017年13期)2018-01-19 02:28:05
作文通讯·高中版(2017年12期)2017-02-06 05:56:42
中小企业管理与科技·下旬刊(2016年11期)2016-11-18 08:19:35
中国科技博览(2016年19期)2016-10-19 11:47:05
西南农业学报(2016年5期)2016-05-17 05:42:36
科技视界(2016年9期)2016-04-26 12:23:48
