
华北落叶松林林分密度对林下更新影响研究
2024-06-05 00:00:00陈欣韩有志
山西农业大学学报(自然科学版) 2024年2期


摘要:[目的]华北落叶松(Larix principis-rupprechtii)是寒温带针叶林的优势树种之一。探究华北落叶松的更新特征,可为深入探测华北落叶松林天然更新潜在的生态学机制提供依据。[方法]以关帝山华北落叶松林为研究对象,设置3 种林分密度类型,分析不同林分密度林下更新苗组成、结构及空间格局特征。[结果]中密度(825~875 株·hm-2)华北落叶松林林下更新状况良好,更新苗数量明显增多(3500 株·hm-2),低龄级更新苗较多,高龄级的更新苗相对较少,更新苗龄级结构呈连续及相对稳定的分布;而低密度(575~700 株·hm-2)和高密度(1000~1125 株·hm-2)华北落叶松林下更新不佳,更新苗数量明显减少(≤1250 株·hm-2),高密度林下更新苗龄级结构间断,幼苗到幼树的过渡不连续。从更新苗的生长情况看,低密度林分中林下更新苗的株高、地径生长表现最好。更新幼树中的优势树种在3 种密度的林分中均呈现明显的集聚分布。低密度和中密度华北落叶松林下更新幼苗的空间异质性强,高密度林下更新幼苗的空间异质性弱。更新幼苗空间变异主要由结构性因素引起。[结论]林分密度过密或过疏都不利于华北落叶松更新,及时进行密度调控有利于华北落叶松林的可持续发展。
关键词:华北落叶松;林分密度;更新苗;格局特征
中图分类号:S791.18 文献标识码:A 文章编号:1671-8151(2024)02-0110-09
森林更新是森林种群自我繁衍、延续的途径[1-2],直接影响森林资源的维持和永续经营[3],在群落结构稳定及演替中发挥重要作用。森林更新中天然更新作为一种自然恢复机制,具备较高的生态适应性和相对较低的成本[4]。幼苗幼树是天然更新过程中种群发展的主要载体,其存活和生长状况受到多种生物因素和非生物的影响,主要包括林分因子、竞争关系、光照、微地形、土壤质量等[5-10]。
林分密度作为影响森林天然更新重要因素的林分因子,往往通过对林内微环境、灌草植被、凋落物及土壤理化性状等的影响[11-14],从而影响林分发育、林分质量、蓄积量、树种更迭及个体分布。贾亚运等[15]探讨造林密度对杉木幼林生长的影响,结果表明,杉木林分平均树高、胸径、单株材积和蓄积量均随着造林密度的增大呈现先增大后减小的规律。欧芷阳等[16]研究密度对蚬木天然更新的影响,发现乔木密度的增加,使幼苗数量减少,有利于幼苗地径和株高生长。陈聪琳等[17]调查不同密度路域巨桉人工林群落结构和物种多样性,结果显示,高密度林分会限制小胸径级和高度级个体生长,群落逐渐衰退。冯燕辉等[18]研究不同密度华北落叶松林林下枯落物及土壤特征,发现适中的林分密度可改善土壤结构,利于土壤养分积累,促进林木更新和生长发育。米爽等[19]比较不同抚育采伐强度的针阔混交林幼苗更新状况后发现,土壤温度随林分密度的降低而增加,林下生境得以改善,促进更新苗生长。这些研究主要从林分密度影响林内生境,进而影响林下更新的角度进行,但对不同林分密度条件下幼树幼苗的更新状况研究相对较少。
华北落叶松是华北地区寒温带针叶林的主要树种之一,是水源涵养和水土保持的重要森林类型。因此,探究华北落叶松的更新特征, 对华北落叶松林的可持续经营,维持森林群落的稳定性具有重要意义。本研究分析关帝山不同密度华北落叶松林下更新苗组成、结构及空间格局特征,以期为深入探测华北落叶松林天然更新潜在的生态学机制提供依据。
1 研究区概况
研究地区位于山西省关帝山林区(37º45′~37º59′N,111º24′~111º37′E),海拔1800~2831 m。本区属温带大陆性气候,年均温4. 3 ℃ ,最低气温−12. 4 ℃,最高气温18 ℃,无霜期平均为100~130 d,年均降水量825 mm。土壤为山地棕壤。研究区主要乔木有华北落叶松(Larix principisrupprechtii)、白杄(Picea meyeri)、青杄(Picea wil⁃sonii)、油松(Pinus tabulaeformis)、红桦(Betula al⁃bosinensis)、白桦(Betula platyphylla)、山杨(Popu⁃lus davidiana)等。常见灌木有:北京花楸(Sorbusdiscolor)、金花忍冬(Lonicera chrysantha)、刚毛忍冬(L. hispida)、毛榛(Corylus mandshurica)、胡枝子(Lespedeza bicolor Turcz)、山刺玫(Rosa davu⁃rica Pall. )等。常见草本有:紫菀(Aster tatari⁃cus)、老鹤草(Geranium wilfordii)、东方草莓(Fragaria orientalis) 、小红菊(Dendranthemachanetii)、黑柴胡(Bupleurum smithii Wolff)、瓣蕊唐松草(Thalictrum petaloideum)等。
2 研究方法
2. 1 样地设置
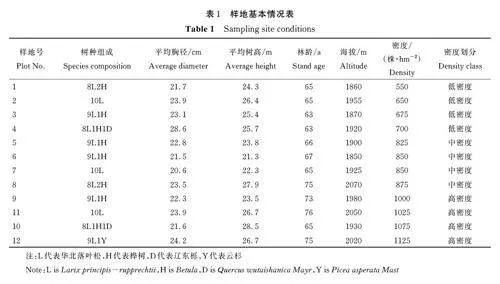
2020 年6 月-9 月,在全面踏查和样地实测基础上,选择立地条件相似,密度梯度不同的华北落叶松纯林标准样地12 个(20 m×20 m),设3 种林分密度类型,即低密度(575~700 株·hm-2)、中密度(825~875 株·hm-2)、高密度(1000~1125 株·hm-2),每个类型4 个样地,重复数为4。样地调查包括:乔木树种种类、数量、胸径、树高,用生长锥测定其林龄;林下更新苗的数量、地径、高度、林龄;样地基本情况见表1。
2. 2 群落调查
(1)分树种进行每木检尺,记录胸径、树高等生长指标。同时在每个样方内将西南角标记为坐标原点(0,0),建立统一坐标系,将东西方向标记为X轴,南北方向为Y轴,记录立木的坐标。(2)将样地分成5 m×5 m 样方,调查样方内更新苗。统计幼树和幼苗种类和株数,记录苗高、地径和年龄,并记录空间位置坐标。幼树指直径10 cm 以下,树高2 m 以上的树木;幼苗指树高≤2 m 的树木。
2. 3 数据分析方法
2. 3. 1 方差均值比法
运用方差均值比(S2/X)法,分析更新幼树分布特征:S2/X=1 时,随机分布;S2/Xlt;1 时,均匀分布;S2/Xgt;1 时,聚集分布。
2. 3. 2 变异函数分析方法
依据变异函数分析[20],结合Kriging 插值,绘制Kriging 插值图,分析更新苗空间格局特征[21]。
3 结果与分析
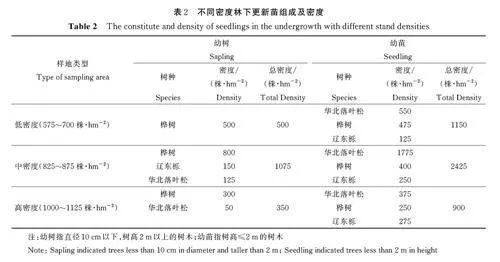
3. 1 不同林分密度林下更新苗组成及密度
不同林分密度林下更新苗组成及密度见表2。可见,不同林分密度林下更新苗组成相同,均出现华北落叶松、桦树和辽东栎更新苗。但从更新密度来看,随着林分密度增加,更新苗密度呈先增加后减少趋势,中密度(825~875 株·hm-2)时更新苗密度达最大,呈中密度林分更新苗(3500 株·hm-2)gt;低密度林分更新苗(1650 株·hm-2)gt;高密度林分更新苗(1250 株·hm-2)的趋势。
3. 2 不同林分密度林下更新苗年龄结构特征
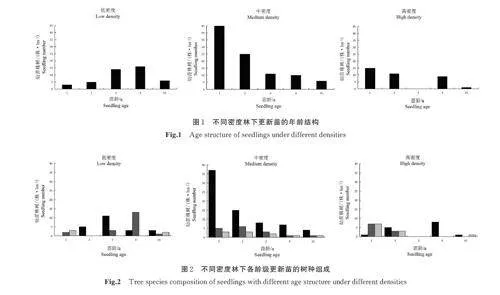
不同林分密度林下更新苗的年龄结构见图1。样地调查结果显示,林下更新幼苗年龄范围在1~8 a,由于5 a 以上幼苗更新数量较少,将5~8 a 更新苗归为同一组(≥5 a)。由图1 可以看出,低密度林分更新苗种群的年龄结构呈偏正态分布,低龄级和高龄级更新苗的个体数较少,3~4 a 更新苗数量远大于其它年龄段,个体数以4 a 最多,为36. 4%。中密度林分中,低龄级更新苗占总株树的72. 2%,高龄级的更新苗相对较少,呈逐渐减少指数型的龄级结构。高密度林分中,更新苗个体数量相对较少,林下1~2 a 更新苗占绝大多数,更新苗在2 a 之后急剧减少,出现部分更新苗龄级缺失现象。由图2 可以看出,林下更新苗以华北落叶松为主,伴随有少量的桦树和辽东栎。
3. 3 不同林分密度林下更新苗生长结构特征
林下更新苗的株高和地径分布见图3。由图3 可以看出,更新苗的株高和地径分布在不同密度林分中差异明显。更新苗的平均株高在低密度林分最大,为38 cm,其次是中密度林分、高密度林分,分别为26、21 cm,更新苗的平均株高表现为低密度gt;中密度gt;高密度的倒J 型曲线。更新苗的最大平均地径出现在低密度林分中,为0. 42 cm,明显高于中密度(0. 25 cm)和高密度林分(0. 22 cm),具体表现为:低密度gt; 中密度gt; 高密度。
林下各树种更新苗的株高和地径分布见图4。由图4 可以看出,各树种更新苗的平均地径和株高在不同密度林分中差异显著。平均地径和株高都在华北落叶松更新苗中出现最大值,说明华北落叶松幼苗在更新苗中处于优势地位。各树种更新苗地径和株高均表现为低密度gt;中密度gt;高密度的倒J 型曲线。
3. 4 不同林分密度林下更新格局特征
3. 4. 1 林下更新幼树的分布特征
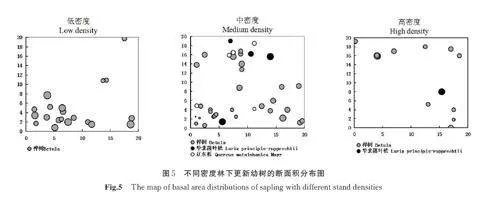
每个样地16 个5 m×5 m 的小样方,分样地计算小样方内更新幼树胸高直径分布,见表3。计算不同树种更新幼树S2/X 比值,分析其分布特征。从表3 可以看出,各更新幼树在3 种林分密度下的方差均值比均大于1,呈集聚分布。
从更新幼树分布情况来看(图5),3 种林分密度林下,优势树种均呈集聚分布。同一更新树种在一地区集聚分布,其它地区散生或无分布。不同更新树种呈现出明显的镶嵌分布。更新幼树中,华北落叶松胸高断面积大,呈零星状分布;桦树和辽东栎胸高断面积虽不大,但其密度却很大,表现出显著的聚集性。
3. 4. 2 林下更新幼苗的空间分布格局分析
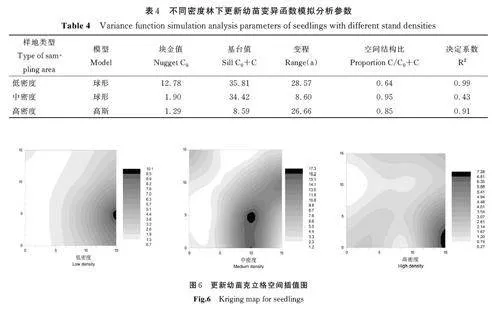
对不同密度林下更新幼苗进行变异函数分析,进行模型拟合(表4)。依据模型参数,分析更新幼苗的空间格局特征。
从表4 可以看出,低密度林分中,林下更新苗基台值为35. 81,空间结构比为0. 64,更新幼苗中64% 的空间变异由结构性因素引起,36% 的空间变异由随机性因素引起。高密度林分中,更新苗的基台值为8. 59,空间结构比为0. 85,更新幼苗中85% 的空间变异由结构性因素引起,15% 的空间变异由随机性因素引起。随着林分密度的增大,林下更新苗基台值减小,空间结构比增大,结构性因素对空间变异中的影响增大,随机性因素作用减小。
基于变异函数分析,估计Kriging 插值,绘制更新幼苗Kriging 插值图(图6),评价更新幼苗的空间分布格局。从图6 可以看出,低密度和中密度华北落叶松林下更新幼苗的空间异质性强,高密度林下更新幼苗的空间异质性弱。随着林分密度的增加,林下更新苗的空间异质程度降低。
4 讨论
4. 1 林分密度对更新苗组成及密度的影响
林分密度是影响森林天然更新的重要因子,密度变化对林下光照强度、水分含量、枯落物蓄积量、土壤养分含量等生态因子均有很大的影响,进而一定程度影响林内种子萌发和幼树生长。本研究表明,中密度华北落叶松林更新状况良好,过疏和过密林分更新状况不佳。这与康冰等[22]、张希彪[23]和卫舒平[24]研究结果一致。林分密度过低,林下太阳辐射强度强,影响种子发芽和灌草植物的光照、水分和养分竞争,抑制幼苗存活。林分密度增大,林下太阳辐射强度降低,促进更新苗存活。然而林分密度过大,林下种间、种内竞争加强,林木生长受到抑制,导致死亡率增加。
4. 2 林分密度对更新苗年龄结构的影响
更新种群的更新密度和年龄结构反应种群的更新潜力和延续能力[25-26]。本研究中,低密度林分更新苗的年龄结构呈偏正态分布;中密度林分的更新苗数量最多,而且从低龄级到高龄级更新苗的个体数呈倒J 型分布,表现出了更为稳定的更新结构[27];高密度林分中的更新苗个体数量相对较少,甚至部分龄级没有更新苗,呈现出有间隔的年龄波。这与王旭刚[28]研究结果一致。低密度林分,荫蔽程度较低,更新苗初期生长遭遇强烈光照,导致低龄级更新苗存活率低。中密度林分,荫蔽程度与光照资源相对平衡,更新苗存活率高,数量多,但随着更新苗对光照、水分和养分等需求增大,种群发生自疏和它疏,死亡率增大,个体数量递减。高密度林分,更新苗接受不到充足的阳光,从而影响更新苗的存活。
4. 3 林分密度对更新苗生长结构的影响
不同林分密度内林内资源不同[29-30],间接影响更新苗的生长[31]。本研究中林下更新苗的株高和地径在不同密度林分中差异明显。更新苗的生长指标(株高、地径)在低密度林分中表现最好。在低密度林分中,更新苗能够获得充足的伸展空间和较好的光照条件,另外生长条件良好,使得种内竞争小,营养物质多,更新苗不会受到养分、水分以及生存空间等资源不足的限制。
4. 4 林分密度对林下更新格局的影响
森林群落空间分布格局受树种特性、物理环境、干扰等因素的影响[32],具有高度的空间异质性。本研究中不同林分密度林下更新苗的分布格局不同。更新幼树中的优势树种集聚分布。更新幼苗空间变异主要由结构性因素引起,随着林分密度的增大,更新苗空间异质强度降低。但本研究中林分种类少、研究样地有限,林下更新苗的空间分布格局研究粗浅,今后应综合各因子的作用及影响程度,深入探究林分因子的异质性及其相关性。
5 结论
分析不同林分密度林下更新苗组成、结构及空间格局特征,结果表明中密度(825~875 株·hm-2)华北落叶松林下更新良好,更新苗数量明显增多(3500 株·hm-2),低龄级更新苗较多,高龄级的更新苗相对较少,更新苗龄级结构呈连续及相对稳定的分布;而低密度(575~700 株·hm-2)和高密度(1000~1125 株·hm-2)华北落叶松林下更新不佳,更新苗数量明显减少(≤1250 株·hm-2),高密度林下更新苗龄级结构间断,幼苗到幼树的过渡不连续。从更新苗的生长情况看,低密度林分中林下更新苗的株高、地径生长表现最好。更新幼树中的优势树种在3 种密度的林分中均呈现明显的集聚分布。低密度和中密度华北落叶松林下更新幼苗的空间异质性强,高密度林下更新幼苗的空间异质性弱。更新幼苗空间变异主要由结构性因素引起。林分密度过密或过疏都不利于华北落叶松更新,及时进行密度调控有利于华北落叶松林可持续发展。但具体林分密度为多少时更有利于华北落叶松更新苗生长和更新,还有待进一步研究。
参 考 文 献
[1]Liang W J,Wei X. Factors promoting the natural regeneration
of Larix principis-rupprechtii plantation in the Lvliang
Mountains of central China[J].PeerJ,2020,8:e9339.
[2]韩有志,王政权.森林更新与空间异质性[J].应用生态学报,
2002,13(5):615-619.
Han Y Z,Wang Z Q. Spatial heterogeneity and forest
regeneration[J]. Chinese Journal of Applied Ecology,2002,13
(5):615-619.
[3]陈启民,罗青红,宁虎森,等.古尔班通古特沙漠南缘不同林龄
人工梭梭林主林层和更新层特征[J].应用生态学报,2017,28
(3):739-747.
Chen Q M,Luo Q H,Ning H S,et al. Characteristics of main
layer and regeneration layer of Haloxylon ammodendron
plantations at different ages on the southern edge of the
Gurbantunggut Desert,Northwest China[J]. Chinese Journal
of Applied Ecology,2017,28(3):739-747.
[4]Kolo H,Ankerst D,Knoke T. Predicting natural forest
regeneration:a statistical model based on inventory data[J].
European Journal of Forest Research,2017,136(5):923-938.
[5]邵佳怡,杜建会,李升发,等.高山林线生态交错区木本植物幼
苗分布特征、更新机制及其对气候变化的响应[J].应用生态
学报,2019,30(8):2854-2864.
Shao J Y,Du J H,Li S F,et al. Tree seedling distribution,
regeneration mechanism and response to climate change in alpine
treeline ecotone[J].Chinese Journal of Applied Ecology,2019,
30(8):2854-2864.
[6]Atanasso J A,Mensah S,Salako K V,et al. Factors affecting
survival of seedling of Afzelia africana,a threatened tropical
timber species in West Africa[J]. Tropical Ecology,2021,62
(3):443-452.
[7]李璟,周朵朵,陈颂,等.关帝山云杉次生林树木更新与土壤养
分的空间关联性[J].应用生态学报,2021,32(7):2363-2370.
Li J,Zhou D D,Chen S,et al.Spatial associations between tree
regeneration and soil nutrient in secondary Picea forest in Guandi
Mountains,Shanxi,China[J]. Chinese Journal of Applied
Ecology,2021,32(7):2363-2370.
[8]黄慧敏,董蓉,何丹妮,等.冠层结构和光环境的时空变化对紫
耳箭竹种群特征的影响[J]. 应用生态学报,2018,29(7):
2129-2138.
Huang H M,Dong R,He D N,et al. Effects of temporal and
spatial variation of canopy structures and light conditions on
population characteristics of Fargesia decurvata[J]. Chinese
Journal of Applied Ecology,2018,29(7):2129-2138.
[9]董莉莉,汪成成,赵济川,等.辽东山区蒙古栎次生林天然更新
特征及其影响因子分析[J].西北农林科技大学学报(自然科
学版),2023,51(1):94-101,109.
Dong L L,Wang C C,Zhao J C,et al. Natural regeneration
characteristics and influencing factors of Quercus mongolica
secondary forest in the mountainous area of Eastern Liaoning
[J]. Journal of Northwest A amp; F University (Natural Science
Edition),2023,51(1):94-101,109.
[10]Dyderski M K,Gazda A,Hachułka M,et al. Impacts of soil
conditions and light availability on natural regeneration of
Norway spruce Picea abies (L.) H. Karst. in low-elevation
mountain forests [J] . Annals of Forest Science,2018,
75(4):91.
[11]吴雪铭,余新晓,陈丽华,等.间伐强度对坝上樟子松林下持
水能力的影响[J].应用生态学报,2021,32(7):2347-2354.
Wu X M,Yu X X,Chen L H,et al. Effects of thinning
intensity on the understory water-holding capacity of Pinus
sylvestris var. mongolica plantation in the Bashang Area of
North China[J].Chinese Journal of Applied Ecology,2021,32
(7):2347-2354.
[12]林立文,邓羽松,李佩琦,等.桂北地区不同密度杉木林枯落
物与土壤水文效应[J]. 水土保持学报,2020,34(5):200-
207,215.
Lin L W,Deng Y S,Li P Q,et al.Study on the effects of litter
and soil hydrology of different density Cunninghamia
lanceolata forests in northern Guangxi[J]. Journal of Soil and
Water Conservation,2020,34(5):200-207,215.
[13]卜瑞瑛,梁文俊,魏曦,等.不同林分密度华北落叶松林的土
壤养分特征[J].森林与环境学报,2021,41(2):140-147.
Bu R Y,Liang W J,Wei X,et al. Soil nutrient characteristics
of Larix principis-rupprechtii forest with different stand
densities[J].Journal of Forest and Environment,2021,41(2):
140-147.
[14]张洋洋,周清慧,许骄阳,等.林分密度对马尾松林下植物与
土壤种子库多样性的影响[J].应用生态学报,2021,32(7):
2355-2362.
Zhang Y Y,Zhou Q H,Xu J Y,et al.Impacts of stand density
on diversity of understory plant and soil seed banks in a Pinus
massoniana plantation [J] . Chinese Journal of Applied
Ecology,2021,32(7):2355-2362.
[15]贾亚运,何宗明,周丽丽,等.造林密度对杉木幼林生长及空
间利用的影响[J].生态学杂志,2016,35(5):1177-1181.
Jia Y Y,He Z M,Zhou L L,et al.Effects of planting densities
on the growth and space utilization of young Cunninghamia
lanceolata plantation[J].Chinese Journal of Ecology,2016,35
(5):1177-1181.
[16]欧芷阳,庞世龙,谭长强,等.林分结构对桂西南蚬木种群天
然更新的影响[J].应用生态学报,2017,28(10):3181-3188.
Ou Z Y,Pang S L,Tan C Q,et al. Effects of forest structure
on natural regeneration of Excentrodendron hsienmu population
in Southwest Guangxi, China[J]. Chinese Journal of Applied
Ecology,2017,28(10):3181-3188.
[17]陈聪琳,孙一淼,龚利梅,等.不同密度路域巨桉人工林群落
结构和物种多样性[J]. 森林与环境学报,2022,42(1):
20-28.
Chen C L,Sun Y M,Gong L M,et al. Community structure
and species diversity of Eucalyptus grandis plantations with
different stand densities in road area[J]. Journal of Forest and
Environment,2022,42(1):20-28.
[18]冯燕辉,梁文俊,魏曦,等.不同林分密度华北落叶松林枯落
物及土壤特征分析[J]. 西南师范大学学报(自然科学版),
2021,46(3):179-187.
Feng Y H,Liang W J,Wei X,et al. On litter and soil
characteristics of Larix principis-rupprechtii forest with
different stand densities[J]. Journal of Southwest China
Normal University (Natural Science Edition),2021,46(3):
179-187.
[19]米爽,宋子龙,秦江环,等.抚育采伐对吉林蛟河针阔混交林
幼苗更新的影响[J]. 北京林业大学学报,2019,41(5):
159-169.
Mi S,Song Z L,Qin J H,et al. Effects of tending thinning on
seedling regeneration in a mixed conifer-broadleaf forest in
Jiaohe,Jilin Province of northeastern China[J]. Journal of
Beijing Forestry University,2019,41(5):159-169.
[20]李颖,张婕,郭东罡,等.基于大样地油松种群的地统计学分
析[J].植物科学学报,2015,33(2):158-164.
Li Y,Zhang J,Guo D G,et al.Geostatistical analysis of Pinus
tabulaeform is population spatial patterns based on large plots
[J].Plant Science Journal,2015,33(2):158-164.
[21]李艳丽,杨华,亢新刚,等.长白山云冷杉针阔混交林天然更
新空间分布格局及其异质性[J]. 应用生态学报,2014,25
(2):311-317.
Li Y L,Yang H,Kang X G,et al. Spatial heterogeneity of
natural regeneration in a spruce-fir mixed broadleaf-conifer
forest in Changbai Mountains[J]. Chinese Journal of Applied
Ecology,2014,25(2):311-317.
[22]康冰,王得祥,崔宏安,等.秦岭山地油松群落更新特征及影
响因子[J].应用生态学报,2011,22(7):1659-1667.
Kang B, Wang D X, Cui H A, et al. Regeneration
characteristics and related affecting factors of Pinus
tabulaeformis secondary forests in Qinling Mountains[J] .
Chinese Journal of Applied Ecology,2011,22(7):1659-1667.
[23]张希彪,上官周平,王金成,等.子午岭人工油松林群落更新
特征及影响因子[J].山地学报,2014,32(5):561-567.
Zhang X B,Shangguan Z P,Wang J C,et al.The regeneration
characteristics and affecting factors of Pinus tabulaeformis
artificial forests of ziwu mountains,China[J]. Mountain
Research,2014,32(5):561-567.
[24]卫舒平,梁文俊,魏曦,等.不同密度华北落叶松林天然更新
及其影响因子[J].应用生态学报,2022,33(10):2687-2694.
Wei S P,Liang W J,Wei X,et al. Natural regeneration of
Larix principis-rupprechtii plantations with different densities
and its influencing factors[J]. Chinese Journal of Applied
Ecology,2022,33(10):2687-2694.
[25]张婷,张文辉,郭连金,等.黄土高原丘陵区不同生境小叶杨
人工林物种多样性及其群落稳定性分析[J].西北植物学报,
2007,27(2):340-347.
Zhang T,Zhang W H,Guo L J,et al. Species diversity and
community stability of Populus simonii plantations in different
habitats in hilly area of the Loess Plateau[J]. Acta Botanica
Boreali-Occidentalia Sinica,2007,27(2):340-347.
[26]Moktan M R,Gratzer G,Richards W H,et al.Regeneration of
structure of mixed conifer forests under single-tree harvest
management in the western Bhutan Himalayas[J]. Forest
Ecology and Management,2009,258(3):243-255.
[27]姜冬冬,林建勇,李娟,等.濒危植物闽楠幼苗更新及环境解
释[J].西北植物学报,2023,43(11):1969-1974.
Jiang D D,Lin J Y,Li J,et al. Characteristics and
environmental interpretation of seedling regeneration in the
endangered species Phoebe bournei[J].Acta Botanica Boreali-
Occidentalia Sinica,2023,43(11):1969-1974.
[28]王旭刚.关帝山不同密度天然次生针叶林林下针叶树更新苗
特征研究[D].太谷:山西农业大学,2018.
Wang X G. Reaserch on the characteristics of regeneration
seedlings in different density of natural secondary coniferous
forest in Guandi Mountain[D]. Taigu:Shanxi Agricultural
University,2018.
[29]薛立,傅静丹.影响植物竞争的因子[J].中南林业科技大学
学报,2012,32(2):6-15.
Xue L,Fu J D.A review on factors affecting plant competition
[J]. Journal of Central South University of Forestry amp;
Technology,2012,32(2):6-15.
[30]Maherali H,DeLucia E H.Influence of climate-driven shifts in
biomass allocation on water transport and storage in ponderosa
pine[J].Oecologia,2001,129(4):481-491.
[31]陈圣宾,宋爱琴,李振基.森林幼苗更新对光环境异质性的响
应研究进展[J].应用生态学报,2005,16(2):365-370.
Chen S B,Song A Q,Li Z J. Research advance in response of
forest seedling regeneration to light environmental
heterogeneity[J]. Chinese Journal of Applied Ecology,2005,
16(2):365-370.
[32]董灵波,田栋元,刘兆刚.大兴安岭次生林空间分布格局及其
尺度效应[J].应用生态学报,2020,31(5):1476-1486.
Dong L B,Tian D Y,Liu Z G.Spatial distribution pattern and
scale effect of secondary forests in Daxing’anling,China[J].
Chinese Journal of Applied Ecology,2020,31(5):1476-1486.
(编辑:郭玥微)
