
长江口南支长江江豚栖息地水生生物群落结构
2024-06-03 21:59:36吴建辉丁玲赵修江
水产科技情报 2024年3期
关键词:群落结构
吴建辉 丁玲 赵修江


摘 要:为探究长江口长江江豚的栖息地选择偏好性,根据2022年夏季和秋季长江口南支长江江豚栖息地的水生生物调查数据,采用相对重要性指数、多样性指数以及丰度/生物量曲线(ABC曲线)等方法,研究分析了该水域水生生物多样性和群落结构特征。结果显示:(1)2022年夏季共采集水生生物9种,优势种为长吻鱼危(Leiocassis longirostris)、安氏白虾(Palaemon annandalei)和光泽黄颡鱼(Pelteobagrus nitidus),秋季共采集水生生物13种,优势种为河蚬(Corbicula fluminea)、长吻鱼危和安氏白虾;(2)通过多样性指数发现,秋季该水域的水生生物多样性水平整体高于夏季,夏季东风西沙水域的多样性水平高于青草沙水域,秋季则相反;(3)ABC曲线结果表明,夏季水生生物群落处于未干扰状态(W=0.196),秋季水生生物群落则处于严重干扰状态(W=-0.069)。结果表明:长江口南支长江江豚栖息地水域水生生物多样性水平仍较低,群落结构特征和优势种存在明显季节性差异,群落稳定性较低。水域内可供长江江豚选择的饵料种类较少且资源量较低,建议加强对该区域水生生物资源的养护管理。
关键词:长江口南支;长江江豚;水生生物;群落结构
长江江豚(Neophocaena phocaenoides asaeorientalis)是世界唯一已知的江豚淡水亚种,主要分布于长江中下游干流以及大型通江湖泊洞庭湖和鄱阳湖[1-2]。大量研究表明,自20世纪80年代以来,长江江豚的种群规模不断下降,目前数量已不足1 000头[3-5]。2013年世界自然保护联盟(IUCN)将长江江豚的濒危等级从“濒危”(EN)升至“极度濒危”(CR)。2021年国家林业和草原局、农业农村部公布的《国家重点保护野生动物名录》,将长江江豚由国家二级保护野生动物升级为国家一级保护野生动物,实施重点保护与管理。
长江口位于长江与东海、黄海的交汇处,在长江冲淡水、台湾暖流、黄海冷水团和苏北沿岸水等多种水流的交互作用下,该水域饵料生物资源丰富[6]。姚思聪等[7]的研究表明,长江口南支从崇明岛西端至青草沙附近水域是长江江豚活动的热点水域。近年来,在人类工业活动和渔业捕捞等因素的影响下,长江口渔业资源结构发生了较大变化,生物多样性趋于简单化,部分物种呈现低龄化、小型化的趋势,群落稳定性日益降低[8-12]。长江江豚主要的摄食对象为小型鱼类、大型鱼类幼体以及虾类[13-14],而生物群落结构的简单化会使得餌料资源密度下降,进而导致长江江豚出现缺食争食等现象[15-16]。
本研究选用2022年夏秋季的调查资料来分析长江口南支水域水生生物的种类组成、优势种以及多样性的变化特征,旨在明晰该水域的生物群落结构变化,了解长江江豚饵料生物的资源状况,并在长江大保护的背景下为长江口南支水域的长江江豚保护和水生生物资源养护管理提供科学依据。
1 材料和方法
1.1 数据来源
本研究于2022年夏季(7—8月)和秋季(10—11月)在长江口开展桁杆拖网水生生物资源调查。调查区域为崇明岛西端的东风西沙水域至上海长江大桥附近的青草沙水域长江口南支水域,采样站位共计4个,具体坐标信息见表1。监测船持有专项(特许)渔业捕捞许可证,采样网具为桁杆拖网,网口长和宽分别为6.0 m和1.8 m,网身长11.0 m,网身网目2 cm,囊网数2个,囊网网目1 cm。监测期间,监测船以2 kn的速度拖曳1顶网具在监测水域水底拖行30 min,获取监测生物样。调查及样品保存方法参照《海洋调查规范》(GB/T 12763—2007)开展,种类鉴定参照《长江口鱼类》[17]和《长江河口大型底栖无脊椎动物》[18]。
1.2 数据处理和分析
1.2.1 优势种分析
采用相对重要性指数(index of relative importance,IRI))确定生物种类在群落中的重要性[19]。计算公式为:
IRI=(Ni+Wi)×Fi(1)
式(1)中,IRI为相对重要性指数,Ni为物种i的数量占总数量的百分比,Wi为物种i的生物量占总生物量的百分比,Fi为物种i出现的站位数占总站位数的百分比。本研究中,IRI≥1 000为优势种,1 000>IRI≥100为重要种,100>IRI≥10为常见种,IRI<10为少见种[20]。
1.2.2 多样性分析
本研究采用Margalef丰富度指数(D)、Pielou均匀度指数(J')和Shannon-Wiener多样性指数(H')[21-23]。由于不同种类间个体差异较大,用生物量表示的多样性更能体现各种类之间的能量分布,因此采用生物量进行多样性指数的计算,计算公式为:
J'=H'/lnS(2)
D=(S-1)/lnW(3)
H'=-∑PilnPi(4)
式(2)~(4)中,J'为Pielou均匀度指数,S为种类数,D为Margalef丰富度指数,W为总生物量,Shannon-Wiener多样性指数(H'),Pi为物种i的生物量占总生物量的比例。
1.2.3 ABC曲线
丰度/生物量曲线(abundance-biomass comparison curves,ABC曲线)基于生物进化r-选择和k-选择理论来评估群落受干扰的状态[24]。生物量优势度曲线高于数量优势度曲线,表示群落中以k-选择种类为主,处于未干扰状态;生物量和数量优势度曲线出现相交,表明群落中k-选择种类数量将逐渐减少,r-选择种类的数量逐渐增多,处于中等干扰状态;数量优势度曲线高于生物量优势度曲线,表明群落中以r-选择种类为主,处于严重干扰状态。本研究采用PRIMER 6.0软件绘制ABC曲线,统计量W值表示生物量与数量的相对关系,当生物量优势曲线在数量优势度曲线之上时,W为正,反之W为负,计算公式如下:
W=∑Si=1(Bi-Ai)50(S-1)(5)
式(5)中,Bi和Ai为ABC曲线中种类序号所对应的生物量和数量的累积百分比,S为出现物种数。
采用EXCEL 2017和PRIMER 6软件进行以上数据分析。
2 结果
2.1 种类组成
2022年夏季共采集水生生物9种,隶属于3纲6目7科9属(见表1)。其中软甲纲2种,均为十足目;瓣鳃纲仅有河蚬1种,属真瓣鳃目;其余均为硬骨鱼纲,分别为鲇形目和鲈形目各2种,鲱形目和鲤形目各1种。
秋季共采集水生生物13种,隶属于3纲8目11科13属(见表1)。其中软甲纲5种,均为十足目;硬骨鱼纲7种,分别为鲈形目2种,鲇形目、鲱形目、鲤形目、鲽形目和仙女鱼目各1种;瓣鳃纲仍仅有河蚬1种。
2.2 优势种分析
由表2可见,2022年夏季水生生物优势种3种,重要种1种,常见种5种,无少见种,其中长吻鱼危相对重要性指数最高(IRI=10 258.34),安氏白虾次之(IRI=4 229.80),光泽黄颡鱼第3(IRI=1 378.16)。秋季水生生物优势种3种,重要种4种,常见种2种,少见种4种,其中河蚬相对重要性指数最高(IRI=10 339.57),长吻鱼危(IRI=2 544.95)和安氏白虾(IRI=1 342.38)虽然优势度明显下降,但仍为优势种。
2.3 群落多样性时空变化特征
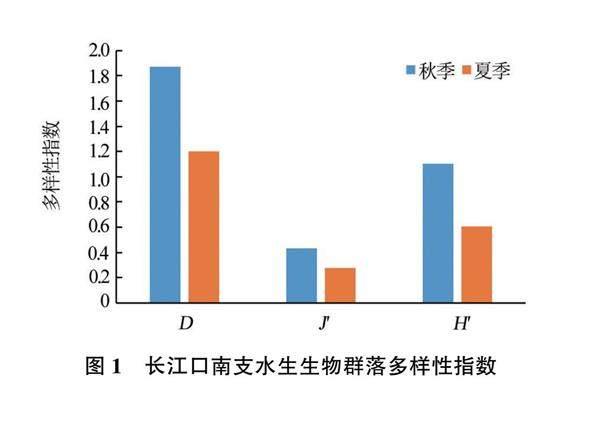
从整个调查水域来看,2022年夏季水生生物群落的Margalef丰富度指数(D)、Pielou均匀度指数(J')和Shannon-Wiener多样性指数(H')均低于秋季(见图1)。
从不同江段来看,夏季东风西沙水域的3项指数均高于青草沙水域,其中Margalef丰富度指数(D)和Shannon-Wiener多样性指数(H')高出较多(见图2);而秋季青草沙水域的3项指数均略高于东风西沙水域(见图3)。
2.4 群落干扰状况分析
2022年夏季水生生物群落的生物量优势度曲线在数量优势度曲线上方,W值为0.196,表明群落处于未干扰状态(见图4)。生物量占比前5位的物种为长吻鱼危、安氏白虾、光泽黄颡鱼、刀鲚和长蛇鱼句;数量占比前5位的物种为安氏白虾、光泽黄颡鱼、长吻鱼危、刀鲚和睛尾蝌蚪虾虎鱼。值得注意的是,以上物种中仅有长吻鱼危生物量占比高于数量占比,并且高出了66.66%,这才使得生物量优势度曲线位于数量优势度曲线的上方。
秋季水生生物群落的数量优势度曲线在生物量优势度曲线上方,W值为-0.069,表明群落处于严重干扰状态(见图4)。生物量占比前5位的物种为长吻鱼危、河蚬、三疣梭子蟹、脊尾白虾和棘头梅童鱼,数量占比前5位的物种为河蚬、安氏白虾、脊尾白虾、三疣梭子蟹和长吻鱼危。河蚬、安氏白虾和脊尾白虾等小型无脊椎动物在秋季群落中占据主导地位,其中河蚬的数量百分比较数量百分比高出39.65%。
3 讨论
3.1 种类组成及优势种演替
根据调查结果,长江口南支水域水生生物物种丰富度较低,2022年夏季捕获水生生物9种,秋季捕获13种,其中有6种在夏秋两季均有捕获,物种种类以河口性和近岸性生物为主,能够适应长江口的低盐环境。夏秋两季均以硬骨鱼类居多,且均只有河蚬1种软体类,而秋季较夏季甲壳类数量有所增多。
优势种凭借其在群落中的重要作用,在表现群落结构的特征上更具代表性。调查结果表明,该水域的夏季优势种为长吻鱼危(IRI=10 258.34)、安氏白虾(IRI=4 229.80)和光泽黄颡鱼(IRI=1 378.16),秋季优势种为河蚬(IRI=10 339.57)、长吻鱼危(IRI=2 544.95)和安氏白虾(IRI=1 342.38),每种优势种的优势度均存在较大的季节差异性,夏季到秋季呈现出由高营养级向中低营养级转变的趋势。杨柯迩[25]的研究表明,水温的变化使得长江口水域在夏秋之交有着较高的物种更替率,这与本研究结果一致。徐信等[26]研究发现,河蚬的繁殖高峰期为6—9月,因此建议捕捞季节可推迟至10月初或10月中旬,这与本研究秋季河蚬优势度大幅上升的结果相符。此外,高营养级的物种对于外界环境的变动具有较低营养级物种更差的适应性[27-28]。长吻鱼危作为温水性鱼类,其最佳生长水温为25~28 ℃[29],因此,5—10月长江口的长吻鱼危产量均保持较高水平,而11月起产量出现明显下降[30]。高营养级生物的减少更可能降低群落结构的稳定性,李健生等[8]研究发现,同水域内高營养级物种的减少会增大中低营养级物种的生存空间,从而使得群落结构发生变化。
3.2 多样性指数和群落结构稳定性
根据多样性指数分析结果,长江口南支水域2022年夏秋季物种多样性水平较低。马金等[31]研究表明,长江口南支水域鱼类资源密度较低,长江口渔业资源密度呈现“北高南低”的分布格局。造成这一现象的原因可能与长江口南支水域船只的频繁通航以及长江口深水航道治理工程的实施有关[32]。秋季的D和H明显高于夏季,群落多样性的季节性差异主要源自于环境因子的变化和生物的生长习性。该水域夏季以淡水性鱼类和河口定居性鱼类居多;而秋季则以近岸底栖鱼类居多,且伴随着甲壳类种类数的增多。这可能是因为夏秋之交水温和盐度等环境因子发生了变化。长江口水温在季节间差异显著,水温会通过影响鱼类种类组成来改变群落结构[31]。另外,盐度被认为是影响河口水域渔业资源时空分布的重要环境因子[33],物种对盐度的耐受范围决定了其生态类型以及栖息的河口水域[34]。马金等[31]研究表明,盐度在长江口夏秋两季均为GAM模型中贡献率最大的因子。通过水域间的多样性指数对比分析可以发现,夏季东风西沙水域的多样性指数明显高于青草沙水域,而秋季则相反,这可能是两个水域环境因子季节变化的差异较大导致的。
群落的优势种在一定程度上决定了生物量优势度曲线和数量优势度曲线的相对位置[35]。根据ABC曲线可以发现,该水域夏季生物量优势度曲线虽在数量优势度曲线之上,但仅有长吻鱼危的生物量占据主要优势。长吻鱼危高达84.62%的生物量占比使得夏季的ABC曲线表现为未干扰状态,而这也意味着一旦长吻鱼危的资源量出现下滑,整个群落会出现巨大波动,群落稳定性将会迅速下降。而在秋季,作为绝对优势种的河蚬数量急剧增长,其数量占比高达71.52%,这使得群落处于严重干扰的状态。河蚬作为一种具有极强环境适应能力的双壳类水生生物,相较于具有特定生态类型的鱼类,能够更轻松地适应长江口水域环境因子的季节性差异,这使得其能在夏秋之交迅速转变为优势种,从而对群落结构造成重大影响。
3.3 水生生物群落对长江江豚资源的影响
长江口南支水域是长江江豚栖息的热点水域。1984—1991年,张先锋等[1]通过长江中下游生态考察,估算出江阴—吴淞口江段有500头长江江豚。2012年,姚思聪等[7]对长江口南支为主的水域开展长江口区域江豚种群考察,共观测到江豚5群,8头个体,估算得出长江口江豚的数量为61头。2014—2016年,唐斌等[36]对长江口南支东风西沙水域的调查中发现江豚11群,68头次。长江口南支水域群落结构的稳定与否直接关乎水域内长江江豚的饵料情况。该水域水生生物多样性水平和资源量整体较低,这使得长江江豚可能会缺乏足够的饵料来源,很可能出现缺食争食等现象。另外,不同于人类牙齿功能的多样化,长江江豚只有犬齿,使得其只能咬住食物而无法咀嚼,因此长江江豚通常采用吞咽的方式进行摄食[15]。这一特点决定了长江江豚对于食物会存在较强的选择性,其在自然水域主要捕食中小型鱼类和虾类[6],而该水域夏季的优势种长吻鱼危和光泽黄颡鱼均具有硬棘和毒刺,使得长江江豚在吞咽这两种鱼的时候会消耗更多的能量,且极易在吞咽时受伤[16];而河蚬作为秋季的优势种,在群落中占据着重大比例,但其不是长江江豚的主要摄食对象,这在一定程度上压缩了其他饵料生物的生存空间,间接减少了长江江豚的食物来源。
渔业资源的养护管理是保护长江江豚的一项重要工作。目前,长江十年禁渔已经顺利开展,在极大程度上能够改善水生生物的群落结构,从而缓解长江江豚饵料缺乏的问题。除此之外,良好的栖息地环境能为长江江豚和水域内自然繁殖的生物资源提供良好的生存条件,因此对于污染的治理和水质的改善也是与保护长江江豚密不可分的工作。
参考文献
[1]张先锋,刘仁俊,赵庆中,等.长江中下游江豚种群现状评价[J].兽类学报,1993,13(4):260-270.
[2]王丕烈.江豚的形态特征和亚种划分问题[J].水产科学,1992,11(11):4-9.
[3]ZHAO X J,BARLOW J,TAYLOR B L,et al.Abundance and conservation status of the Yangtze finless porpoise in the Yangtze River,China[J].Biological Conservation,2008,141(12):3006-3018.
[4]梅志刚,郝玉江,郑劲松,等.长江江豚种群衰退机理研究进展[J].生命科学,2011,23(5):519-524.
[5]张先锋,王克雄.长江江豚种群生存力分析[J].生态学报,1999,19(4):529-533.
[6]郑元甲,陈雪忠,程家骅,等.东海大陆架生物资源与环境[M].上海:上海科学技术出版社,2003.
[7]姚思聪,樊明宁,唐文乔,等.长江河口区江豚种群调查[J].动物学杂志,2014,49(2):145-153.
[8]李建生,姜亚洲,金艳.冬季长江口渔场游泳动物群落种类组成和多样性的年代际变化[J].自然资源学报,2017,32(3):507-516.
[9]刘淑德,线薇微.长江口及其邻近水域鱼类浮游生物群落的时空格局[J].生物多样性,2009,17(2):151-159.
[10]李建生,胡芬,林楠.长江口及邻近海域春季仔、稚鱼的生态分布研究[J].南方水产科学,2015,11(1):1-8.
[11]金显仕,单秀娟,郭学武,等.长江口及其邻近海域渔业生物的群落结构特征[J].生态学报,2009,29(9):4761-4772.
[12]李建生,李圣法,丁峰元,等.长江口近海鱼类多样性的年际变化[J].中国水產科学,2007,14(4):637-643.
[13]陈佩薰,刘沛霖,刘仁俊,等.长江中游(武汉—岳阳江段)豚类的分布、生态、行为和保护[J].海洋与湖沼,1980,11(1):73-84.
[14]陈佩薰,刘仁俊,王丁.白既鱼豚生物学及饲养与保护[M].北京:科学出版社,1997.
[15]杨丙山.半自然水域中长江江豚食物选择的初步观察[J].环保科技,2019,25(5):18-21.
[16]连玉喜,杨晓鸽,万安,等.安庆西江长江江豚饵料鱼资源评估[J].长江流域资源与环境,2018,27(5):1051-1060.
[17]庄平.长江口鱼类[M].上海:上海科学技术出版社,2006.
[18]刘文亮,何文珊.长江河口大型底栖无脊椎动物[M].上海:上海科学技术出版社,2007.
[19]PINKAS L,OLIPHANT M,IVERSON I.Fish bulletin 152.food habits of albacore,bluefin tuna,and bonito in California waters[J]. Fish Bull US,1970: 1-139.
[20]程濟生.黄海无脊椎动物资源结构及多样性[J].中国水产科学,2005,12(1):68-75.
[21]PIELOU E C.Ecological diversity[J].Limnology & Oceanography,1977,22(1):174-174.
[22]SHANNON C E.A mathematical theory of communication[J].The Bell System Technical Journal,1948,27(3):379-423.
[23]MARGALEF R.Information theory in biology[J].General Systems Yearbook,1958,3:36-71.
[24]CLARKE K R,WARWICK R M.Change in marine communities:An approach to statistical analysis and interpretation[M].Plymouth:Plymouth Marine Laboratory,2001.
[25]杨柯迩.长江口鱼类群落特征及粒径结构稳定性分析[D].上海:上海海洋大学,2022.
[26]徐信,钱玲妹,张兰,等.淀山湖河蚬繁殖周期和增殖研究[J].水产学报,1987,11(2):135-142.
[27]唐启升,范元炳,林海.中国海洋生态系统动力学研究发展战略初探[J].地球科学进展,1996,11(2):160-168.
[28]金显仕,邓景耀.莱州湾渔业资源群落结构和生物多样性的变化[J].生物多样性,2000,8(1):65-72.
[29]肖明松,万全.长江长吻鱼危的生物学习性及其发展前景[J].安徽技术师范学院学报,2001,15(3):49-51.
[30]孙帼英,吴志强,陈建国,等.长江口长吻鱼危的生物学和渔业[J].水产科技情报,1993,20(6):246-250.
[31]马金,黄金玲,陈锦辉,等.基于GAM的长江口鱼类资源时空分布及影响因素[J].水产学报,2020,44(6):936-946.
[32]史赟荣,晁敏,全为民,等.2010年春季长江口鱼类群落空间分布特征[J].中国水产科学,2011,18(5):1141-1151.
[33]戴黎斌,陈锦辉,田思泉,等.基于CART算法的长江口鱼种丰富度预测[J].中国水产科学,2018,25(5):1082-1090.
[34]张衡,朱国平.长江河口潮间带鱼类群落的时空变化[J].应用生态学报,2009,20(10):2519-2526.
[35]李忠义,吴强,单秀娟,等.渤海鱼类群落结构的年际变化[J].中国水产科学,2017,24(2):403-413.
[36]唐斌,仝云云,唐文乔,等.长江口东风西沙水域江豚种群调查[J].上海海洋大学学报,2018,27(1):126-132.
Aquatic community structure of the Yangtze finless porpoise habitat in the south branch of Yangtze River Estuary
WU Jianhui1,4,5, DING Ling2,3 , ZHAO Xiujiang2
(1. Shanghai Aquatic Wildlife Conservation and Research Center,
Shanghai 200092, China; 2. Shanghai Investigation, Design & Research Institute Co.,Ltd., Shanghai 200434, China;
3. Yangtze Eco-Environment
Engineering Research Center, China Three Gorges Corporation, Wuhan 430014, China; 4. Joint Laboratory for Monitoring and Conservation of Aquatic
Living Resources in the Yangtze Estuary, Shanghai 202162, China;
5. Shanghai
Station of Aquatic Organisms Monitoring in the Yangtze River of the Ministry of Agriculture and Rural Affair,
Shanghai 200092, China)
Abstract: Based on the fishery survey data collected in the southern branch of the Yangtze River Estuary in summer and autumn of 2022,the index of relative importance,diversity indices and ABC curves were selected to analyze the structure and diversity of nekton community.A total of 9 aquatic organisms were captured in the summer of 2022,and the dominant species were Leiocassis longirostris,Palaemon Annandale,and Pelteobagrus nitidus,while 13 aquatic organisms were obtained in autumn,and the dominant species were Corbicula fluminea,L. longirostris and P. annandalei.The diversity indices showed that the diversity level in autumn was higher than that in summer.The diversity indices in the Dongfengxisha waters were higher than that in the Qingcaosha waters in summer,contrary to that in autumn.The aquatic organisms community was undisturbed (W=0.196) in summer but serious disturbed (W=-0.069) in autumn as revealed on the ABC curves.In conclusion,the diversity level of aquatic organisms in the southern branch of the Yangtze River Estuary was low.The characteristics of community structure and dominant species showed obvious seasonal differences,and the stability of community was low.There were few bait types and low resources available for the Yangtze finless porpoise in the waters.It was suggested to improve the habitat conditions and strengthen the conservation and management of fishery resources.
Key words: south branch of Yangtze River Estuary; Yangtze finless porpoise; aquatic organisms; community structure
猜你喜欢
热带农业科学(2016年11期)2017-01-21 14:59:21
热带农业科学(2016年11期)2017-01-21 14:57:08
河北渔业(2015年12期)2015-12-21 14:37:31
河北渔业(2015年11期)2015-11-18 16:39:54
河北渔业(2015年11期)2015-11-18 16:37:49
河北渔业(2015年10期)2015-10-15 11:14:57
湖北农业科学(2015年18期)2015-10-13 21:34:01
河北渔业(2015年7期)2015-07-21 11:06:52
河北渔业(2015年6期)2015-06-24 11:04:37
安徽农学通报(2015年5期)2015-03-23 23:51:20
