
4种柳树无性系生物学特性对比试验
2024-05-22 06:08:25崔长健肖杰翟雨涵赵凌全
防护林科技 2024年3期
崔长健 肖杰 翟雨涵 赵凌全

摘要:为比较不同柳树的生物学特性,以齐齐哈尔地区适生的4种无性系垂柳(CL14、CL 26、CL 21、CL 17)1年生苗为试验材料,对其物候、株高、形态特征、成活率及繁殖率进行为期2年观测研究,并利用WPS进行生长趋势分析,分析垂柳4种无性系一年生苗木物候期及生长节律。结果表明:4月上旬开始萌芽,4月中旬开始展叶,速生期于7-8月份,9月下旬生长基本停止;叶变色期于9月中旬,落叶期于10月中旬;CL 26当年生扦插苗平均高 1.50 m,分别比对照品种CL 14和CL 21大3.4%和 22.4%,一年生苗期CL 26株高生长与CL 14有显著差异,与CL 21有极显著差异。CL 26苗期生长节律表明,树高生长出现 3 个高峰在垂柳生长速生关键期加强水肥管理,可促进苗木快速生长;在成活率方面:CL 14、CL 26的成活率均为95%,CL 21的柳树成活率是50%;CL 17的成活率是50%,较前两种略逊色一筹。
关键词:柳树;物候期;生长节律;成活率;
Abstract: A 2-year observation study was conducted on the phenology, plant height, morphological characteristics, survival rate, and reproduction rate of four clones of weeping willow (cl14, cl26, cl21, cl17) suitable for growing in Qiqihar. The growth trend analysis was conducted using WPS to analyze the phenology and growth rhythm of the four clones of weeping willow. The results showed that germination began in early April, leaf expansion began in mid April, and the rapid growth period was from July to August, The growth basically stops in late September; The leaf discoloration period is in mid September, and the leaf shedding period is in mid October; The average height of the annual cutting seedlings of cl26 is 1.50 meters, which is 3.4% and 22.4% higher than the control varieties cl14 and cl21, respectively. During the one-year seedling stage, there is a significant difference in height growth between cl26 and cl14, and a very significant difference with cl21. The growth rhythm of Cl26 seedlings indicates that there are three peaks in tree height growth. Strengthening water and fertilizer management during the critical period of rapid growth of weeping willows can promote rapid growth of seedlings; In terms of survival rate, the survival rates of cl14 and cl26 are both 95%, and the survival rate of willow trees in cl21 is 50%; The survival rate of cl17 is 50%, slightly inferior to the first two.
Keywords: willow tree; Phenological period; Growth rhythm; Survival rate;
柳樹是杨柳科( Salicaceae) 柳属( Salix) 植物的泛称,落叶乔木或灌木,全球约520多种,具有速生、耐寒性强、根系比较发达、树干柔软、叶子细长等特点。中国是世界上柳树的主要分布区域之一,我国已记录了257个种122个变种33个变型[1],是世界上最大的分布区域。天然环境下,柳树种间和属间的亲缘关系密切,具有丰富的遗传多样性。柳树是一种重要的防护林、水土保持林、景观林和用材林,用途十分广泛。柳树是一种重要的防护林、保土林、景观林和用材林,用途十分广泛。在此背景下,大力发展柳树产业,对缓解木材供需矛盾,改善城市生态环境,改善城市风貌,提高城市景观质量,都有着十分重要的现实意义。柳树的垂直分布广泛[2]在发达国家,与柳树有关的研究开始得比较早,从微观[3-5]到宏观[6-12]都比较系统。但是速生、观赏性强且在黑龙江地区物候期较长的柳树品种,确是比较缺乏。
在我国平原地区,柳树是重要的造林绿化树种,它是防护林、水土保持林、风景林及用材林的重要组成树种。具有较高的生态价值与经济效益[13]。近年来,受自然与人类活动双重影响,尤其是在人类活动不断增多的情况下,对坡地进行了大规模的开垦,导致了黑土区土壤侵蚀加剧,生态环境不断恶化。同时在小流域水土不断流失的情况下,对柳树的需求量又逐渐增大,在水土保持过程中可以起到不可替代的作用。
因此本文针对黑土区水土流失严重、土壤退化以及水土保持林建设中存在的主要问题,以4种柳树为研究对象,以防治水土流失为重点,筛选出最优柳树新品种为目的,以探索最优的水土流失治理模式为目标,从水土保持林植物材料对比入手,开展水土保持林优良品种的物候期及生长节律对比研究,进行筛选其适合黑土地区生长,并能够速生的新材料,应用于小流域治理当中,并通过扦插、组织培养等方式对优良品种进行扩繁和区域化试验,筛选出最具有地方特色,集生态、经济、于一身的优良水土保持新材料。
1 研究区概况
富拉尔基区位于北纬47°12',东经123°40′。年平均气温4℃,属于中温带大陆性季风气候区。春季风大少雨,夏季炎热多雨,秋季降温迅速,冬季漫长寒冷干燥,多西北风。年平均气温0.7℃,最高气温37.5℃,最低气温-39℃,≥10℃年积温为2598.1℃,无霜期为115天左右,早霜9月,晚霜10月,年平均降水量396mm左右,降水时间多集中在7-8月份,解冻期4月下旬,结冻期11月中旬。土壤为黑钙土、砂土。平均土层厚度50cm,pH值中性偏碱。富拉尔基区有一江一河和众多湖泡,水域宽广。地表径流量达0.085亿m3,嫩江富区江段年平均流量为172亿m3,为区内工农业用水,提供足够的水源。柳树区域化试验布局在位于齐齐哈尔市富拉尔基区的黑龙江省林科院齐齐哈尔分院试验基地和齐齐哈尔市绿昊苗圃。
2研究方法
选择CL14、CL 26、CL 21、CL 174个柳树无性系进行定点定位定期观察。
将4份柳树无性系2年生枝条截成长15cm的插穗,选取均匀一致的插穗扦插于试验地。扦插后浇透水,正常管理,当苗高20cm时,进行抹芽,保留壮芽1个,开始观察其物候期,形态特征及生长节律,平均10d进行1次测量并拍照记录。同时进行标记,保证每次测量的是同一株树苗。
釆用wps2023 Excl整理并统计试验数据,用SPSS 22对试验数据作分析。
3结果与分析
3.1形态特征
4种柳树均为乔木,而且是雄株,树干非常笔直,树冠呈长椭圆形,枝间分枝角为26.5度,冠幅较大,平均 2.85m。树干下半部分纵向有裂痕,颜色为灰色。苗期树干上半部分部树皮光滑,颜色为深红色,4种柳树的老枝都比较光滑,颜色呈灰绿色或浅褐色。芽的形状为先端钝圆,锐角三角形,同时有少量细绒毛。叶呈针形,先端渐尖或尾尖,基部呈倒三角形,较为平展。短枝上的叶长3.5-11.0 cm,乔木,雄株,主干直立,冠长卵状,分枝角为26.5°,冠幅为2.72m。主干基部有浅浅的纵纹,灰褐色。幼年树干上半部的树皮平滑,呈暗红色,老龄的枝条平滑,呈灰绿或淡棕色。花蕾呈三角形,顶端钝,具绒毛。叶披针状,顶端渐尖,或有尾尖,底部呈楔状,扁平。短枝叶片的长度在3.5~11.0cm之间,宽0.4 ~- 1.7 cm,叶柄长0.25~0.65cm,有绒毛和1~2个腺体的基部。初生的叶片具托叶,呈倾斜的三角形或直线状,随后渐渐脱落。长枝叶叶长5.3~16.8 cm,宽 0.5~2.4cm,叶柄长0.3~0.9 cm。花与叶同时开放,雄花絮圆柱形,黄绿色,花絮长 1.8~2.6cm,粗6~7 mm,花药为卵形,黄绿色,花粉淡黄色;盾状苞片,顶部渐尖或尾部尖,黄绿色,被一根比苞片更长的绒毛覆盖,在背面和腹部各有一腺体。
3.2四种柳树物候期观测结果
项目组分别于2021年-2022年的柳树生长期,在齐齐哈尔市富拉尔基区科研基地和齐齐哈尔种畜场绿浩苗圃开展了经过初选的柳树无性系物候期观测记录工作。经过2年的观测统计,4个供试柳树品种(cl26、cl21、cl14、cl17)的物候期为:4月9日-15日为芽膨大期、4月26-30日为芽开放期、4月30日-5月2日为展叶期、4月15日-4月25日为花絮出现期、5月2日-5月6日为开花始期、9月4日-9月9日为顶芽形成期、9月13日-9月20日为叶始变色期、10月7日-10月11日为叶全变色期、10月14日-10月25日为落叶始期。
在齊齐哈尔市,这4种柳树4月上旬芽会开始生长,4月中旬开始,花会进行开放,芽同时进行生长,花会在4-5天后展开叶子,然后在第2天就会开花、9月上旬生长停止,10月15日左右落叶。(表1)
3.3生长节律研究
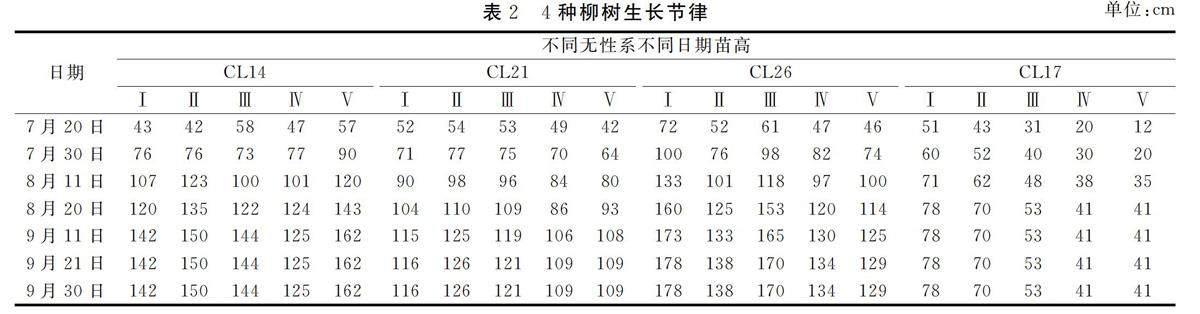
从2022年7月20日开始测定CL17,CL14,CL21,CL26株高,直至柳树落叶期。在绿昊苗圃试验地,由表2可见,CL26当年生扦插苗平均高 1.50 m,分别比对照品种CL14和CL21大3.4%和 22.4%,一年生苗期CL26株高生长与CL14有显著差异,与CL21有极显著差异。CL26苗期生长节律表明,树高生长出现 3 个高峰,从总体上看,树高生长持续时间较长,直至9月中下旬还有生长,经苗期观察结果表明,CL26快速生长的最大原因是其快速生长的持续时间较长。
由表2可知,CL17、CL26、CL14的当年生长量均高于CL21,平均高出30%。最大的17号高出35%。由此可见其中CL14、CL26、CL17垂柳生长速率较快。
经对1年生4个无性系垂柳株高单因素方差分析(one-way ANOVA)检验,CL14与CL21差异显著(P<0.05),与其他植物差异不显著(P>0.05);CL21与其他植物差异显著(P<0.05);CL26与CL21差异显著(P<0.05),与其他植物差异不显著(P>0.05);CL17与CL21差异显著(P<0.05),与其他植物差异不显著(P>0.05)。
经每次测量后,通过WPS2023 处理可得图一四种柳树生长走势图如下:
4种不同无性系柳树,株高生长在7-8月为快速生长期,8-9月为缓慢生长期,9月中旬开始出现停滞的状态,生长曲线为典型的“S”曲线,8月中旬之前除CL17生长缓慢以外,其余生长速率几乎一样,但最后发现直至9月中下旬,CL26还在生长,同时可出现3个峰值,由此可见,CL26生长周期长是CL26速生的最主要因素。
3.4成活率及繁殖率比较研究
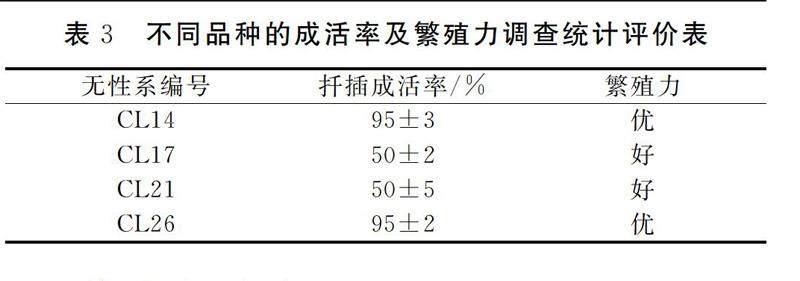
扦插2个月后调查无性系的成活率并计算,不同品种的成活率、繁殖力评价结果如表4所示。
从表3中可以发现,CL14、CL26的成活率均为95%,CL21的柳树成活率是50%;CL17的成活率是50%,较前两种略逊色一筹;
4结论与讨论
柳树物候期分为,芽生长期、芽长出期、叶开展期、花絮萌发期、芽开放期、顶芽生成期、叶初变色期、叶全变色期、落叶期等。在齐齐哈尔市,这4种柳树4月中上旬芽会开始生长,4月中旬开始,花会进行开放,芽同时进行生长,花会在4-5天后展开叶子,然后在第二天就会开花,7、8月份为速生期,9月上旬生长停止,10月15日左右落叶。
CL26当年生扦插苗平均高 1.50 m,分别比对照品种CL14和CL21大3.4%和 22.4%,一年生苗期CL26株高生长与CL14有显著差异,与CL21有极显著差异。CL26苗期生长节律表明,树高生长出现 3 个高峰,7-8月份是速生期,其生长量最快,从总体上看,树高生长持续时间较长,直至9月中下旬还有生长,经苗期观察结果表明,CL26快速生长的最大原因是其快速生长的持续时间较长。
CL26及CL14扦插成活率几乎一样的情况下,但是CL21生长速率更快,所以在大面积推广过程中,更应推广CL26。以4种柳树无性系苗木年生长节律的观测研究为基础,对4种柳树9个无性系生长期进行划分,针对不同无性系的生长阶段制定不同的栽培方案在生产中是十分必要且切实可行。速生期是植株生长的关键期[14],通过研究4种柳树的速生期,并在对应时期加强水肥管理,可有效促进柳树这种植物的快速生长。
4个垂柳新无性系均能适应本地土壤、气候等因子、生长迅速、适应性强、适宜本地大面积推广。4~5月应重点做好抗旱工作,确保苗木成活,7~8月苗木高生长高峰期,是管理的关键,应及时降渍排水、除去杂草、同时进行病虫害防治、追施肥料、创造苗木快速生长的适宜条件,确保齐苗壮苗,提高优质苗出圃率。
对CL17、CL26、CL14、CL21垂柳的形态特征、物候期及生长节律等进行了系统研究,证明其CL14、CL26、CL17垂柳生长量明显高于CL21垂柳,CL14,CL26,CL17等垂柳树可以作为建筑工程,也可以作为农用用途。结果表明,CL17、CL26和CL14三种柳树在营造速生林、三北防护林、进行低洼小流域治理和园林城市绿化等方面都有非常良好的应用前景,在我国有着很好的发展空间。其中CL26、CL17垂柳更具有速生的优势,长势更迅猛,更适合大面积推广且在短的时间内可以培养出水土保持的柳树新材料,从而在为黑土区小流域治理提供一份力量,在生产过程中,可以快速生長成木材,解决木材紧缺的情况,长成大树以后还可以应用于风景园林中,应用效果更好。
参考文献
[1] 中国科学院《中国植物志》编辑委员会.中国植物志: 第20卷 第2分册 [M].北京: 科学出版社, 1984.
[2] 任宪威.树木学[M]. 北京:中国林业出版社, 1997.
[3] Pohjonen V. Selection of species and clones for biomass willow forestry in Finland[J].Acta Foresttalia Fennica,1991,221(221):1-58.
[4] Nakai A, Y Yurugi, H Kisanuki. Stress responses in Salix gracilistyla cuttings subjected to repetitive alternate flooding and drought[J].Trees,2010,24(6):1087-1095.
[5] Tsarouhas V, U Gullberg, U Lagercrantz. Mapping of quantitative trait loci (QTLs) affecting autumn freezing resistance and phenology in Salix[J].Theoretical and applied genetics,2004,108(7): 1335-1342.
[6] R?nnberg- W?stljung A C, C Glynn, M Weih. QTL analyses of drought tolerance and growth for a Salix dasyclados × Salix viminalis hybrid in contrasting water regimes[J].Theoretical and Applied Genetics,2005,110(3):537-549.
[7] Hanley S, J Barker, J Van Ooijen, et al. A genetic linkage map of willow (Salix viminalis) based on AFLP and microsatellite markers [J].Theoretical and Applied Genetics,2002,105(6/7):1087-1096.
[8] Skvortsov A K. Willows of Russia and adjacent countries[M]. Beijing: Science Press,1999
[9] Kuzovkina Y A. Establishment and Maintenance of Living Structures Madeof Willow (Salix) Stems[J].Arboriculture & Urban Forestry,2008,34(5):290-295.
[10] Ball J, J Carle, A Del Lungo. Contribution of poplars and willows to sustainable forestry and rural (21):3-9. development[J].Unasylva,2005,56.
[11] Sviatlana T, J ?árka, M William, et al. A Genetic Study of a Salix Germplasm Resource Reveals New Insights into Relationships Among Subgenera, Sections and Species[J].Bioenerg. Res,2008,1: 67-79.
[12] Dorn R D. A systematic study of Salix section Cordatae in North America[J].Canadian Journal of Botany,1975,53(15):1491-1522.
[13] Dorn R D. A taxonomic study of Salix section Cordatae subsection Luteae (Salicaceae)[J].Brittonia,1995,47(2):160-174.
猜你喜欢
现代畜牧科技(2021年10期)2021-11-19 08:42:22
现代畜牧科技(2021年5期)2021-07-20 08:07:24
河北果树(2020年1期)2020-02-09 12:31:42
小天使·一年级语数英综合(2018年4期)2018-06-22 10:33:26
少儿科学周刊·儿童版(2017年5期)2017-06-29 22:14:38
现代农业科技(2017年2期)2017-03-16 20:29:21
现代园艺(2016年7期)2017-01-09 22:25:27
农业与技术(2016年15期)2016-11-09 07:16:25
科技视界(2016年14期)2016-06-08 18:58:03
兽医导刊(2016年6期)2016-05-17 03:50:30
