
知觉学习中非显著性刺激视觉加工的学习机制
2024-06-11 07:38:24张琪王紫乐吴美君
心理学报 2024年6期
关键词:眼动
张琪 王紫乐 吴美君

摘 要 非显著性刺激的知觉学习研究发现成人大脑具有可塑性, 但是知觉学习如何影响不同的视觉加工阶段仍不清楚。通过将眼动指标划分为3个视觉加工阶段来探究知觉学习的机制:搜索潜伏期(早期), 是指从搜索屏呈现到第一次眼跳离开初始注视点位置的时间, 代表了在搜索屏中选择第一个搜索位置的时间; 注视点个数和平均注视时间(中期), 代表搜索过程中注视加工的位置个数和平均加工时间; 确定时间(后期), 代表判断当前刺激是否为目标并做出反应的时间。结果发现对训练刺激的搜索正确率提高, 反应时变快, 同时搜索潜伏期显著增加, 注视点个数和平均注视时间减少, 且行为和眼动指标的变化都没有迁移至未训练刺激。说明知觉学习会影响早期和中期视觉加工阶段, 可能通过增长搜索潜伏期, 同时减少眼跳的次数和降低注视时间来提高搜索表现。
关键词 知觉学习, 非显著性刺激, 学习机制, 视觉加工, 眼动
分类号 B842
1 引言
知觉学习是指知觉能力随着知觉训练或由于经验而改变的现象(Fahle, 2005)。早期的研究者认为, 大脑的可塑性是婴幼儿和儿童时期具有的能力, 成人大脑不具备可塑性(Hubel & Wiesel, 1965), 因此早期感觉区域的可塑性被认为局限于出生后生命早期的关键期。然而, 研究发现即使是初级感觉皮层在成年后也具有可塑性(Gilbert et al., 2001)。视觉领域知觉学习的研究采用多种任务, 都发现人类的视觉表现会随着练习而显著提高, 比如视觉光栅的检测或辨别(Kahnt et al., 2011)、运动方向辨别(Liu & Weinshall, 2000)、纹理辨别任务(Karni & Sagi, 1991)等。
知觉学习有两个基本特性:迁移性与特异性(马小丽 等, 2009)。迁移性是指经过训练后, 被试对训练刺激的视知觉能力有明显提高, 同时这种能力的提高也表现在与之相似的刺激上(马小丽 等, 2009; Fahle, 2005)。以往研究表明知觉训练的学习效果在某些情况下可以迁移到未训练的刺激, 比如朝向(J. Y. Zhang et al., 2010)、空间位置(Xiao et al., 2008)、運动方向(Liu & Weinshall, 2000)等。采用联合特征刺激(例如颜色和形状)的知觉学习研究发现, 知觉学习效应具有迁移性(Ding et al., 2023; Su et al., 2014), Su等人(2014)和Ding等人(2023)分别探讨了短期(40 ~ 50分钟)和长期(6 ~ 10天)的线段颜色?朝向联合特征搜索学习的迁移性, 结果都发现学习效应不能迁移到颜色和朝向同时变化的目标条件, 但可以部分迁移至只有一个特征(颜色或朝向)发生变化的目标条件。在Ding等人(2003)采用线段朝向作为刺激进行知觉学习, 研究结果发现虽然知觉学习效应在行为上没有表现出迁移性, 但在大脑活动上出现了迁移。然而, 知觉学习的效应并不总是迁移的, 其他一些研究发现知觉学习具有特异性(Fahle & Edelman, 1993; Saffell & Matthews, 2003)。特异性指的是知觉学习效应只局限于训练的刺激特征(Saffell & Matthews, 2003)。例如, 在游标视敏度辨别任务中, 对水平朝向游标进行训练后, 辨别能力提高, 但这种表现的提高并未迁移到垂直朝向(Fahle & Edelman, 1993)。单眼训练任务的结果也发现, 辨别能力的提高不会从训练眼迁移到未训练眼(Fahle et al., 1995)。
显著性视觉刺激在环境中相对醒目, 比如说明亮的交通灯、颜色鲜艳的广告牌等等, 因其与周围环境的差异更易引起人们的注意。而非显著性视觉刺激在环境中相对不引人注意, 与周围刺激的同质性较高, 比如在不同朝向的三角形中搜索某个特定朝向的三角形。目前已经有一些研究证据表明非显著性刺激的知觉学习与初级感觉皮层的变化有关(Fang et al., 2005; Maertens & Pollmann, 2005)。Maertens和Pollmann (2005)使用功能磁共振成像(fMRI)研究了知觉学习与初级视觉皮层的神经相关性, 实验结果发现知觉学习表现的提高伴随初级视觉皮层(V1)神经活动的增加, 同时这种知觉学习效果具有视野位置特异性。但也有证据表明非显著性刺激的知觉学习与更高水平的认知过程变化有关。例如, 研究者认为与自主注意力控制或决策相关的高级脑区(如额叶和顶叶皮层)在知觉学习中起重要作用(Kahnt et al., 2011; Law & Gold, 2008, 2009)。初级视觉皮层或高级大脑皮层的参与实际上反映了知觉学习特异性和迁移性的不同神经机制。对于初级视觉皮层来说, 神经元的感受野范围有限, 不同的朝向或位置由不同的神经元表征, 因此随着知觉学习的进行, 只有表征训练刺激的神经元发生了改变, 所以当知觉学习的变化涉及初级视觉皮层的改变时, 通常缺乏迁移性(Sagi & Tanne, 1994)。Bao等人(2010)发现光栅对比度检测任务具有位置特异性, 且在知觉学习后C1成分振幅增大, C1被认为源于V1, 表明知觉学习可能改变了初级视觉皮层的神经元感受野特性。而高级视觉皮层具有更大的感受野, 单个神经元可以对不同的简单线段朝向和位置敏感(Desimone & Ungerleider, 1989), 因此在涉及高级皮层的参与时, 知觉学习一般表现出迁移性。有研究采用运动方向辨别任务探究知觉学习的神经机制, 发现辨别能力提高且可以迁移到未训练刺激, 同时外侧顶内区(LIP)反应增强, 说明表征决策的高级皮层变化与知觉学习相关(Law & Gold, 2008)。
朝向属性的知觉学习研究一般包括简单线段朝向(Ahissar & Hochstein, 1997)和复杂形状朝向(例如三角形和T形) (Hu et al., 2018; Qu et al., 2017)两种。有研究者发现简单线段朝向的知觉学习具有迁移性(Ding et al., 2003)。Fang等人(2005)研究发现线段朝向知觉学习与初级视觉皮层相关, 被试对特定朝向线段训练后, V1区域活动水平增强。Qu等人(2017)对复杂形状朝向的知觉学习发现, 在训练后N2pc成分振幅增大, 表明复杂形状朝向在知觉学习后可以自动捕获注意。Sigman等人(2000)的研究發现复杂形状朝向的知觉学习具有位置及朝向特异性, 所以认为复杂形状朝向的知觉学习可能与初级视觉皮层有关。
Qu等人(2017)的研究中采用三角形朝向进行训练, 知觉学习后被试的搜索表现显著提高, 但未发现迁移性。前人研究发现任务难度会影响知觉学习的迁移(Ahissar & Hochstein, 1997), 在Qu等人(2017)的研究中训练后三角形的搜索正确率(p)仅为0.345, 因此本研究通过减小搜索集大小来降低任务难度, 探究三角形朝向在知觉学习后迁移性的缺失是由于任务难度还是由于三角形朝向本身具有知觉学习的特异性。此外, 实际的搜索过程包含不同的认知加工阶段及指标, 如搜索潜伏期、注视时间、注视点个数等。有研究者发现视觉搜索训练后, 搜索潜伏期延长, 注视点个数减少, 浏览时间变短, 搜索效率提高(Q. Zhang et al., 2022)。采用现实场景作为刺激的研究发现, 当目标出现在高概率区域时相比于低概率区域平均注视时间更少, 确认时间更短(Malcolm & Henderson, 2010)。因此, 只通过正确率和反应时并不能反映实际的视觉加工过程。
我们参考前人研究将眼动划分3个阶段(Malcolm & Henderson, 2009)来探讨非显著性刺激的知觉学习机制:第一个阶段为早期视觉加工阶段, 选择搜索潜伏期作为分析指标, 是指从搜索屏呈现到第一次眼跳离开初始注视点位置的时间, 代表了在搜索屏中选择第一个搜索位置的时间, 我们假设搜索潜伏期可能反映了在眼跳之前被试内隐注意的分配。有研究发现在眼跳之前, 注意就已经被分配到了目标出现的位置(Casteau & Smith, 2020; Kowler et al., 1995)。Q. Zhang等人(2022)的研究发现, 在知觉学习后, 搜索潜伏期延长, 并在第一次眼跳前观察到了目标引发的N2pc成分, 认为被试在眼跳前已经将内隐注意分配到目标位置。因此我们假设, 如果知觉学习后, 早期视觉加工阶段即搜索潜伏期显著变长, 说明被试可能通过增强内隐注意来促进搜索。第二个阶段为中期视觉加工阶段, 即浏览阶段, 被定义为从第一次眼跳到最后一次注视开始的时间, 这个阶段包括的指标有注视点个数和注视点的平均停留时间。研究者发现知觉学习后, 注视点个数(Q. Zhang et al., 2022)会发生变化, 表明知觉学习后被试只需要搜索更少的位置。因此知觉学习后, 如果注视点个数显著变少, 那么说明是通过搜索更少的位置来促进搜索表现; 如果注视时间显著变短, 那么说明加工每一个位置的视觉加工速度变快。第三个阶段为后期视觉加工阶段, 被定义为从最后一个注视点开始到按键为止, 即确定时间, 代表被试确定当前位置刺激是否为目标并做出判断的过程。有研究发现, 当目标出现在高概率区域时确认时间变短(Malcolm & Henderson, 2010), 表明被试可以更快识别目标。因此如果知觉学习后后期视觉加工阶段发生变化, 说明知觉学习会影响被试对当前位置是否有目标的判断过程。因此, 通过分析3个阶段的指标以及在前后测的差异, 可以探讨非显著性刺激的知觉学习效应如何影响不同的视觉加工阶段, 为进一步理解知觉学习的机制提供实验证据。
2 实验方法
2.1 被试
基于Hu等人(2018)的相似实验范式对非显著性刺激知觉学习的研究所得出的效应量大小(η2 p = 0.373), 采用MorePower 6.0 (Campbell & Thompson,
2012)计算至少需要16名被试(power = 0.8, α = 0.05)。共24名健康的成年人(M = 21.23岁, SD = 2.02岁; 19个女性)有偿参加了实验。所有被试视力或者矫正视力正常, 右利手, 并且对研究的目的不知情。实验前获得所有被试的书面知情同意书以及闽南师范大学心理与教育研究伦理委员会的批准(伦理批准号:2022-06-01)。
2.2 设备和刺激
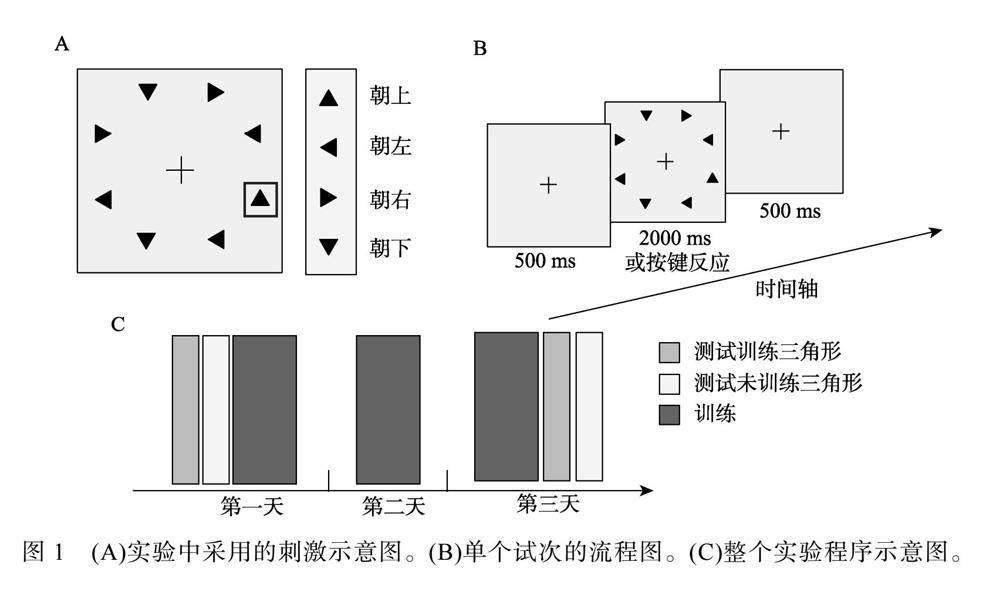
采用EyeLink 1000 (SR Research, Ontario, Canada)眼动仪记录被试的眼动轨迹, 并使用9点校准的方式进行校准和确认过程, 每组任务开始之前会进行漂移校正。采用MATLAB (MathWorks, Natick, MA, USA)环境下的Psychtoolbox 3.0工具包(Brainard, 1997)生成并呈现实验刺激。在实验过程中采用头托来固定被试的头部。显示器的分辨率为1920×1080, 屏幕长度为52.7 cm, 被试离屏幕的距离为63 cm。刺激阵列由8个黑色三角形组成(图1A), 用Spyder X (Lin et al., 2023)测得亮度值为0.12 cd/m2, 所有的刺激都呈现在亮度值为20.9 cd/m2的灰色背景上。8个三角形的边长都为1°, 且距离屏幕中央7°视角。实验中三角形有4种可能的朝向(朝上、朝下、朝左、朝右)。
2.3 实验程序
整个实验共3天, 分为前测、训练和后测三个过程(图1C)。第一天进行前测和训练, 第二天只进行训练, 第三天进行训练和后测。每个被试随机分配其中一个朝向作为训练三角形(例如朝上), 随机分配另外一个朝向作为未训练三角形(例如朝右), 其他朝向的三角形始终作为干扰物呈现。
在前测与后测会有两个搜索任务:分别为搜索
训练三角形任务(以朝上为例)和搜索未训练三角形任务(以朝右为例), 各160个试次, 同时记录被试的眼动轨迹。例如, 当被试的任务是搜索训练三角形时, 在目标不出现的试次(50%试次)中, 搜索显示的8个三角形由3种朝向(朝下、朝左、朝右)的三角形随机呈现。在目标出现的试次中(50%试次), 其中一个干扰物被训练三角形替换。当被试的任务是搜索未训练三角形时, 在目标不出现的试次(50%试次)中, 搜索显示的8个三角形由3种朝向(朝下、朝左、朝上)的三角形随机呈现。在目标出现的试次中(50%试次), 其中一个干扰物被未训练三角形替换。训练过程每天完成1600个试次(每组400个试次, 共4组), 不记录眼动轨迹。在训练过程, 只对训练刺激进行训练, 与前后测中搜索训练三角形的任务相同。
前后测及训练的实验流程图(如图1B), 每个试次开始会呈现中央注视点, 在记录眼动时(前后测), 被试需要持续盯住中央注视点(离中央注视点视角小于2°) 500 ms搜索屏才会呈现; 没有记录眼动时(训练), 中央注视点固定呈现500 ms后呈现搜索屏。搜索屏呈现之后被试可以自由眼动搜索, 并在保证正确的前提下尽快判断目标是否出现(50%的试次出现), 被试需要在搜索屏消失之前按键, 通过按键盘方向键的左键来报告目标出现, 或按右键来报告目标不出现, 搜索屏在按键后立即消失或在未按键的情况下呈现2000 ms后消失。
2.4 数据分析
2.4.1 行为分析
正确率和反应时(只计算正确试次)指标均采用2 (目标类型:训练三角形vs未训练三角形) × 2 (测试阶段:前测vs后测)的重复测量方差分析。我们采用以下公式報告搜索任务的正确率:
p = (p ? fp)/(1 ? fp)
其中p和fp分别指击中和虚报反应的比例(Hu et al., 2018; Qu et al., 2017)。为了使研究结果更为可靠, 我们报告了贝叶斯因子BF10 (Wagenmakers et al., 2018; Wasserstein & Lazar, 2016)。BF10小于1反映了更有利于零假设的证据(BF10为1 ~ 0.33反映了较弱的证据; 0.33 ~ 0.1为中等证据; < 0.1为支持零假设的非常强的证据), BF10大于1反映了更有利于替代假设(H1)的证据(BF10为1 ~ 3反映了较弱的证据; 3 ~ 10为中等证据; > 10为支持替代假设的非常强的证据)。
2.4.2 眼动分析
眼动指标的分析同样采用2 (目标类型:训练三角形vs未训练三角形) × 2 (测试阶段:前测vs后测)重复测量方差分析。将眼动轨迹划分为注视、眼跳和眨眼, 眼跳的定义为速度超过30°/s, 加速度超过8000°/s2, 运动距离超过0.1°; 眨眼的定义为超过3 ms的数据丢失; 其余的区间则被划分为注视, 且注视时间超过100 ms以上被视为一个注视点。
分析了第一个阶段的搜索潜伏期指标, 定义为从搜索屏呈现到第一次眼跳离开初始注视点位置的时间; 第二个阶段为浏览阶段, 定义为从第一次发生眼跳到最后一次注视开始的扫描时间, 浏览阶段中包括的指标有注视点个数和注视点的平均停留时间; 第三个阶段为确定时间, 定义为被试做出反应前最后一次注视的持续时间。当试次开始时, 被试伴随着眨眼或眼跳, 或搜索潜伏期小于80 ms的试次从分析中剔除, 同时行为反应错误的试次也在眼动分析中删除。在目标出现试次和目标不出现试次中, 前测与后测搜索训练三角形和未训练三角形的正确试次个数如图2所示。
3 研究结果
3.1 行为结果
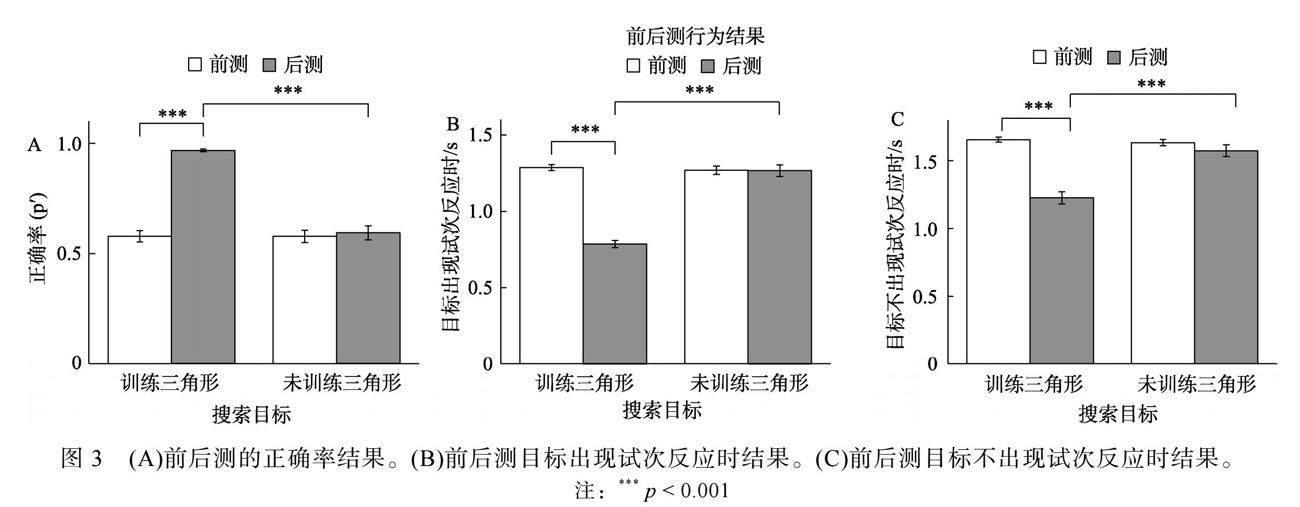
对于正确率(图3A), 2 (目标类型:训练三角形vs未训练三角形) × 2 (测试阶段:前测vs后测)重复测量方差分析结果显示, 目标类型(F(1, 23) = 83.25, p < 0.001, η2 p = 0.78)、测试阶段的主效应(F(1, 23) = 49.59, p < 0.001, η2 p = 0.68)及其交互作用(F(1, 23) = 101.68, p < 0.001, η2 p = 0.82)显著。简单效应分析发现, 训练三角形后测的正确率显著高于前测(Δ ?0.37 ± 0.14, t(23) = ?13.31, p < 0.001, Cohens d = 2.77, BF10 = 2.99, 95% CI = [?0.43, ?0.31]), 未训练三角形前后测差异不显著(Δ 0.00 ± 0.18, t(23) = 0.07, p = 0.942)。前测时, 两种目标类型正确率无显著差异(Δ 0.01 ± 0.12, t(23) = 0.22, p = 0.829), 后测时, 两种目标类型的正确率差异显著(Δ 0.38 ± 0.15, t(23) = 12.15, p < 0.001, Cohens d = 2.53, BF10 = 6.05, 95% CI = [0.32, 0.45])。
对于反应时, 当目标出现时(图3B), 目标类型(F(1, 23) = 178.68, p < 0.001, η2 p = 0.89)、测试阶段主效应(F(1, 23) = 130.64, p < 0.001, η2 p = 0.85)及其交互作用(F(1, 23) = 142.72, p < 0.001, η2 p = 0.86)显著。简单效应分析发现, 训练三角形后测显著快于前测(Δ 0.50 ± 0.10 s, t(23) = 24.26, p < 0.001, Cohens d =5.06, BF10 = 7.29, 95% CI = [0.45, 0.54]), 而未训练三角形前后测差异不显著(Δ ?0.01 ± 0.18 s, t(23) = ?0.40, p = 0.696)。前测时, 两种目标类型反应时无显著差异(Δ 0.0 1 ± 0.10 s, t(23) = 0.75, p = 0.462), 后测时, 两种目标类型反应时差异显著(Δ ?0.50 ± 0.16 s, t(23) = ?14.76, p < 0.001, Cohens d = 3.08, BF10 = 2.28, 95% CI = [?0.57, ?0.43])。当目标不出现时(图3C), 目标类型(F(1, 23) = 80.46,
p < 0.001, η2 p = 0.78)、测试阶段主效应(F(1, 23) = 65.62, p < 0.001, η2 p = 0.74)及其交互作用(F(1, 23) = 60.34, p < 0.001, η2 p = 0.72)显著。简单效应分析发现, 训练三角形后测的反应时显著快于前测(Δ 0.45 ± 0.20 s, t(23) = 11.06, p < 0.001, Cohens d = 2.28, BF10 = 8.74, 95% CI = [0.36, 0.53]), 而未训练三角形前后测反应时无显著差异(Δ 0.04 ± 0.17 s, t(23) = 1.13, p = 0.270)。前测时, 两种目标类型反应时无显著差异(Δ 0.04 ± 0.11 s, t(23) = 1.64, p = 0.114), 后测时, 两种目标类型反应时差异显著(Δ ?0.37 ± 0.21 s, t(23) = ?8.85, p < 0.001, Cohens d = 1.84, BF10 = 1.72, 95% CI = [?0.46, ?0.28])。这些结果表明, 训练三角形的正确率和反应时在知觉学习后表现出明显的知觉学习效果, 并且这种效果不会迁移到未训练三角形。
为了探究知觉学习效应随着训练量的变化, 我们将总训练量平均分成6个部分(训练量1-6)。正确率(图4A)的重复测量方差分析结果显示训练量主效应显著, F(1, 23) = 135.00, p < 0.001, η2 p = 0.85。事后比较的结果发现, 除训练量4和训练量5差异不显著(p = 1.000), 其余相邻训练量差异均显著(ps < 0.05)。目标出现试次反应时(图4B)重复测量方差
分析结果发现, 训练量主效应显著, F(1, 23) = 167.38, p < 0.001, η2 p = 0.88。进一步分析表明, 所有相邻训练量差异均显著(ps < 0.01)。目标不出现试次反应时(图4C)重复测量方差分析结果发现, 训练量主效应显著, F(1, 23) = 65.96, p < 0.001, η2 p = 0.74。进一步分析表明, 训练量2和训练量3差异不显著(p = 0.144), 训练量3和训练量4差异不显著(p = 0.113), 其余相邻训练量差异均显著(ps < 0.01)。这些结果说明经过3天的训练, 被试表现逐步提升。
3.2 眼动结果
3.2.1 搜索潜伏期
目标出现试次搜索潜伏期(图5A)的2 (目标类型:训练三角形vs未训练三角形) × 2(测试阶段:前测vs后测)重复测量方差分析结果显示, 目标类型(F(1, 23) = 2.00, p = 0.171)和测试阶段主效应 (F(1, 23) = 1.51, p = 0.232)不显著, 交互作用(F(1, 23) = 4.53, p = 0.044, η2p = 0.16)显著。简单效应分析结果表明, 训练三角形后测的搜索潜伏期显著长于前测(Δ ?32.43 ± 63.95 ms, t(23) = ?2.48, p = 0.021, Cohens d = 0.52, BF10 = 2.65, 95% CI = [?59.43, ?5.42]), 未训练三角形搜索潜伏期前后测差异不显著(Δ 0.31 ± 83.42 ms, t(23) = 0.02, p = 0.986)。前测时, 两种目标类型搜索潜伏期差异不显著(Δ ?3.99 ± 49.91 ms, t(23) = ?0.39, p = 0.699), 后测时, 两种目标类型搜索潜伏期差异显著(Δ 28.75 ± 63.51 ms, t(23) = 2.22, p = 0.037, Cohens d = 0.46, BF10 = 1.66, 95% CI = [1.93, 55.57])。
目标不出现试次的搜索潜伏期(图5B)结果同样发现, 目标类型主效应(F(1, 23) = 4.52, p = 0.044)显著, 测试阶段主效应(F(1, 23) = 1.06, p = 0.315)不显著, 交互作用显著, F(1, 23) = 8.88, p = 0.007, η2p = 0.28。简单效应分析结果表明, 训练三角形后测的搜索潜伏期显著长于前测(Δ ?45.16 ± 75.56 ms, t(23) = ?2.93, p = 0.008, Cohens d = 0.61, BF10 = 6.12, 95% CI = [?77.06, ?13.25]), 未训练三角形搜索潜伏期前后测无显著差异(Δ 13.51 ± 101.67 ms, t(23) = 0.65, p = 0.52)。前测时, 两种目标类型搜索潜伏期差异不显著(Δ ?9.67 ± 65.28 ms, t(23) = ?0.73, p = 0.475), 后测时差异显著(Δ 49.00 ± 67.07 ms, t(23) = 3.58, p = 0.002, Cohens d = 0.75, BF10 = 23.40, 95% CI = [20.68, 77.32])。这些结果说明, 无论目标是否出现, 训练三角形的搜索潜伏期在训练后显著性增加, 并且不会迁移到未训练三角形。
3.2.2 注视点个数
目标出现试次注视点个数(图6A)结果发现, 目标类型(F(1, 23) = 76.59, p < 0.001, η2 p = 0.77)、测试阶段主效应(F(1, 23) = 79.39, p < 0.001, η2 p = 0.78)及其交互作用(F(1, 23) = 51.74, p < 0.001, η2 p = 0.69)显著。简单效应分析结果表明, 训练三角形后测的注视点个数显著少于前测(Δ 2.04 ± 0.50, t(23) = 19.89, p < 0.001, Cohens d = 4.15, BF10 = 9.37, 95% CI = [1.83, 2.26]), 未训练三角形注视点个数前后测无显著差异(Δ ?0.02 ± 1.16, t(23) = ?0.10, p = 0.919)。前测时, 两种目标类型注视点个数差异不显著(Δ 0.09 ± 0.89, t(23) = 0.48, p = 0.638), 后测时差异显著(Δ ?1.98 ± 0.88, t(23) = ?11.10, p < 0.001, Cohens d = 2.31, BF10 = 9.32, 95% CI = [?2.35, ?1.61])。
目标不出现试次注视点个数(图6B)结果发现, 目标类型(F(1, 23) = 59.65, p < 0.001, η2 p = 0.72)、测试阶段主效应(F(1, 23) = 71.24, p < 0.001, η2 p = 0.76)及其交互作用(F(1, 23) = 48.14, p < 0.001, η2 p = 0.68)显著。简单效应分析结果表明, 训练三角形后测注视点个数显著少于前测(Δ 2.23 ± 0.85, t(23) = 12.84, p < 0.001, Cohens d = 2.68, BF10 = 1.46, 95% CI = [1.87, 2.59]), 未训练三角形注视点个数前后测无显著差异(Δ 0.18 ± 1.14, t(23) = 0.79, p = 0.437)。前测时, 训练三角形浏览过程中注视点个数显著多于未训练三角形(Δ 0.40 ± 0.73, t(23) = 2.65, p < 0.05, Cohens d = 0.55, BF10 = 3.59, 95% CI = [0.09, 0.70]), 后测时, 训练三角形注视点个数显著少于未训练三角形(Δ ?1.65 ± 0.91, t(23) = ?8.92, p < 0.001, Cohens d = 1.86, BF10 = 1.97, 95% CI = [?2.03, ?1.27])。这些结果说明, 无论目标是否出现, 训练后注视个数变少, 并且这种知觉学习效应不会迁移到未训练三角形上。
3.2.3 平均注视时间
目标出现试次平均注视时间(图7A)结果发现, 目标类型(F(1, 23) = 82.58, p < 0.001, η2 p = 0.78)、测试阶段主效应(F(1, 23) = 39.63, p < 0.001, η2 p = 0.63)及其交互作用(F(1, 23) = 44.05, p < 0.001, η2 p = 0.66)显著。简单效应分析结果表明, 训练三角形后测平均注视时间显著短于前测(Δ 63.40 ± 42.04 ms,t(23) = 7.39, p < 0.001, Cohens d = 1.54, log (BF10) = 11.47, 95% CI = [45.65, 81.16]), 未训练三角形平均注视时间前后测无显著差异(Δ ?4.82 ± 23.23 ms, t(23) = ?1.02, p = 0.321)。前测时, 两种目标类型平均注视时间差异不显著(Δ ?6.59 ± 19.90 ms, t(23) = ?1.62, p = 0.118), 后测时差异显著(Δ ?74.81 ± 42.84 ms, t(23) = ?8.56, p < 0.001, Cohens d = 1.87, log (BF10) = 13.80, 95% CI = [?92.90, ?56.73])。
目标不出现的试次平均注视时间(图7B)结果发现, 目标类型(F(1, 23) = 10.06, p < 0.01, η2 p = 0.30)主效应显著, 测试阶段主效应(F(1, 23) = 1.31, p = 0.263)及其交互作用(F(1, 23) = 0.29, p = 0.597)不显著。这些结果说明, 当目标出现时, 在训练后搜索训练目标的平均注视时间变短, 并且这种知觉学习效应不会迁移到未训练三角形。
3.2.4 确认时间
目标出现试次(图8A)的确认时间结果发现。目标类型(F(1, 23) = 0.99, p = 0.329)、测试阶段主效应(F(1, 23) = 1.25, p = 0.274)及其交互作用(F(1, 23) = 1.70, p =0.206)均不显著。目标不出现试次(图9B)确认时间的结果显示, 目标类型(F(1, 23) =2.68, p = 0.115)、测试阶段主效应(F(1, 23) = 0.37, p = 0.552)及其交互作用(F(1, 23) = 2.47, p = 0.129)也均不顯著。这些结果说明训练并没有显著影响视觉加工的后期阶段。
4 讨论
本研究采用与Qu等人(2017)类似的实验范式, 将整个视觉搜索的过程划分为3个视觉加工阶段, 更能直观地探究知觉学习对不同阶段产生的影响。本研究结果发现训练刺激在搜索潜伏期和浏览阶段出现了显著的知觉学习效应, 然而代表后期视觉
加工阶段的确认时间并没有受到知觉学习的影响, 这表明知觉学习效应发生在视觉加工的相对早期和中期阶段。为了进一步探讨知觉学习后搜索潜伏期增长与注视点个数减少之间的关系, 我们对搜索潜伏期与注视点个数进行了皮尔逊相关分析, 结果发现搜索潜伏期与注视点个数呈显著负相关(r23 = ?0.416, p < 0.05), 表明搜索潜伏期越长, 注视点个数越少。同时我们根据注视点个数将目标出现的试次分为两类, 浏览过程中注视点个数为0是指刺激出现后被试只进行了一次眼跳就做出了判断, 所以被试从第一个眼跳直到按键中间仅有一个注视点, 而此注视点归为确定时间, 并未计算到浏览过程中, 所以浏览过程中的注视点个数为0; 浏览过程中注视点个数大于0是指刺激出现后, 被试进行了多于1次眼跳后才做出判断, 即刺激出现后到按键中间有多于1个注视点。结果发现注视点个数为0 (30.32 ± 20.90%)的试次搜索潜伏期显著长于注视点个数大于0 (69.70 ± 20.90%)的试次(p < 0.001)。Qu等人(2017)的研究也发现非显著刺激的知觉学习引发了来自外纹状视皮层LOC的N2pc成分, Q. Zhang等人(2022)也在知觉学习后发现了特异于训练目标的显著N2pc成分且在眼跳发生之前, 说明知觉学习后空间注意会集中到训练目标刺激, 并在眼跳前进行注意分配, 以优化之后的搜索过程。因为只有在眼动产生之前将内隐注意力分配给刺激时, 才能观察到眼动前N2pc成分(Talcott & Gaspelin, 2021)。本文发现在知觉学习后注视点个数以及平均注视时间减少, 这可能在眼动之前就对内隐注意进行分配, 从而促进搜索。
有研究者同样记录了知觉学习前后的眼动轨迹, 发现知觉学习后搜索潜伏期增长, 注视点个数减少(Q. Zhang et al., 2022), 这与本研究结果一致。
Q. Zhang等人(2022)采用的颜色?朝向联合特征刺激属于维度间特征联结, 而本研究采用的三角形是维度内特征联结, 两者都属于非显著性刺激, 说明无论是几何形状的知觉学习还是联合特征的知觉学习, 在早期视觉加工阶段可能存在相似的学习机制。但是在中期视觉加工阶段, 尽管几何形状的知觉学习和联合特征的知觉学习都有更少的注视点个数, 但联合特征在知觉学习前后的平均注视时间差异不显著(Q. Zhang et al., 2022), 这与几何形状的知觉学习结果不一致, 说明尽管几何形状和联合特征的知觉学习在中期视觉加工阶段可能存在相似的学习机制, 但是两者之间的学习机制也并非完全相同。
双可塑性模型(Dual-Plasticity model) 提出了两种类型的知觉学习可塑性(Watanabe & Sasaki, 2015):基于特征的可塑性和基于任务的可塑性。基于特征的可塑性被视为对训练特征的神经反应的变化, 而基于任务的可塑性是指通过训练改善了与任务相关的处理过程, 这与视觉表征和认知区域之间的连接性的变化相关(Shibata et al., 2016)。这两种可塑性说明知觉学习存在着不同的机制。在本研究中, 如果知觉学习是基于特征的可塑性, 那么目标特征出现时的知觉学习效应, 应该与目标不出现时不同。而本研究结果发现搜索潜伏期、注视点个数以及确认时间无论在目标特征出现或者不出现时的知觉学习效应都是一致的, 说明非显著刺激的知觉学习可能是基于任务的可塑性。
不同任务难度对知觉学习的结果也有不同影响, 较简单任务的知觉学习一般涉及高级视皮层的变化, 因此会表现出迁移性, 而较困难和精细任务知觉学习一般涉及初级视觉皮层变化, 表现出特异性(Ahissar & Hochstein, 1997)。Qu等人(2017)在任务较难的情况下没有发现知觉学习的迁移性, 本研究减少搜索集大小来降低任务难度后, 训练三角形朝向的知觉学习效应仍未迁移到未训练三角形, 具有特异性。研究者认为知觉学习缺乏迁移性可能是由于学习效应的神经基础位于感觉加工的早期阶段(Karni & Sagi, 1991), 这种可塑性变化可能发生在初级视觉皮层。因为早期视觉皮层的神经元具有较小的感受野, 并对简单的刺激特征(如朝向)具有选择性, 因此更容易表现出特异性。尽管一些研究者将知觉学习的特异性归因于V1神经元的可塑性, 但知觉学习的神经机制可能更复杂。不仅涉及初级皮层的参与, 也涉及到感觉处理、决策、自上而下的任务相关性、行动选择过程等大脑区域网络的参与(Law & Gold, 2008, 2009)。这些感觉过程和认知过程都可能在任务过程中发生改变, 知觉学习可能涉及多個过程、层次和大脑区域。因此, 未来研究应把知觉学习视为过程性的改变, 可以从神经生理角度探究不同脑区在知觉学习不同阶段的参与情况, 而并非简单的将知觉学习特异性归为V1皮层的可塑性。
除了任务难度对知觉学习效果有影响之外, 训练时间对知觉学习也有不同影响。本研究在Qu等人(2017)的基础上, 降低任务难度的同时虽然缩短了训练时间, 但前后测的结果表明被试的搜索能力有显著的提高。有研究发现在短短一个小时的训练后, 知觉学习的成绩迅速提高(Fahle et al., 1995), 同样Ahissar与Hochstein (1996)在对pop-out检测的知觉学习中也发现, 行为表现在训练1400个试次后有了显著提升。Karni与Sagi (1993)提出知觉学习可能有快速学习阶段和慢速学习阶段, 快速学习阶段是在训练后短时间有了显著改善; 而慢速学习阶段是在最后一次训练后6~8小时之后才表现出明显的改善, 这一阶段的变化通常与睡眠有关。Ding等人(2023)的研究对颜色?朝向联合特征刺激进行搜索训练, 训练后发现了显著的知觉学习效应, 且这种知觉学习效应会迁移到与目标共享相同颜色或朝向特征的新目标上。在训练几个月后再次后测时发现, 颜色特征的搜索表现相比于第一次后测显著下降, 而朝向刺激的搜索表现与第一次后测没有显著差异, 研究者认为, 朝向特征搜索能力的提高相对困难, 且慢速学习一般可以长期维持, 所以认为朝向的学习主要是慢速学习的结果, 而颜色特征的学习可能是快速学习。
Ding等人(2023)的研究发现联合特征的学习效果不仅可以部分迁移到与被训练目标具有相同颜色或相同朝向的新目标上, 并且学习效应的总量近似等于单个特征迁移效应的总和。说明联合特征的知觉学习后并不是形成一个新的功能单元, 而是对单个特征进行学习的, 说明了对联合特征知觉学习是基于特征的注意增强机制(Ding et al., 2023; Su et al., 2014)。而根据单元化假设(unitization hypothesis), 维度内联合特征(如三角形)经过知觉学习后会形成一个与联合特征相对应的新的功能单元(Czerwinski et al., 1992), 并且知觉学习效应特异于训练目标, 不能迁移到任何新的目标上(Su et al., 2014)。未来研究需要更多的电生理证据从更深层次脑区加工的角度来探究两者之间的学习机制。
5 结论
本研究通过考察不同眼动指标所代表的视觉加工阶段探讨知觉学习促进搜索的学习机制。研究结果发现三角形朝向的知觉学习效应主要发生在早期以及中期的视觉加工阶段, 且具有特异性, 知觉学习可能是通过增加第一次眼跳前的搜索潜伏期来减少搜索的位置以及更快的加工每一个位置进而提高搜索表现。
参 考 文 献
Ahissar, M., & Hochstein, S. (1996). Learning pop-out detection: Specificities to stimulus characteristics. Vision Research, 36(21), 3487?3500. https://doi.org/10.1016/0042- 6989(96)00036-3
Ahissar, M., & Hochstein, S. (1997). Task difficulty and the specificity of perceptual learning. Nature, 387(6631), 401?406. https://doi.org/10.1038/387401a0
Bao, M., Yang, L., Rios, C., He, B., & Engel, S. A. (2010). Perceptual learning increases the strength of the earliest signals in visual cortex. Journal of Neuroscience, 30(45), 15080?15084. https://doi.org/10.1523/JNEUROSCI.5703-09. 2010
Brainard, D. H. (1997). The psychophysics toolbox. Spatial Vision, 10(4), 433?436. https://doi.org/10.1163/156856897X00357
Campbell, J. I. D., & Thompson, V. A. (2012). MorePower 6.0 for ANOVA with relational confidence intervals and Bayesian analysis. Behavior Research Methods, 44(4), 1255?1265. https://doi.org/10.3758/s13428-012-0186-0
Casteau, S., & Smith, D. T. (2020). Covert attention beyond the range of eye-movements: Evidence for a dissociation between exogenous and endogenous orienting. Cortex, 122, 170?186. https://doi.org/10.1016/j.cortex.2018.11.007
Czerwinski, M., Lightfoot, N., & Shiffrin, R. M. (1992). Automatization and training in visual search. The American Journal of Psychology, 105(2), 271?315. https://doi.org/ 10.2307/1423030
Desimone, R., & Ungerleider, L. G. (1989). Neural mechanisms of visual processing in monkeys. Handbook of Neuropsychology, 2(1983), 267?299.
Ding, Y., Li, T., & Qu, Z. (2023). Is a new feature learned behind a newly efficient color-orientation conjunction search? Psychonomic Bulletin and Review, 30(1), 250?260. https://doi.org/10.3758/s13423-022-02156-3
Ding, Y., Song, Y., Fan, S., Qu, Z., & Chen, L. (2003). Specificity and generalization of visual perceptual learning in humans: An event-related potential study. NeuroReport, 14(4), 587?590. https://doi.org/10.1097/00001756-200303240- 00012
Fahle, M. (2005). Perceptual learning: Specificity versus generalization. Current Opinion in Neurobiology, 15(2), 154?160. https://doi.org/10.1016/j.conb.2005.03.010
Fahle, M., & Edelman, S. (1993). Long-term learning in vernier acuity: Effects of stimulus orientation, range and of feedback. Vision Research, 33(3), 397?412. https://doi.org/ 10.1016/0042-6989(93)90094-D
Fahle, M., Edelman, S., & Poggio, T. (1995). Fast perceptual learning in visual hyperacuity. Science, 35(21), 3003?3013. https://doi.org/10.1126/science.1589770
Fang, F., Murray, S. O., Kersten, D., & He, S. (2005). Orientation-tuned fMRI adaptation in human visual cortex. Journal of Neurophysiology, 94(6), 4188?4195. https://doi.org/ 10.1152/jn.00378.2005
Gilbert, C. D., Sigman, M., & Crist, R. E. (2001). The neural basis of perceptual learning. Neuron, 31(5), 681?697. https://doi.org/10.1016/S0896-6273(01)00424-X
Hu, L., Ding, Y., & Qu, Z. (2018). Perceptual learning induces active suppression of physically nonsalient shapes. Psychophysiology, 56(9), 1?17. https://doi.org/10.1111/ psyp.13393
Hubel, D. H., & Wiesel, T. N. (1965). Binocular interaction in striate cortex of kittens reared with artificial squint. Journal of Neurophysiology, 28(6), 1041?1059. https://doi.org/10.1152/ jn.1965.28.6.1041
Kahnt, T., Grueschow, M., Speck, O., & Haynes, J. D. (2011). Perceptual learning and decision-making in human medial frontal cortex. Neuron, 70(3), 549?559. https://doi.org/ 10.1016/j.neuron.2011.02.054
Karni, A., & Sagi, D. (1991). Where practice makes perfect in texture discrimination: Evidence for primary visual cortex plasticity. Proceedings of the National Academy of Sciences of the United States of America, 88(11), 4966?4970. https://doi.org/10.1073/pnas.88.11.4966
Karni, A., & Sagi, D. (1993). The time course of learning a visual skill. Nature, 365(6443), 250?252. https://doi.org/ 10.1038/365250a0
Kowler, E., Anderson, E., Dosher, B., & Blaser, E. (1995). The role of attention in the programming of saccades. Vision Research, 35(13), 1897?1916. https://doi.org/10.1016/ 0042-6989(94)00279-u
Law, C. T., & Gold, J. I. (2008). Neural correlates of perceptual learning in a sensory-motor, but not a sensory, cortical area. Nature Neuroscience, 11(4), 505?513. https://doi.org/10.1038/nn2070
Law, C. T., & Gold, J. I. (2009). Reinforcement learning can account for associative and perceptual learning on a visual-decision task. Nature Neuroscience, 12(5), 655?663. https://doi.org/10.1038/nn.2304
Lin, Z., Ma, Q., & Zhang, Y. (2023). Psycalibrator: An open-source package for display gamma calibration and luminance and color measurement. Advances in Methods and Practices in Psychological Science, 6(2), 1?14. https://doi.org/10.1177/25152459221151151
Liu, Z., & Weinshall, D. (2000). Mechanisms of generalization in perceptual learning. Vision Research, 40(1), 97?109. https://doi.org/10.1016/S0042-6989(99)00140-6
Ma, X. L., Yang, B., Zhong, X., & Song, Y. (2009). The neural mechanism of perceptual learning. Advances in Psychological Science, 17(4), 653?658.
[马小丽, 杨彬, 钟翔, 宋艳. (2009). 知觉学习的神经机制. 心理科学进展, 17(4), 653?658.]
Maertens, M., & Pollmann, S. (2005). fMRI reveals a common neural substrate of illusory and real contours in V1 after perceptual learning. Journal of Cognitive Neuroscience, 17(10), 1553?1564. https://doi.org/10.1162/089892905774597209
Malcolm, G. L., & Henderson, J. M. (2009). The effects of target template specificity on visual search in real-world scenes: Evidence from eye movements. Journal of Vision, 9(11), 1?13. https://doi.org/10.1167/9.11.8
Malcolm, G. L., & Henderson, J. M. (2010). Combining top-down processes to guide eye movements during real-world scene search. Journal of Vision, 10(2), 1?11. https://doi.org/10.1167/10.2.4
Qu, Z., Hillyard, S. A., & Ding, Y. (2017). Perceptual learning induces persistent attentional capture by nonsalient shapes. Cerebral Cortex, 27(2), 1512?1523. https://doi.org/10.1093/ cercor/bhv342
Saffell, T., & Matthews, N. (2003). Task-specific perceptual learning on speed and direction discrimination. Vision Research, 43(12), 1365?1374. https://doi.org/10.1016/ S0042-6989(03)00137-8
Sagi, D., & Tanne, D. (1994). Perceptual learning: Learning to see. Current Opinion in Neurobiology, 4(2), 195?199. https://doi.org/10.1016/0959-4388(94)90072-8
Shibata, K., Sasaki, Y., Kawato, M., & Watanabe, T. (2016). Neuroimaging evidence for 2 types of plasticity in association with visual perceptual learning. Cerebral Cortex, 26(9), 3681?3689. https://doi.org/10.1093/cercor/bhw176
Sigman, M., & Gilbert, C. D. (2000). Learning to find a shape. Nature Neuroscience, 3(3), 264?269. https://doi.org/ 10.1038/72979
Su, Y., Lai, Y., Huang, W., Tan, W., Qu, Z., & Ding, Y. (2014). Short-term perceptual learning in visual conjunction search. Journal of Experimental Psychology: Human Perception and Performance, 40(4), 1415?1424. https://doi.org/ 10.1037/a0036337
Talcott, T. N., & Gaspelin, N. (2021). Eye movements are not mandatorily preceded by the N2pc component. Psychophysiology, 58(6), e13821. https://doi.org/10.1111/ psyp.13821
Wagenmakers, E. J., Marsman, M., Jamil, T., Ly, A., Verhagen, J., Love, J., ... Morey, R. D. (2018). Bayesian inference for psychology. Part I: Theoretical advantages and practical ramifications. Psychonomic Bulletin and Review, 25(1), 35?57. https://doi.org/10.3758/s13423-017-1343-3
Wasserstein, R. L., & Lazar, N. A. (2016). The ASAs statement on p-values: Context, process, and purpose. American Statistician, 70(2), 129?133. https://doi.org/ 10.1080/00031305.2016.1154108
Watanabe, T., & Sasaki, Y. (2015). Perceptual learning: Toward a comprehensive theory. Annual Review of Psychology, 66, 197?221. https://doi.org/10.1146/annurev- psych-010814-015214
Xiao, L. Q., Zhang, J. Y., Wang, R., Klein, S. A., Levi, D. M., & Yu, C. (2008). Complete transfer of perceptual learning across retinal locations enabled by double training. Current Biology, 18(24), 1922?1926. https://doi.org/10.1016/ j.cub.2008.10.030
Zhang, J. Y., Zhang, G. L., Xiao, L. Q., Klein, S. A., Levi, D. M., & Yu, C. (2010). Rule-based learning explains visual perceptual learning and its specificity and transfer. Journal of Neuroscience, 30(37), 12323?12328. https://doi.org/ 10.1523/JNEUROSCI.0704-10.2010
Zhang, Q., Huang, Z., Li, L., & Li, S. (2022). Visual search training bene?ts from the integrative e?ect of enhanced covert attention and optimized overt eye movements. Journal of Vision, 22(8), 1?52. https://doi.org/10.1167/jov.22.8.7
The mechanism of visual processing for nonsalient stimuli in perceptual learning
ZHANG Qi1,2,3, WANG Zile4, WU Meijun1
(1 School of Education and Psychology, Minnan Normal University, Zhangzhou 363000, China)
(2 Institute of Applied Psychology, Minnan Normal University, Zhangzhou 363000, China)
(3 Fujian Province Key Laboratory of Applied Cognition and Personality, Zhangzhou 363000, China)
(4 Institute of Psychology, Chinese Academy of Sciences, Beijing 100101, China)
Abstract
Previous studies have found that perceptual learning can improve the performance on visual search tasks. However, many cognitive processes are involved in visual search, and it is unclear at which visual processing stage perceptual learning facilitates search performance. The current study explored the mechanism of perceptual learning by dividing the eye movement metrics into three visual processing stages: search initiation time (the early visual processing stage), which represents the cognitive process of the time of processing the current location and selecting the next search location; scanning time (the middle visual processing stage), which includes the number and processing time of fixation positions; verification time (the late visual processing stage), which represents determining whether the current stimulus is the target and making a verification.
A 2 (target type: trained vs. untrained triangle) × 2 (test stage: pretest vs. posttest) within-subjects design was used to address the above issue. 24 healthy young adults (5 males; mean age: 21.23 ± 2.02 years) participated as paid volunteers in this study. We randomly selected one of the four orientations of the triangle (Up, Down, Left, or Right) as the trained triangle, which would receive three days of training. Moreover, to ensure that the visual search training was specific to the trained triangle, the trained and untrained triangles were tested by recording the behavior results and eye movement before and after training (untrained triangle was randomly selected from the distractors). Each trial started with a fixation cross (When eye movement was recorded, the search display would not appear until the participants fixated on the center cross for more than 500 ms; when eye movement was not recorded, the central fixation cross was presented for 500 ms and then the search screen was presented). Then a search display was presented until the key response or the elapse reached 2000 ms since its onset. The response was recorded only before the fixation cross disappeared. The task of participants was to determine whether or not the target was presented as quickly as possible. Participants pressed the left arrow key to report the presence of a target or the right arrow key to report its absence.
A two-way repeated-measures ANOVA was conducted with the factors of target type (trained vs. untrained triangle) and test stage (pretest vs. posttest). The behavior results found the reduced response time and increased accuracy when searching for trained stimuli after training. However, there was no significant difference in response time or accuracy between pretest and posttest for untrained stimuli. The results of eye movement tracking are as follows: (1) in the early visual processing stage, the search initiation time of the trained stimuli increased significantly after training, and there was no significant difference in the search initiation time between pretest and posttest for untrained stimuli. (2) In the middle visual processing stage, the number of fixations and the average fixation time of trained stimuli were significantly reduced after training, and there was no significant difference for untrained stimuli before and after training. (3) In the late visual processing stage, there was no significant difference in verification time between the pretest and posttest for both trained and untrained stimuli.
In conclusion, the accuracy and search initiation time of searching for trained stimuli was increased, while the number of fixations and the fixation time decreased. Moreover, the changes in behavior and eye movement indexes did not transfer to untrained stimuli. It is suggested that perceptual learning can affect the early and middle visual processing stages, and search performance may be improved by increasing the search latency, reducing the number of saccades, and reducing the fixation time.
Keywords perceptual learning, nonsalient stimuli, learning mechanism, visual processing, eye movement
猜你喜欢
包装工程(2023年18期)2023-09-27 09:17:36
心理学探新(2022年1期)2022-06-07 09:15:34
汽车实用技术(2022年7期)2022-04-20 11:44:42
载人航天(2021年5期)2021-11-20 06:04:32
大自然探索(2019年7期)2019-12-13 07:24:27
中国生物医学工程学报(2019年5期)2019-07-16 07:56:54
天津体育学院学报(2016年3期)2016-12-18 08:24:32
阅读(书香天地)(2016年5期)2016-05-30 01:01:58
工业设计(2016年12期)2016-04-16 02:51:51
外语学刊(2016年4期)2016-01-23 02:34:15
