
Pre-miR172及miR172调控油菜AP2基因表达的规律分析
2024-06-27 12:49刘芳郝小花陈中元何昊
广西植物 2024年5期
关键词:油菜
刘芳 郝小花 陈中元 何昊


DOI: 10.11931/guihaia.gxzw202305065
刘芳, 郝小花, 陈中元, 等, 2024.
Pre-miR172及miR172调控油菜AP2基因表达的规律分析 [J].
广西植物, 44(5): 936-950.
LIU F, HAO XH, CHEN ZY, et al., 2024.
Analysis of expression regulation of AP2 gene by pre-miR172 and miR172 in rape [J].
Guihaia, 44(5): 936-950.
摘 要: 为探究油菜miR172前体(pre-miR172)及成熟体(miR172)对AP2基因的调控功能,该研究通过生物信息学方法对miR172和AP2启动子进行调控元件预测,分析6条油菜AP2基因的进化关系及miR172与AP2的靶向关系;通过qRT-PCR方法检测AP2、miR172和pre-miR172在早熟和晚熟油菜不同组织的表达规律;比较分析miR172丰度和AP2表达量间的相关关系,以及比较分析 pre-miR172和miR172在表达水平上的相关关系;通过过表达pre-miR172,再次验证pre-miR172对成熟体miR172及AP2的作用。结果表明:(1)miR172和AP2启动子区均存在调控花发育的顺式元件。(2)6条AP2序列均经历了强烈的纯化选择,均具备miR172的结合位点,属miR172的靶基因。(3)miR172家族成员均可促进早熟油菜AP2表达,但miR172d作用不明显。在晚熟油菜中,miR172a和miR172c作用微弱,miR172b和miR172d二者共同发挥作用降低AP2的表达水平。(4)pre-miR172家族对于早熟油菜中miR172家族的表达水平均有促进作用;在晚熟油菜中pre-miR172a和pre-miR172b对其成熟序列的形成发挥正调控作用,pre-miR172c和pre-miR172d则对于其成熟序列的形成发挥负调控作用。过表达pre-miR172后,miR172和AP2表达规律与上述结果保持一致,证实pre-miR172对miR172及AP2的调控功能。该研究结果丰富了油菜AP2基因的功能调控路径,为基因的调控功能研究提供了新的思路。
关键词: 油菜, AP2, miR172, pre-miR172, 表达规律
中图分类号: Q943
文献标识码: A
文章编号: 1000-3142(2024)05-0936-15
收稿日期: 2023-09-12 接受日期: 2023-11-30
基金项目: 国家自然科学基金(32071930); 湖南省自然科学基金(2022JJ40288); 湖南文理学院博士科研启动项目(19BSQD22)。
第一作者: 刘芳(1988—),博士,讲师,研究方向为油菜分子育种,(E-mail)g5n2a5f@163.com。
*通信作者: 何昊,博士,讲师,研究方向为果树栽培,(E-mail)271285785@qq.com。
Analysis of expression regulation of AP2 gene
by pre-miR172 and miR172 in rape
LIU Fang1,2, HAO Xiaohua1, CHEN Zhongyuan1,3, HE Hao1,2*
( 1. College of Life and Environmental Science, Hunan University of Arts and Science, Changde 415000, Hunan, China; 2. Changde Research
Centre Agricultural Biological Macromolecule, Changde 415000, Hunan, China; 3. Hunan Provincial Key Laboratory
for Molecular Immunity Technology of Aquatic Animal Diseases, Changde 415000, Hunan, China )
Abstract: The appropriate flowering time is of great significance for crop yield. In the cropping pattern of “rice-rice-oil” in southern region, researching on the flowering period of Brassica napus can provide a theoretical basis for the breeding of early maturing rape varieties. The AP2 family transcription factors in rape are widely involved in the growth and development and play an important role during flower development. However, there are few studies exploring the regulation of AP2 at the microRNA level. In order to investigate the regulatory functions of the miR172 precursor (pre-miR72) and mature body (miR172) on AP2 gene in rape, the regulatory elements of miR172 and AP2 promoters were predicted based on bioinformatics, then the evolutionary relationship of six rape AP2 genes and the targeting relationship between miR172 and AP2 were analyzed, and the expression patterns of AP2, miR172 and pre-miR172 in different tissues of early and late maturing rape were detected by qRT-PCR. Finally, the correlation between miR172 abundance and AP2 expression level was studied, as well as the correlation between pre-miR172 and miR172. The results were as follows: (1) Both miR172 and AP2 promoter regions had cis-elements that regulated flower development. (2) The six AP2 sequences holded the strong purification selection, and they were the target genes of miR172 based on their binding sites for miR172. (3) miR172 family could promote the flowering of early maturing rape by increasing AP2 expression levels, except for miR172d. In late maturing rape, miR172a and miR172c performed weakly in flowering, while miR172b and miR172d worked together to reduce the expression level of AP2 to inhibit flowering. (4) The pre-miR172 family had a promoting effect on the expression level of miR172 family in early maturing rape; in late maturing rape, pre-miR172a and pre-miR172b exerted positive regulation on the formation of their mature bodies, while pre-miR172c and pre-miR172d exerted the opposite effects. After overexpression of pre-miR172, the expression patterns of miR172 and AP2 remained consistent with the above results, confirming the regulatory function of pre-miR172 on miR172 and AP2. The results of this study enrich the functional regulation pathway of rape AP2 gene, and provide new ideas for the study of gene regulatory function.
Key words: rape, AP2, miR172, pre-miR172, expression pattern
MicroRNA(miRNA)是一类内源性非编码小RNA,一般长度为18~36个核苷酸,参与基因转录水平的调控(张幸媛等,2021)。有研究表明,miRNA可以通过调控靶基因来调节植物生长发育(Huijser & Schmid, 2011)、开花时间(Spanudakis & Jackson,2014)和逆境胁迫(Liu & Axtell, 2015)。miRNA的形成先经过RNA聚合酶转录产生初级miRNA前体(primary miRNA precusor,pri-miRNA),再经DCL1切割后保留70~500个核苷酸序列形成茎环二级结构,即miRNA前体(miRNA precusor,pre-miRNA),最后pre-miRNA通过相似剪切后形成成熟miRNA(Phillips et al., 2007)。可见pre-miRNA对于成熟体miRNA的形成具有重要意义。
miR172广泛存在于植物中,最早在拟南芥中被克隆出来,其对植物的生长发育起基础作用(Park et al., 2002),在植物的生殖转换、花发育、响应胁迫机制等方面具有重要作用(王幼宁等,2016;赵晓晖等,2017)。拟南芥中miR172过表达抑制AP2蛋白水平,同时抑制成花因子TOE1和TOE2,导致拟南芥提前开花(Aukerman & Sakai, 2003)。在水稻中光敏色素可降低miR172d的表达,从而抑制AP2家族中OsIDS1基因和SNB基因的表达,诱导开花(Lee et al., 2014)。Li等(2019)在大岩桐中发现miR172过表达导致SsAP2-like基因表达量下降,促使其提前开花。此外,miR172通过靶向AP2类基因可以调控花器官形态的建成,如藏红花(Tsaftaris et al., 2012)、芥菜(Shivaraj & Singh, 2016)、玉米(Chuck et al., 2007)、水稻(Hu et al., 2009)、月季(Franois et al., 2018)、苎麻(马鑫,2017)等,可见miR172-AP2在花器官发育中发挥着重要作用(Ji et al., 2011)。
AP2亚家族是AP2/ERF转录因子家族成员之一,包含2个重复的AP2保守结构域,主要参与植物的生长发育过程(Trupiano et al., 2013;Zhao et al., 2015; Neogy et al., 2019)。最早,AP2结构域在拟南芥中发现,其与花发育相关(Jofuku et al., 1994),此后,陆续在水稻(Nakano et al., 2006)、油菜(Ghorbani et al., 2020)、苹果(Cheng et al., 2020)、葡萄(Licausi et al., 2010)等多个物种中发现,其参与花发育过程,但功能不尽相同。AP2的功能发挥会受到miRNA的调控,其调控开花时间的机制往往有miR172的参与来实现。
油菜miRNA的研究起步较晚,Shen等(2015)利用油菜基因组,鉴定到645个MIR基因,大部分miRNA具有高度保守性,调控油菜生长发育的各个方面,包括参与杂种优势调控、种子发育、植物病原互作、非生物胁迫和花发育等方面(陈丽等,2018)。Wang等(2019)分析早、晚花油菜中miR172或AP2类转录因子的表达模式和功能特性,发现它们对花器官发育具有调节作用,其中BnaAP2-1、BnaAP2-5和BnaTOE1-2可能起到抑制花开的作用。Shivaraj 和Singh(2016)分析白菜型油菜和芥菜中AP2的表达水平,揭示其参与芸薹属植物花的发育;对miR172和靶基因AP2突变体间杂交能进行分析,发现二者存在复杂的互作模式。突变miR172获得的转基因系均显示出加速开花的现象(Shivaraj et al., 2018)。目前,关于油菜中miRNA的研究大多集中在miRNA的挖掘与预测,而涉及miRNA与靶基因的调控关系,以及pre-miRNA对于miRNA调控的报道较少。
本文以油菜开花基因AP2的调控为研究区域,依托白菜和甘蓝中已发现的AP2基因为切入点,进行比对并获得油菜的AP2基因,采用生物信息学方法、qRT-PCR方法、相关性分析及过表达载体侵染油菜子叶的方法,通过对AP2进行蛋白理化性质、系统进化关系、选择压力、基序预测和顺式调控元件分析;对miR172、pre-miR172和AP2表达规律及miR172和AP2之间及pre-miR172与miR172之间的相关性进行分析;对过表达pre-miR172质粒侵染油菜子叶后miR172和AP2表达水平进行分析,拟探讨以下问题:(1)预测miR172和AP2与花发育的关系;(2)miR172对AP2的调控功能;(3)pre-miR172对miR172的调控功能。
1 材料与方法
1.1 试验材料
早熟油菜品种为湘油420,晚熟油菜品种为湘油15,均由湖南农业大学肖钢教授提供。将油菜种子种于大田,待其长至5~7叶期,选择长势一致且完整的植株,取根和叶,液氮速冻保存于-80 ℃冰箱中备用,待油菜生长至花期,取完整的花,液氮速冻保存于-80 ℃冰箱中备用。
1.2 油菜中AP2基因的鉴定
从TAIR(http://www.arabidopsis.org)和BRAD(http://brassicadb.cn)下载白菜、甘蓝的AP2基因和蛋白序列,经过NCBI比对,选择E-value≤10-10的序列作为目标油菜序列。为进一步明确候选油菜AP2家族成员,从miRBase(http://www.mirbase.org/)下载油菜miR172成熟序列(miR172)和前体序列(pre-miR172),将miR172成熟序列与已获得的油菜AP2序列通过TAPIR(http://bioinformatics.psb.ugent.be/webtools/tapir/)网站进行预测,保留具有靶向关系的AP2序列;同时,基于miRNA和靶基因间高度同源,利用DNAMAN比对miR172与油菜AP2基因,保留碱基错配数小于3的AP2序列作为本研究的候选基因。
1.3 miR172家族和AP2基因启动子分析
分别截取pre-miR172和AP2基因上游2 000 bp作为启动子区,通过New PLACE(https://www.dna.affrc.go.jp/PLACE/?action=newplace)网站进行顺式调控元件预测,初步预测miR172对于油菜开花的作用。
1.4 AP2系统发育分析
1.4.1 油菜AP2蛋白性质分析 利用在线软件Expasy(http://web.expasy.org/protparam/)预测分析候选油菜AP2蛋白分子量、等电点(PI),通过Cell-PLoc网站(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc/)进行亚细胞定位预测分析。
1.4.2 系统进化和选择压力分析 利用MEGA11软件的邻接法进行AP2、miR172和pre-miR172核苷酸序列的进化树分析,检验方法为步长检验(bootstrap),抽样次数1 000(1 000 replications)。利用DnaSp V6软件分析AP2基因的非同义突变率(non-synonymous,ka)和同义突变率(synonymous,ks),计算非同义突变率和同义突变率比值(ka/ks),ka/ks <1,认为有纯化选择作用,ka/ks >1,认为有正选择效应,ka/ks =1,认为存在中性选择。
1.4.3 油菜AP2基因基序分析 通过WebLogo(http://weblogo.berkeley.edu/logo.cgi)绘制油菜AP2蛋白序列结构域的序列标志图。通过MEME(http://meme suite.org/)鉴定油菜AP2蛋白内部的保守基序。
1.5 qRT-PCR检测AP2、miR172及pre-miR172表达规律
采用miRcute多糖多酚植物miRNA提取分离试剂盒[天根生化科技(北京)有限公司]提取油菜根、叶和花的miRNA。使用miRcute增强型miRNA cDNA第一链合成试剂盒[天根生化科技(北京)有限公司]合成cDNA,以此cDNA为模板,选择U6作为内参基因,采用miRcute增强型miRNA荧光定量检测试剂盒[天根生化科技(北京)有限公司]进行qRT-PCR检测。miR172家族成员的检测引物见表1,具体反应体系按照miRcute增强型miRNA荧光定量检测试剂盒说明书进行,每个样本3次重复,在CFX96 (BIO-RAD)定量PCR仪上进行扩增,运行程序为95 ℃ 15 min;94 ℃ 20 s,60 ℃ 34 s,40个循环;溶解曲线分析为65 ℃ 5 s, 95 ℃ 0.5 s。
采用RNA提取试剂盒(北京全式金生物技术股份有限公司)提取油菜根、叶和花的总RNA,并合成cDNA第一链。以cDNA为模板,以Actin为内参基因,采用TaKaRa公司[宝生物工程(大连)有限公司]TB Green Premix Ex Taq试剂盒进行qRT-PCR实验,检测AP2基因和pre-miR172表达情况,具体所用引物见表1。参照说明书按照25 μL体系进行配液,每个样本3次重复,在CFX96 (BIO-RAD)定量PCR仪上进行扩增,程序为95 ℃ 30 s;95 ℃ 5 s,55 ℃ 30 s,40个循环;溶解曲线分析为65 ℃ 5 s,95 ℃ 0.5 s。
1.6 油菜pre-miR172家族成员二级结构分析
通过RNAfold(http://rna.tbi.univie.ac.at//cgi-bin/RNAWebSuite/RNAfold.cgi)在线网站分析油菜pre-miR172家族成员,在默认参数条件下,预测其二级结构和自由能。
1.7 Pre-miR172过表达对成熟miR172及AP2基因的影响
1.7.1 构建pre-miR172过表达载体 为鉴定pre-miR172对miR172成熟体表达水平的影响,克隆pre-miR172前体序列(包含发夹区域上下游各100 bp区域序列),并在序列5′端引入酶切位点XhoI,3′端引入EcoRI酶切位点。经过XhoI和EcoRI双酶切分别处理质粒pGreen_GUS_competitor(addgene ID 55208)和含pre-miR172序列的T载体,将pre-miR172连接到pGreen_GUS_competitor上,构建过表达载体。将已构建的过表达载体(实验组)和空载体pGreen_GUS_competitor(对照组)分别转化至农杆菌GV3101(成智博等,2019)。
1.7.2 农杆菌瞬时侵染油菜子叶及体内验证实验
将实验组和对照组的农杆菌分别进行瞬时侵染油菜子叶, 具体方法参照谭小力等 (2012)。侵染后于25 ℃条件下避光培养过夜,然后转移至25 ℃,光周期16 h/8 h条件下培养3 d后取样,对miR172和AP2基因进行qRT-PCR检测,具体方法同上。
1.8 数据处理
1.8.1 相关性分析 通过Excel软件分别对油菜AP2和miR172家族以及miR172家族和pre-miR172家族进行相关性分析,在P<0.05的条件下,相关系数r绝对值越接近1,相关性越强。0≤|r|≤0.5,基本不相关或低度相关;0.5<|r|≤0.8,显著相关;0.8<|r|<1,高度相关;|r|=1,完全相关。
1.8.2 差异显著性分析 利用Microsoft Excel 2010对实验所得数据进行单因素方差分析,计算所得P值小于0.05为差异显著。
2 结果与分析
2.1 油菜中AP2基因的鉴定
油菜基因组来源于白菜和甘蓝的杂交,选择甘蓝和白菜中已鉴定的AP2基因,通过BLAST获得油菜中的同源序列,经过筛选去除重复序列,得到9条序列;经TAPIR网站预测miR172和AP2的互补配对位点(表2),选择错配碱基数小于3的序列保留,最终得到6条AP2候选序列。其中,根据白菜AP2序列比对得到2条序列XM_013887071和XM_048778068,其对应的蛋白序列为XP_013742525和XP_048600981;根据甘蓝AP2序列比对得到4条序列HQ637468、XM_048770445、XM_013887073和XM_048763426,其对应的蛋白序列为ADU04499、CDY29538、XP_013742527和XP_048619383。
2.2 油菜miR172和AP2启动子区顺式调控元件的预测分析
截取pre-miR172和AP2基因上游2 000 bp作为启动子区,经NewPLACE网站预测分析,在pre-miR172和AP2启动子区发现存在TATA-box、CAAT-box等核心启动子元件,光应答元件(G-box、AAAC-motif、Box4、GT-1等,用于响应光照长度和光周期刺激),激素应答元件(ABRE、 W-box、GARE、CGTCA、TGACG等,用于响应乙烯、脱落酸、赤霉素、生长素、乙酰水杨酸、茉莉酸的刺激),生长发育应答元件(TATTAG,参与细胞分裂),逆境胁迫响应元件(MYB2、UP1-motif等,参与响应水胁迫、损伤胁迫等机制),此外,启动子区还存在组织特异性元件(root-motif、POLLEN-ELEMENT,调控相应基因在根和花这些特定组织中表达)。这说明miR172和AP2可能参与调控开花过程。
2.3 油菜AP2系统发育分析
2.3.1 AP2蛋白性质分析 由表2可知,AP2蛋白由357~433个氨基酸组成;分子量为39.7~47.9 kDa;等电点为6.31~6.77,此外,亚细胞定位预测发现,6条油菜AP2均位于细胞核,说明这6条AP2序列均属核基因,在细胞核内发挥不同功能。
2.3.2 油菜AP2的选择压力分析 对油菜中6个AP2候选序列与白菜(Bra017809和Bra011741)和甘蓝(KC584094)中AP2进行ka、ks计算,通过比较ka/ks进行选择压力分析。由表3可知,旁系同源基因7对,直系同源基因6对,全部ka/ks<1,表明AP2家族在进化过程中经历了强烈的纯化选择。
2.3.3 系统进化树分析 为进一步确定油菜AP2、miR172和pre-miR172各自的进化关系,利用MEGA6软件对多种植物的AP2家族、miR172成熟序列和pre-miR172序列进行进化树分析,结果显示油菜AP2序列
XM_013887071、XM_048770445、XM_013887073与白菜BraAP2-2关系较近,XM_048763426与甘蓝BroAP2关系较近(图1:A)。Pre-miR172 进化树分析发现,bna-miR172a与bra-miR172a关系最近;bna-miR172b和bna-miR172c聚类在一组,和bra-miR172d、ath-miR172e关系较近;bna-miR172d和bra-miR172c关系较近(图1:C)。miR172 成熟序列的进化树分析,发现miR172b和miR172c聚类在一组(图1:B),表明二者可能功能相似;miR172a和miR172d分别聚类在其他组(图1:B),可能与miR172b、miR172c的功能存在差异。
2.3.4 AP2蛋白基序分析 由图2可知,所有油菜AP2蛋白候选序列均存在3个保守基序(图2: A),基序1(Motif 1)和基序2(Motif 2)高度保守,基序3(Motif 3)有3个氨基酸位置保守性较低(图2: B),说明不同油菜AP2基因功能的差异可能依赖于基序3来发挥。
2.4 miR172家族与AP2表达水平及二者相关性分析
由图3可知,AP2在根和叶中的表达水平呈现为早熟低于晚熟,但在花中则早熟明显高于晚熟(图3:A)。对于早熟油菜,根和叶中AP2表达量明显低于花的表达量;对于晚熟油菜则相反,呈现根和叶中AP2表达量高于花的。早熟油菜花中AP2表达量显著高于晚熟油菜花,初步推测AP2基因的综合作用结果是促进开花。
通过miRBase下载得到4条成熟油菜miR172家族成员,分别是bna-miR172a、bna-miR172b、bna-miR172c和 bna-miR172d。经过多重比对miR172家族成员和AP2基因,发现4个家族成员高度保守,仅存在1个碱基的差异,AP2基因和miR172高度同源互补,仅有1~3个碱基的差异(图3:F),初步推测这些AP2基因是miR172的靶基因。
通过qRT-PCR检测miR172家族的表达水平,结果如图3:B-E所示,miR172家族在早熟和晚熟油菜的根、叶和花中均有表达,而且花中的表达水平普遍高于根和叶,只有miR172d在晚熟油菜花中的表达量低于根和叶。 miR172a、miR172b和miR172c在早熟油菜不同组织中的表达规律和AP2一致,花中表达量高于根和叶,说明miR172a、miR172b和miR172c对于早熟油菜AP2基因有促进表达的作用。在晚熟油菜中,miR172a和miR172c在花中表达水平显著高于非花组织,但差距较小,说明miR172a和miR172c对于AP2基因虽表现为负调控,但作用较小;miR172b在花中表达量高于非花组织,与AP2表达规律相反,说明在晚熟油菜中miR172b对于AP2起到负调控的作用。在早熟油菜中,miR172d在不同组织之间表达量差异不显著,在晚熟油菜中miR172d表达规律和AP2一致,非花组织的表达量显著高于花的表达量,说明miR172d对于晚熟油菜AP2可能起正调控作用,而在早熟油菜中可能不发挥功能或作用微弱。
为进一步确定miR172家族各成员与AP2基因的关系,分析AP2和miR172家族各成员表达规律之间的相关系数(r值)和差异显著性(P值),结果如表4所示,在早熟油菜中miR172家族各成员与AP2均呈显著或高度正相关,在晚熟油菜中miR172d与AP2呈高度正相关, miR172b与AP2呈负相关,miR172a和miR172c与AP2低度相关,说明在早熟油菜中miR172家族促进AP2表达,但在晚熟油菜中miR172b抑制AP2表达,miR172d促进AP2表达。
综上所述,油菜AP2基因可能促进开花,而且其功能的发挥受到miR172的靶向调控。在早熟油菜中miR172家族促进AP2表达水平增加进而促进开花,但其中miR172d作用不明显。在晚熟油菜中,miR172a和miR172c对于AP2的调控作用较小,miR172b和miR172d二者共同发挥作用降低AP2的表达水平,从而抑制晚熟油菜的开花。
2.5 Pre-miR172的二级结构分析
从miRBase下载到4个欧洲油菜pre-miR172家族成员,4个白菜pre-miR172家族成员,2个甘蓝pre-miR172家族成员。利用 RNAfold(3.2)软件预测油菜、白菜和甘蓝的pre-miR172家族基因二级结构,结果如表5所示,pre-miR172的基本二级结构均为茎环结构,其自由能为40.3~67.7 kcal·mol-1,其中欧洲油菜pre-miR172c的二级结构自由能最高,欧洲油菜pre-miR172a的二级结构自由能最低。由图4可知,白菜和甘蓝pre-miR172a二级结构一致,但由于5′端缺少CGUU 4个碱基,3′端缺少CGACG 5个碱基,导致其在油菜中的二级结构缺少1个环区和部分茎部碱基配对,致使自由能下降3.1 kcal·mol-1;pre-miR172b在白菜和甘蓝中的二级结构一致,但欧洲油菜中则由于碱基差异,导致其茎部碱基配对数目增加,同时,虽然同为5个环区,但欧洲油菜中的环更小,因此二级结构自由能增加,稳定性更强。与白菜相比,欧洲油菜pre-miR172c整体碱基数更多,茎部配对数目更多,导致自由能更高,二级结构更稳定;欧洲油菜pre-miR172d相较于白菜多形成了1个环区,自由能降低,二级结构稳定性更弱一些。欧洲油菜中pre-miR172a的稳定性最低,然后依次是pre-miR172d、pre-miR172b,pre-miR172c稳定性最强。
2.6 miR172与pre-miR172功能的相关性分析
通过qRT-PCR检测pre-miR172家族各成员的表达水平,将pre-miR172家族和miR172家族表达量进行对比,发现在早熟油菜和晚熟油菜花中pre-miR172a表达量高于根和叶,miR172a也随之在花中呈现最高表达量(图5: A),说明pre-miR172a对于miR172a的形成有促进作用;但在晚熟油菜中,miR172a的变化幅度明显高于pre-miR172a的变化幅度(图5: A),说明除了pre-miR172a表达丰度对miR172a的形成发挥促进作用,可能还存在其他加工层面的调控参与。 pre-miR172b与对应的成熟体序列表达趋势一致,不同组织中前体序列表达量增加,成熟序列的表达量也随之增加(图5: B),说明pre-miR172b对miR172b的形成具有促进作用。pre-miR172c和pre-miR172d早熟油菜花中表达量高于非花组织,miR172c和miR172d与其前体保持同样的表达规律,说明pre-miR172c和pre-miR172d对于早熟油菜miR172c和miR172d的形成同样具有促进作用;但在晚熟油菜中,pre-miR172c和pre-miR172d的表达规律与其相应成熟体表达趋势相反(图5:C,D),说明在晚熟油菜中pre-miR172c和pre-miR172d对于其成熟体的形成具有负作用。
为进一步确定miR172的功能发挥与pre-miR172的关系,分析miR172和pre-miR172表达水平间的相关系数(表6),结果发现在早熟和晚熟品种中表现出一致的规律,miR172a和pre-miR172a间显著正相关。miR172d和pre-miR172d间显著相关,但在早熟品种中表现为正相关,在晚熟品种中表现为负相关, 说明成熟序列miR172a和miR172d的形成与相应前体表达丰度高低具有显著的相关关系。
综合上述结果, pre-miR172家族对于早熟油菜中miR172的表达水平具有促进作用,其中miR172a和miR172d的表达水平主要受pre-miR172a和pre-miR172d丰度的调控。在晚熟油菜中pre-miR172a和pre-miR172b对其成熟序列的形成发挥正向调控,pre-miR172c和pre-miR172d则对于其成熟序列的形成发挥负向调控。
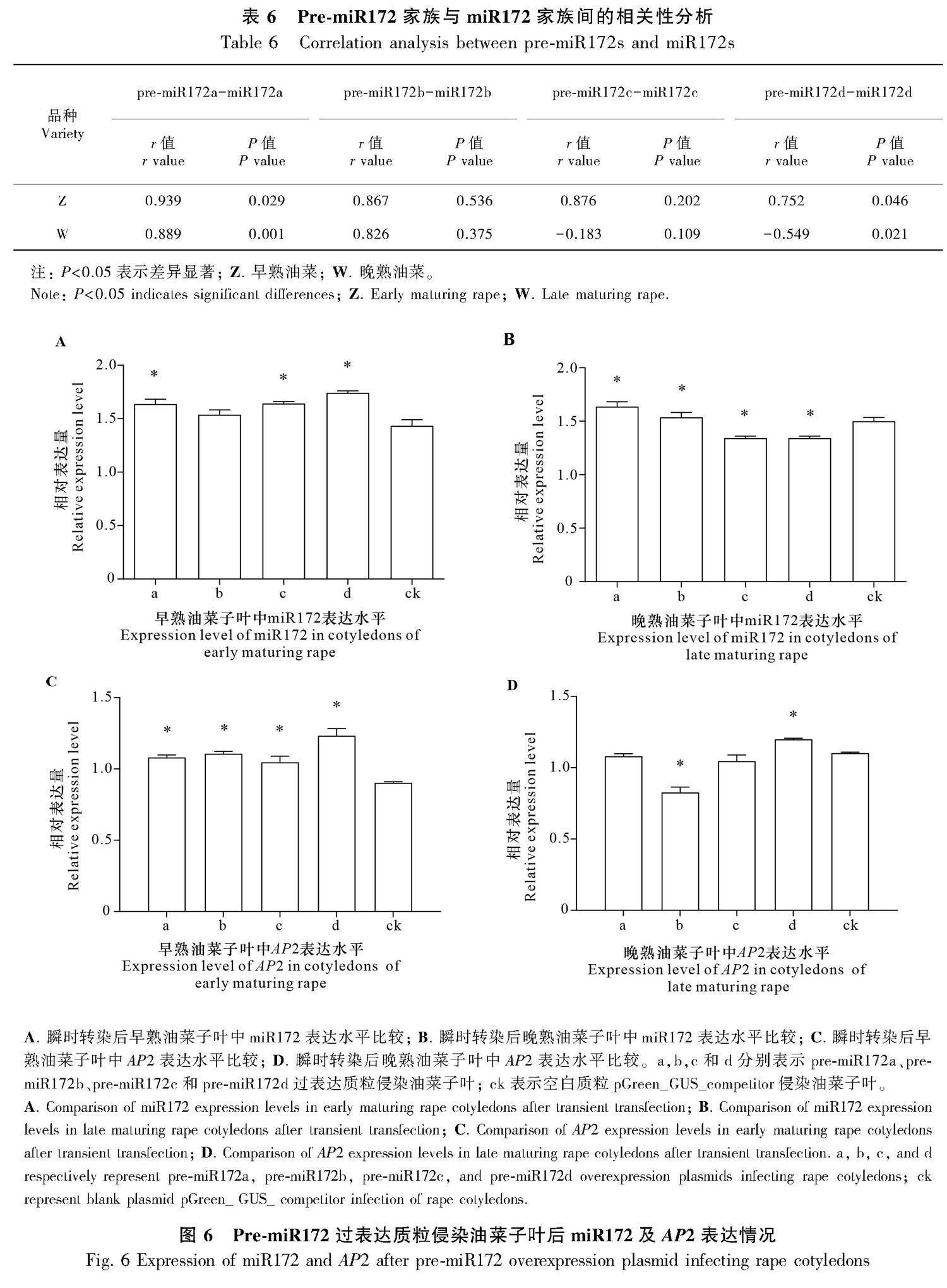
2.7 Pre-miR172对miR172及AP2表达水平影响
通过携带pre-miR172过表达质粒的农杆菌侵染油菜子叶进行体内验证实验,结果如图6所示,早熟油菜子叶中pre-miR172过表达均导致其对应的成熟体表达量高于对照组(ck),但其中miR172b与对照组相比差异不显著(图6: A),可见基本上,在早熟油菜中pre-miR172促进其成熟体的形成;晚熟油菜子叶中miR172a和miR172b表达量高于对照组,miR172c和miR172d则显著低于对照组(图6: B),表明前二者促进其成熟体形成,后二者抑制其成熟体形成。早熟油菜子叶中pre-miR172过表达后发现AP2基因表达水平显著高于对照组(图6: C),晚熟油菜子叶中过表达pre-miR172b时AP2表达量低于对照组,过表达pre-miR172d时AP2表达量高于对照组,过表达pre-miR172a和pre-miR172c时AP2表达水平略低于对照组,但差异不显著(图6: D)。过表达后AP2的表达规律与正常油菜中的表达规律一致,再次证明miR172靶向调控AP2基因。
3 讨论
3.1 AP2参与油菜花发育
在拟南芥中,AP2除了在花发育过程中表达,在非花组织茎和叶中也有表达,但在非花组织的表达水平较低,在花序分生组织和幼嫩花蕾中表达增强(赵奇等,2005)。本研究发现,AP2在油菜根、叶和花中均有表达,在早熟油菜的花中AP2表达量最高,与之规律一致;在晚熟油菜的根中AP2表达量较高,而花中却较低,初步推测获得的油菜AP2序列综合作用后可以正向调控开花的进行。Wang等(2019)认为其发现的euAP2家族成员中部分序列发挥负调控作用,其他成员在其所选样本中表达水平无明显差异。这与本研究结果不同,第一,筛选的候选AP2序列存在差异,可能导致检测结果不同;第二,本研究选择所有候选AP2序列的保守区设计的通用引物,并未分别检测每个家族成员的表达水平,其中发挥正向和负向调控作用的成员综合作用后呈现了正向调控的效果,这也可能是与前人结果不同的原因。
3.2 miR172对AP2的调控
基因表达受到不同水平的调控,包括染色体水平、转录水平、转录后水平和翻译水平等。转录因子在转录水平中发挥重要作用,转录因子主要通过与靶基因启动子区的DNA结合位点结合,激活或抑制下游基因的转录水平,进而起到调控基因表达的作用(Vaqueizas et al., 2009)。Saito等(2009)研究发现,转录因子可以与miRNA的启动子区结合,从而调节miRNA表达,miRNA通过与靶基因的3′UTR区特异性结合,进而在转录后水平抑制靶基因表达或者降解靶mRNA。转录因子和miRNA的有机结合增加了基因表达调控网络的复杂性(熊莉丽等,2014)。
AP2亚家族作为转录因子AP2/ERF基因家族成员之一,包含ANT和euAP2两种类型, 其中euAP2家族成员均含有miR172的结合位点。本研究获得的油菜AP2序列,与miR172比对后呈现1~3个碱基的错配,均含有miR172的结合位点,初步认为获得的AP2序列均属euAP2家族成员。拟南芥miR172已被证实通过切割euAP2和抑制蛋白翻译的方式调控AP2基因的表达(Aukerman & Sakai, 2003;Chen,2004)。通过分析油菜miR172和AP2表达规律间的相关性,本研究发现miR172家族丰度在早熟油菜中与AP2表达水平显著或高度正相关。但是,在晚熟油菜中仅miR172b和miR172d丰度与AP2表达量高度相关,而且前者为负相关,后者为正相关,miR172a和miR172c表达丰度与AP2表达量低度相关。这表明miR172在不同油菜品种中对于AP2调控功能的发挥存在差异。推测miR172家族成员对于AP2基因的调控不仅依赖于表达丰度,还存在加工层面的调控,致使在不同油菜品种中miR172家族成员的功能发挥有所不同。在早熟油菜中miR172家族成员对AP2表达水平起促进作用;在晚熟油菜品种中仅miR172d发挥促进作用,miR172a、miR172b和miR172d的作用可能经过加工后被削弱甚至不能发挥,导致其与AP2的关系不具备相关性,甚至呈负相关。
3.3 Pre-miR172调节miR172的表达水平
关于miRNA的研究大多集中在miRNA成熟体,有关pre-miRNA的报道较少。植物pre-miRNA对于成熟体miRNA的功能发挥有着重要意义。本研究通过检测pre-miR172和miR172表达量以及对二者进行相关性分析,发现miR172的表达量与pre-miR172的丰度有关,其中pre-miR172a和miR172a及pre-miR172d和miR172d高度或显著相关, pre-miR172b和miR172b及pre-miR172c和miR172c在丰度上不呈现线性相关,可能是因为miR172丰度不仅受到前体丰度调控,还受到加工层面的调控,所以单从表达量上不表现出线性相关。张俊红等(2012)在落叶松胚珠中同样发现这一规律,miR156、miR166、miR397、miR398、miR408丰度与其前体表达量均不呈现线性相关,认为其可能是受到加工调控;miR166在前4个发育阶段与前体表达量呈线性相关, 认为在前4个阶段miR166成熟体受到前体丰度调控,到后期受到加工调控。
在晚熟油菜中发现,pre-miR172a和miR172a的相关系数为0.889,表现为显著正相关,过表达pre-miR172a后miR172a的表达量也显著提高,说明pre-miR172a对于miR172a表达水平有正向调控作用;pre-miR172a表达量在根、叶、花中变化幅度不大,而miR172a变化幅度较大,这可能是由于pre-miR172a相对于成熟体更加不稳定,容易发生降解(Schmittgen et al., 2008),因此不能完全检测到。pre-miR172a自由能相较于其他家族成员最低,也表明其稳定性更低,容易降解。pre-miR172b和miR172b基本表现一致的表达规律,但二者不呈现线性关系,推测pre-miR172b对成熟体miR172b还有其他层面的加工调控。过表达pre-miR172b后,miR172b表达水平显著提高。 pre-miR172c和miR172c表达量在晚熟油菜中呈现相反的趋势,同时,过表达pre-miR172c后,miR172c表达水平降低且二者不呈现线性相关,可能是由于晚熟油菜中miR172c形成后,其前体pre-miR172发生了降解,因此检测到的前体表达量较低,但成熟体表达量较高。pre-miR172d与miR172d在晚熟油菜中呈负相关,表明pre-miR172d丰度的增加会抑制miR172d的形成,过表达pre-miR172d后,miR172d的表达水平显著降低,进一步证明在表达水平上pre-miR172d对miR172d起抑制作用。
耿立英等(2015)研究表明pre-miRNA种子区的单个碱基的差异就足以导致miRNA成熟体功能的不同,从而调控不同靶基因。油菜中miR172序列差异较小,但在不同材料不同组织中表达水平差异较大,其原因可能在于前体的调节,前体的调节方式不仅仅局限于丰度调节,还有加工层面的调控,有待进一步的研究。
4 结论
本研究分析miR172和AP2的启动子区,发现均存在调控花发育的顺式元件,可能参与花发育的调控。获得的6条AP2序列均具备miR172的结合位点,属miR172的靶基因,说明AP2可能受到miR172的调控。在早熟油菜中,miR172家族成员均可促进AP2表达,其中miR172d作用不显著。在晚熟油菜中,主要是miR172b和miR172d共同作用降低AP2的表达水平。miR172的形成受到其前体的调节,在早熟油菜中,pre-miR172家族促进miR172家族的表达;在晚熟油菜中pre-miR172a和pre-miR172b正向调控其成熟体的形成,pre-miR172c和pre-miR172d则对于其成熟体的形成发挥负调控作用。
参考文献:
AUKERMAN MJ, SAKAI H, 2003. Regulation of flowering time and floral organ identity by a microRNA and its APETALA2-like target genes [J]. Plant Cell, 15(11): 2730-2741.
CHEN L, LU HQ, LI RH, et al., 2018. Research progress and prospect of miRNA in Brassica napus [J]. Chin J Oil Crop Sci, 40(5): 664-673. [陈丽, 鲁海琴, 李日慧, 等, 2018. 油菜miRNA研究现状与展望 [J]. 中国油料作物学报,40(5): 664-673.]
CHEN X, 2004. A microRNA as a translational repressor of APETALA2 in Arabidopsis flower development [J]. Science, 303(5666): 2022-2025.
CHENG L, ZHAO T, WU YX, et al., 2020. Identification of AP2/ERF genes in apple (Malus× domestica) and demonstration that MdERF017 enhances iron deficiency tolerance [J]. Plant Cell Tissue Organ Cult, 143(2): 465-482.
CHENG ZB, WANG HT, ZHAO Q, et al., 2019. MiRNA172b-5p, miRNA172e-5p and miRNA472-3p responded to Cd stress by targeting MSH6 gene in Arabidopsis thaliana [J]. Chin J Ecol, 38(12): 3738-3746. [成智博, 王鹤潼, 赵强, 等, 2019. 拟南芥miRNA172b-5p,miRNA172e-5p和miRNA472-3p靶向MSH6基因参与Cd应激响应 [J]. 生态学杂志, 38(12): 3738-3746.]
CHUCK G, MEELEY R, IRISH E, et al., 2007. The maize tasselseed4 microRNA controls sex determination and meristem cell fate by targeting Tasselseed6/indeterminate spikelet1 [J]. Nat Genet, 39(12): 1517-1521.
FRANOIS L, VERDENAUD M, FU XP, et al., 2018. A miR172 target-deficient AP2-like gene correlates with the double flower phenotype in roses [J]. Sci Rep, 8(1): 12912.
GENG LY, ZHANG CS, ZHAO SY, et al., 2015. Polymorphism of pre-microRNA-1658 gene in chicken [J]. Sci Agric Sin, 48(19): 3919-3930. [耿立英, 张传生, 赵书雨, 等, 2015. 鸡mir-1658前体基因多态性分析 [J]. 中国农业科学, 48(19): 3919-3930.]
GHORBANI R, ZAKIPOUR Z, ALEMZADEH A, et al., 2020. Genome-wide analysis of AP2/ERF transcription factors family in Brassica napus [J]. Physiol Mol Biol Plants, 26(7): 1463-1476.
HU QH, UPADHYAYA NM, GUBLER F, et al., 2009. Over-expression of miR172 causes loss of spikelet determinacy and floral organ abnormalities in rice (Oryza sativa) [J]. BMC Plant Biol, 9(1): 149.
HUIJSER P, SCHMID M, 2011. The control of developmental phase transitions in plants [J]. Development, 138(19): 4117-4129.
JI LJ, LIU XG, YAN J, et al., 2011, ARGONAUTE10 and ARGONAUTE1 regulate the termination of floral stem cells through two microRNAs in Arabidopsis [J]. PLoS Genet, 7(3): e1001358.
JOFUKU KD, DEN BG, VAN MM, et al., 1994. Control of Arabidopsis flower and seed development by the homeotic gene APETALA2 [J]. Plant Cell, 6(9): 1211-1225.
LEE YS, LEE DY, CHO LH, et al., 2014. Rice miR172 induces flowering by suppressing OsIDS1 and SNB, two AP2 genes that negatively regulate expression of Ehd1 and florigens [J]. Rice, 7(1): 1-13.
LI XY, GUO F, MA SY, et al., 2019. Regulation of flowering time via miR172-mediated APETALA2-like expression in ornamental gloxinia (Sinningia speciosa) [J]. J Zhejiang Univ-Sci B (Biomed & Biotechnol), 20(4): 322-331.
LICAUSI F, GIORGI FM, ZENONI S, et al., 2010. Genomic and transcriptomic analysis of the AP2/ERF superfamily in Vitis vinifera [J]. BMC Genomics, 11(1): 719.
LIU Q, AXTELL MJ, 2015. Quantitating plant microRNA-mediated target repression using a dual-luciferase transient expression system [J]. Methods Mol Biol, 1284: 287-303.
MA X, 2017. Preliminary study on Ramie flower development related AP2/ERF gene [D]. Changsha: Hunan Agriculture University: 1-63. [马鑫, 2017. 苎麻花发育相关AP2/ERF类基因的初步研究 [D]. 长沙: 湖南农业大学: 1-63.]
NAKANO T, SUZUKI K, FUJIMURA T, et al., 2006. Genome-wide analysis of the ERF gene family in Arabidopsis and rice [J]. Plant Physiol, 140(2): 411-432.
NEOGY A, GARG T, KUMAR A, et al., 2019. Genome-wide transcript profiling reveals an auxin-responsive transcription factor, OsAP2/ERF-40, promoting rice adventitious root development [J]. Plant Cell Physiol, 60(10): 2343-2355.
PARK W, LI JJ, SONG RT, et al., 2002. CARPEL FACTORY, a Dicer homolog, and HEN1, a novel protein, act in microRNA metabolism in Arabidopsis thaliana [J]. Curr Biol, 12(17): 1484-1495.
PHILLIPS JR, DALMAY T, BARTELS D, 2007. The role of small RNAs in abiotic stress [J]. FEBS Lett, 581(19): 3592-3597.
SCHMITTGEN TD, LEE EJ, JIANG J, et al., 2008. Real-time PCR quantification of precursor and mature microRNA [J]. Methods, 44(1): 31-38.
SHEN EH, ZOU J, FALK HB, et al., 2015. Identification, evolution, and expression partitioning of miRNAs in allopolyploid Brassica napus [J]. J Exp Bot, (22): 7241-7253.
SHIVARAJ SM, JAIN A, SINGH A, 2018. Highly preserved roles of Brassica MIR172 in polyploid Brassica: ectopic expression of variants of Brassica MIR172 accelerates floral transition [J]. Mol Genet Genom, 293(5): 1121-1138.
SHIVARAJ SM, SINGH A, 2016. Sequence variation in Brassica AP2 and analysis of interaction of AP2-miR172 regulatory module [J]. Plant Cell Tiss Org, 125(2): 191-206.
SPANUDAKIS E, JACKSON S, 2014. The role of microRNAs in the control of flowering time [J]. J Exp Bot, 65(2): 365-380.
TAN XL, ZHUGE RJ, LI GY, et al., 2012. An agrobacterium-mediated transit transformation system in Brassica napus cotyledon [J], J Biol, 29(6): 93-96. [谭小力, 诸葛锐军, 李冠英, 等, 2012. 农杆菌介导的油菜子叶瞬时表达 [J]. 生物学杂志, 29(6): 93-96.]
TRUPIANO D, YORDANOV Y, REGAN S, et al., 2013. Identification, characterization of an AP2/ERF transcription factor that promotes adventitious, lateral root formation in Populus [J]. Planta, 238(2): 271-282.
TSAFTARIS AS, PASENTSIS K, MADESIS P, et al., 2012. Sequence characterization and expression analysis of three APETALA2-like genes from saffron crocus [J]. Plant Mol Biol Rep, 30(2): 443-452.
VAQUERIZAS JM, KUMMERFELD SK, TEICHMANN SA, et al., 2009. A census of human transcription factors: function, expression and evolution analysis [J]. Nat Rev Genet, 10(4): 252-263.
WANG TY, PING XK, CAO YR, et al., 2019. Genome-wide exploration and characterization of miR172/euAP2 genes in Brassica napus L. for likely role in flower organ development [J]. BMC Plant Biol, 19(1): 1-15.
WANG YN, SU C, ZOU YM, et al., 2016. Research progress of microRNA172 in plant development and stress responses [J]. Chin Bull Life Sci, 28(6): 645-654. [王幼宁, 苏超, 邹艳敏, 等, 2016. MicroRNA172 参与植物生长发育及逆境响应的研究进展 [J]. 生命科学, 28(6): 645-654.]
XIONG LL, ZHANG B, GUO ZY, 2014. MicroRNA and transcription factor regulatory network [J]. J Chongqing Univ (Nat Sci Ed), 37(6): 33-37. [熊莉丽, 张宝, 郭志云, 2014. MicroRNA与转录因子调控网络综述 [J]. 重庆大学学报(自然科学版), 37(6): 33-37.]
ZHANG JH, ZHANG SG, WU T, et al., 2012. Expression of five miRNA precursors and mature bodies during somatic embryo development in Larix gmelinii [J]. Chin Bull Bot, 47(5): 462-473. [张俊红, 张守攻, 吴涛, 等, 2012. 落叶松体胚发育中5个MiRNA前体与成熟体的表达 [J]. 植物学报, 47(5): 462-473.]
ZHANG XY, TIAN YH, QIN YZ, et al., 2021. The role of miR169 family members in the processes of growth, development and abiotic stress response in planta [J]. J Plant Genet Resour, 22(4): 900-909. [张幸媛, 田宇豪, 秦玉芝, 等, 2021. MiR169在植物生长发育与非生物胁迫响应中的作用 [J]. 植物遗传资源学报, 22(4): 900-909.]
ZHAO Q, WANG T, WEI XD, 2005. Function of AP2 gene during floral organs development in higher plant review [J]. Chin J Trop Agric, 25(3): 50-56. [赵奇, 王台, 魏小弟, 2005. AP2基因在高等植物花器官发育中的作用概述 [J]. 热带农业科学, 25(3): 50-56.]
ZHAO XH, KONG FJ, LIU BH, 2017. Research progress of miR172 and its targets in regulation of flowering and development [J]. Heilongjiang Agric Sci, (2): 126-130. [赵晓晖, 孔凡江, 刘宝辉, 2017. 植物miR172及其靶基因调控开花与发育的研究进展 [J]. 黑龙江农业科学, (2): 126-130.]
ZHAO Y, CHENG SF, SONG YL, et al., 2015. The interaction between rice ERF3 and WOX11 promotes crown root development by regulating gene expression involved in cytokinin signaling [J]. Plant Cell, 27(9): 2469-2483.
(责任编辑 邓斯丽)
猜你喜欢
今日农业(2021年21期)2021-11-26
今日农业(2021年14期)2021-10-14
今日农业(2021年7期)2021-07-28
西藏农业科技(2019年3期)2019-11-04
娃娃乐园·综合智能(2018年3期)2018-03-22
数学小灵通(1-2年级)(2017年10期)2017-11-08
湖南农业(2016年3期)2016-06-05
农家科技中旬版(2016年12期)2016-04-16
现代农业(2016年6期)2016-02-28
英语学习(2015年12期)2015-02-01
