
黄鹂无齿鲹胚胎及胚后发育特征
2024-05-20 05:22黄炜坪甘松永杨少森黄锦雄赵艳飞邹翠云黄培卫张潇潇陈永南秦真东吴锦辉
水产学报 2024年4期
黄炜坪, 甘松永, 王 庆, 杨少森, 黄锦雄, 赵艳飞, 邹翠云,黄培卫, 张潇潇, 陈永南, 秦真东, 吴锦辉*, 林 蠡*
(1. 广东省水环境与水产品安全工程技术研究中心,广州市水产病害与水禽养殖重点实验室,仲恺农业工程学院动物科技学院,广东 广州 510222;2. 广东省农业技术推广中心,广东 广州 510520;3. 华南农业大学海洋学院,广东 广州 510642)
黄鹂无齿鲹(Gnathanodon speciosus)属于鲈形目 (Perciformes) 鲹 科 (Carangidae) 无 齿 鲹 属(Gnathanodon)。其体色金黄,故俗称黄金鲹,体侧具7~11 条的黑色纵向条纹[1]。黄鹂无齿鲹主要分布于我国南海,日本本州岛以南、琉球群岛、印度洋太平洋及东南亚珊瑚礁海域等地。游泳能力强,成群活动,个体中小型,常常跟随在大型鱼类后方,如鲨鱼、大型石斑鱼等。肉食性,依靠可伸缩的吻部吸食珊瑚礁中的小型无脊椎动物为生,其幼鱼期下颌具有少数齿,而成鱼下颌和犁骨部位则无齿,仅有舌面较为粗糙,同类相残比较少。黄鹂无齿鲹成鱼可达1 m 以上,肉质鲜美,可加工为刺身食用。由于其肉质比主养品种卵形鲳鲹(Trachinotus ovatus)更加细嫩,市场售价也更高。这些特征使得黄鹂无齿鲹既是良好的观赏性鱼类,又是高经济价值的食用鱼。
黄鹂无齿鲹的人工养殖以海水网箱和室内海水循环水池模式为主。目前国内只针对水族养殖的黄鹂无齿鲹的细菌性疾病进行了初步研究[2],国外主要集中在其环境因子适应性、生长性能、食性、营养需求和微卫星序列测定等方面[3-9],该鱼种的研究还处于比较初级的阶段,对于其生殖生理特性以及胚胎发育过程等的研究还较缺乏。本研究通过人工繁殖黄鹂无齿鲹,并使用显微拍摄技术对其胚胎发育和胚后发育过程进行了记录,描述了不同发育阶段的形态特征,为后续开发该品种的人工繁殖和苗种培育提供基础。
1 材料与方法
1.1 实验对象
实验用黄鹂无齿鲹成鱼购于海南陵水当地渔民捕捞的野生个体,转运至广东省农业技术推广中心大亚湾基地海水循环水系统中养殖,养殖池大小为5 m×5 m×3 m,光周期为12L∶12D,光照强度约3 000 lx、DO≥5 mg/L、pH 值 (7.66±0.17)、水温(27.75±1.38) °C、盐度32.32±0.59。亲鱼强化培育交替使用冰鲜鱼和人工配合饲料。人工配合饲料使用鳗鱼粉、鱿鱼浆、沙虫浆按照5∶3∶2的比例制作成颗粒。隔天投喂冰鲜的日本鳀(Engraulis japonicus)、沙丁鱼(Sardina pilchardus)、日本鲭(Scomber japonicus)等杂鱼,连续投喂15~30 d 后检查成熟度。实验过程中操作人员严格遵守实验动物伦理规范,并按照仲恺农业工程学院动物科技学院伦理委员会制定的规章制度执行。
1.2 亲鱼繁殖及受精卵的获得
强化培育15~30 d 后检查成熟度,使用氧化锌丁香酚水门汀麻醉后,当雄鱼能挤出精液及雌鱼腹部柔软微微隆起、生殖孔发红,转移至催产池进行人工催产。
人工催产采用两针催产法,均采用背部肌肉注射,第1 针注射促黄体素释放激素A3 (LHRHA3),剂 量 为3~5 μg/kg。12 h 后 注 射 第2 针,LHRH-A3 和绒毛膜促性腺激素(HCG)联合用药,LHRH-A3 的剂量为10~15 μg/kg,HCG 的剂量为7.5~10.0 μg/kg。注射完第2 针后12 h 通常可以获得成熟的卵。用干法受精进行人工授精操作:将成熟鱼卵放入干净的烧杯中,加入成熟的精液,用羽毛搅拌均匀,静置3 min 后加入新鲜海水,轻轻搅动5 min 后,用新鲜海水冲洗鱼卵,挑选浮性卵放入孵化桶中孵化。
1.3 受精卵的孵化、胚胎发育及胚后发育观察
将受精卵置于500 L 循环桶中孵化,同时通气保持水体充氧。受精卵在Leica DM2000 显微镜下进行观察,显微图像系统拍摄图片,记录各时期特征,每次观察取样胚胎30 粒。
仔鱼出膜后,在直径6.0 m,水深1.5 m 的圆形水泥池中进行苗种培育,饵料系列为经营养强化的褶皱臂尾轮虫(Brachionus plicatilis)、丰年虫(Artemia salina)、冻存的低盐咸水剑水蚤(Halicyclops aequoreus)、丰年虫成体及微囊颗粒饲料等。初孵仔鱼培养密度约为10 000 尾/m3,水质条件为温 度(25.59±0.21) °C、盐 度31.36±0.47、pH 值7.58±0.11、 DO≥5 mg/L、光照强度约3 000 lx。胚后发育的时间采用日龄(day post hatch, dph)进行记录,初孵的仔鱼定义为0 日龄,以此类推。胚后发育观察为每日选取25~30 尾健康仔鱼,在Leica S8AP0 体视镜下拍照,显微图像系统测量记录全长、卵黄囊大小等。
1.4 卵径的测量及胚胎发育总积温的计算方法
根据有效积温法则,胚胎发育总积温的公式:
式中,K代表总积温(h·°C);N代表完成某一发育阶段所需的时间(h);T代表发育期间的平均水温(°C);C代表该生物种类的发育起始温度(°C)。利用显微图形软件测量100 粒受精卵的直径,取这100 粒卵直径的算术平均值。
1.5 数据分析
受精卵与仔鱼可量性状数据以平均值±标准差(mean±SD)表示。
2 结果
2.1 受精卵
黄鹂无齿鲹受精卵为非黏性、圆球形的透明浮性卵,但大量受精卵聚集在一起时呈现淡黄色。受精卵平均卵径为(764.29±14.74) μm,具有1 个油球,油球平均直径为(166.32±18.28) μm。
2.2 胚胎发育
受 精 卵 在 盐 度30.26±0.67、 温 度(24.72±0.32) °C、pH 值7.46±0.12、溶 解 氧(5.13±0.33)mg/L、光照强度约3 000 lx 的海水环境中孵化,18 h 30 min 后,开始出膜。整个胚胎发育过程所需要的总积温为470.8~503.8 °C·h,黄鹂无齿鲹胚胎发育时序见表1。

表1 黄鹂无齿鲹胚胎发育特征Tab. 1 Embryonic development characteristics of G. speciosus
胚盘期 黄鹂无齿鲹的受精卵(图版Ⅰ-1)在受精后15 min,细胞质与细胞核开始从植物极定向流动向动物极,形成盘状的胚盘隆起(图版Ⅰ-2)。
卵裂阶段 受精后25 min,黄鹂无齿鲹的受精卵进行第1 次卵裂,标志着胚胎发育进入2细胞期。此时胚盘中央区域由外向内进行缢裂,均等分成2 个分裂球,这是整个胚胎发生过程中的首次细胞分裂(图版Ⅰ-3)。受精后35 min~1 h,胚胎连续进行从第2 次到第5 次的细胞分裂,即胚胎从2 细胞期快速发育到32 细胞期。每次细胞分裂所形成的分裂面均与上一轮分裂面垂直,使得胚胎整体维持球对称性,而细胞数则以几何级数翻倍增长(图版Ⅰ-4~7)。受精后1 h 10 min,随着胚盘中央区域的快速分裂,各个细胞或细胞团大小开始不均等,中心区域细胞较小,边缘区域较大。细胞之间分界面也变得不再规则,呈现出桑椹状早期胚胎的特征(图版Ⅰ-8)。15 min 后,受精卵发育到桑椹胚晚期,此时分裂球继续变小,特别是边缘区域细胞分裂数目增加,使得整个胚盘轮廓近似圆形。细胞分化初步完成,胚胎发育进入下一个更为复杂的阶段 (图版Ⅰ-9)。
囊胚阶段 受精后2 h 5 min,黄鹂无齿鲹的受精卵进入囊胚发育阶段。此时胚盘细胞极度缩小,并在一侧形成半圆形的囊胚层。同时胚盘开始向一侧移动,标志着高囊胚期的开始(图版Ⅰ-10)。此后随着时间的推移,在受精后3 h,越来越多的囊胚层表层细胞向下层及外围区域生长、扩散和迁移,囊胚腔可见,进入低囊胚期 (图版Ⅰ-11)。

图版 Ⅰ 黄鹂无齿鲹受精卵至出膜前期阶段图谱1. 受精卵,2. 胚盘形成,3. 2 细胞期,4. 4 细胞期,5. 8 细胞期,6. 16 细胞期,7. 32 细胞期,8. 桑椹胚早期,9. 桑椹胚晚期,10. 高囊胚期,11. 低囊胚期,12. 原肠胚早期,13. 原肠胚中期,14. 原肠胚晚期,15. 胚孔闭合早期,16. 胚孔闭合期,17. 头部原基形成期,18. 眼基形成期,19. 眼囊形成期,20. 心脏出现期,21. 晶状体出现期,22. 心跳期,23. 出膜前期,24. 出膜前期(侧)。Plate Ⅰ Photos of G. speciosus from fertilized eggs to pre-hatching stage 1. fertilized eggs, 2. blastodisc formation, 3. 2-cell stage, 4. 4-cell stage, 5. 8-cell stage, 6. 16-cell stage, 7. 32-cell stage, 8. early morula stage, 9. late morula stage, 10. high blastula stage, 11. low blastula stage, 12. early gastrula stage, 13. middle gastrula stage, 14. late gastrula stage, 15. early blastopore closure stage, 16. closure of blastopore, 17. head primordium formation stage, 18. ocular primordium stage, 19. ocular sac stage, 20. cardiac onsets, 21.eye lens emergence, 22. heart-beating stage, 23. pre-hatching stage, 24. pre-hatching stage (side).
原肠胚阶段 受精后3 h 35 min,原肠胚发育进入早期,此阶段可观察到两层胚层中的外层细胞开始向原卵黄囊1/3 区域发育、扩张,而尚未形成完整的胚环,仅有模糊虚影。此时出现原始的胚盾结构,但细胞分化还不明显(图版Ⅰ-12)。4 h 时,原肠胚发展到中期,细胞快速分裂增殖,外层胚层完全覆盖了卵黄囊右上方1/2 的表面,胚环边界清晰可辨,且胚盾增厚放大,细胞分化明显,预示原肠腔形成(图版Ⅰ-13)。40 min后,进入原肠胚晚期,外层细胞开始向另一侧覆盖,到达卵黄囊的2/3 左右。此时胚盾变窄,原肠胚发育结束(图版Ⅰ-14)。
神经胚阶段 受精后5 h 30 min,黄鹂无齿鲹胚胎发育进入神经胚阶段。在胚盾外层可见神经板细胞出现分化,神经芽的雏形出现,同时胚孔接近闭合(图版Ⅰ-15)。受精后6 h 10 min,胚层细胞已经几乎完全覆盖整个卵黄囊,尚存的孔隙被填平,胚胎从单层的胚盘发育成了由两层细胞构成的柱状胚体结构(图版Ⅰ-16)。1 h 后,神经板进一步发育为中空的圆筒状神经管,头部原基在膨大的柱状胚体一端成形,这标志着胚胎器官和系统开始进行高速分化,进入新的发育阶段(图版Ⅰ-17)。
器官形成阶段 受精后8 h 30 min,黄鹂无齿鲹胚胎发育进入眼原基形成期,头部两侧隆起眼芽结构的雏形。与此同时柱状胚体前端呈现铲状扁平结构,为头部特征发育奠定基础 (图版Ⅰ-18)。在受精后的9 h,眼原基的外胚层向内下陷,逐渐膨胀凸出而形成眼囊。这标志着视觉器官的关键分化已经开始(图版Ⅰ-19)。发育到受精后11 h 35 min,心脏作为第一个运动器官而出现,呈现简单的管状结构位于头部末端略为下方,这预示着胚胎循环系统形成(图版Ⅰ-20)。 13 h 30 min,在扩张的眼囊下方可观察到光学性晶状体的出现,视网膜和其他结构将在其后形成(图版Ⅰ-21)。至13 h 45 min 心脏开始有自主规律的收缩和舒张,标志着有效的血液循环系统已经建立(图版Ⅰ-22)。18 h 30 min 进入出膜前期,最后阶段胚胎整体呈 “C” 字形,心跳和血流明显,破膜孵化的最后阶段即将到来(图版Ⅰ-23~24)。

图版 Ⅱ 黄鹂无齿鲹前仔鱼期1. 初孵仔鱼(正),2. 初孵仔鱼(侧),3. 1 日龄仔鱼,4. 2 日龄仔鱼,5. 3 日龄仔鱼。Plate Ⅱ G. speciosus larvae-prelarval stage 1. newly hatched larvae (frontal), 2. newly hatched larvae (side), 3. 1 dph larvae, 4. 2 dph larvae, 5. 3 dph larvae.
2.3 胚后发育
黄鹂无齿鲹从破壳而出到尾鳍骨骼开始形成的0~20 dph 为仔鱼期,其中0~3 dph 为前仔鱼期(图版Ⅱ),是整个仔鱼发育中最为关键的阶段。刚孵化时体内卵黄尚存余,口部和消化道尚未完全形成,不能进食,依赖储存养分。在0~3 dph,一系列器官和系统快速发育,口咽开启,肠胃通路建立,与此同时眼部色素沉积,视网膜和晶状体功能加速建立,这为下一个积极摄食和快速成长的发育阶段奠定了基础。
4~20 dph 为黄鹂无齿鲹的后仔鱼期(图版Ⅲ),以卵黄囊被完全吸收作为进入该阶段的标志。后仔鱼期作为其生长发育中极为关键的一个时期,卵黄囊储备已消耗殆尽,消化道系统完善,通过主动捕食获得外部营养。同时各鳍条迅速延长,使运动更为灵活敏捷,追逐和捕食能力大幅提高。20 dph 后仔鱼进入稚鱼阶段,以脊索的完全弯曲作为进入该阶段的标志。同时各鳍和消化系统发育完善,身体大面积浸染色素,不再透明,具备一定的保护色,这标志着其由依赖型向独立型生长的转变。

图版 Ⅲ 黄鹂无齿鲹6、9、12、15 和20 dph 图谱6 日龄仔鱼,2. 9 日龄仔鱼,3. 12 日龄仔鱼,4. 15 日龄仔鱼,5. 20 日龄仔鱼。Plate Ⅲ Photos of 6, 9, 12 and 20 dph of G. speciosus larvae 6 dph larvae, 2. 9 dph larvae, 3. 12 dph larvae, 4. 15 dph larvae, 5. 20 dph larvae.
卵黄囊前仔鱼期 黄鹂无齿鲹初孵仔鱼全长为(1 520±19) μm,卵黄囊呈长椭圆形,长径为(981±12) μm,短径为(597±11) μm,位于仔鱼身体前下方,油球位于卵黄囊的最前端(图版Ⅱ-1~2)。此位置构造与其他鲹科鱼类有所不同,其他鲹科鱼类初孵仔鱼的卵黄囊一般与头部齐平或不超过头部位置,且油球位于卵黄囊中部或中前端。初孵仔鱼刚脱膜时,身体弯曲,躯体前半部分伏在卵黄囊上,呈现弯曲弓背状,腹部朝天。头部与躯干可见散点黑色素,细长肠道紧贴在腹面,眼囊与晶状体不着色(图版Ⅳ-1)。
1 dph 仔鱼:全长为(1 737±24) μm,卵黄囊体积缩小约75%,不再遮盖整个头部。仔鱼身体完全伸展,尾部提供动力,能在水中上下调整身姿,但活动能力仍较虚弱。背鳍与臀鳍基部色素沉积增加,形成色素带,眼囊开始着色(图版Ⅳ-2、7),躯干两侧颜色加深,尾部近乎透明。细长肠道尚未充气膨胀(图版Ⅱ-3、图版Ⅳ-11)。
2 dph 仔鱼:全长为(1 981±41) μm,卵黄囊体积仅余10%左右。此时能够平衡悬浮并自主游动,通过尾鳍摆动快速变换身体位置。各鳍面的发育加快,尤其眼囊区黄色素沉淀,不透明(图版Ⅳ-3)。消化道管腔扩大,轮廓清晰可见(图版Ⅱ-4)。
3 dph 仔鱼:全长为(2 126±39) μm,原有巨大卵黄囊储备已完全被吸收消化。尾端残留少量散点状色素沉淀,躯干区域颜色转为棕黄。活动行为仍较虚弱。眼部因黑色素高度富集而全黑(图版Ⅳ-4)。心脏血液循环明显。口器张开,下颌支撑骨可见。消化道贯通畅通,肠道表面出现褶皱(图版Ⅱ-5、图版Ⅳ-12)。
脊索弯曲后仔鱼期 6 dph 仔鱼:全长为(2 796±53) μm,头腹内原有的油球储备已完全被吸收。鱼鳔形成,与孵化初期相比,其游泳和避障能力获得明显提高,能够主动在水体中活动,向光源方向趋化,这揭示了视觉系统和平衡装置的逐步成熟。消化活动增多,肠前段和胃后方之间形成了生理性的弯曲(图版Ⅳ-13),鱼体躯干部分以及上下两侧体表可看到大块颜色深沉的色素细胞聚集区块,而眼囊区域因黑色素高度富集而呈现出黑色。此时仔鱼整体体色以黄褐色为主(图版Ⅲ-1)。
9 dph 仔鱼:全长为(3 266±63) μm,与6 日龄相比,躯干部分黑色素细胞沉积大量增加,在体表上形成了大块黑色色斑。虹膜环明显变宽,内缘呈灰白色,外缘呈黄绿色(图版Ⅳ-5)。背鳍位置向尾部后移,尾鳍逐渐呈扇形,尾鳍柄缩细,雏形开始体现 (图版Ⅲ-2、图版Ⅳ-8)。
12 dph 仔鱼:全长为(3 849±65) μm,背鳍和臀鳍区域开始出现明显的色素沉积,尾柄处变得更加纤细,肠道出现第2 个生理弯曲(图版Ⅳ-14),除头下半部和尾端外,整个身体上遍布有分散的棕黄色色斑,整体颜色为深褐色 (图版Ⅲ-3)。
15 dph 仔鱼:全长为(5 016±88) μm,除了尾端外,其他区域可见大量黑色素遍布鱼体表面,体色以黄黑为主。虹膜环占眼球面积的2/3,均匀分布大量的颗粒状色素(图版Ⅳ-6)。尾柄部位愈加明确,脊索弯曲(图版Ⅳ-9),各鳍面逐步完善,尾鳍、第一背鳍、第二背鳍及臀鳍出现 (图版Ⅲ-4)。
20 dph 仔鱼:全长为(10 743±127) μm,除尾部和眼囊,全身大量色素沉淀,不透明,脊索末端弯曲完成。腹鳍出现,第一背鳍鳍条6 根;第二背鳍鳍条18 根;臀鳍鳍条24 根;尾鳍扇形面积增大,尾鳍鳍条34 根,尾叉出现(图版Ⅲ-5、图版Ⅳ-10)。

图版 Ⅳ 黄鹂无齿鲹各部位发育形态特征1~6. 眼睛发育,7~10. 尾鳍发育,11~14. 肠道发育。Plate Ⅳ Morphological characteristics of various parts of G. speciosus in development 1-6. eye development, 7-10. caudal fin development, 11-14. intestine development.
3 讨论
3.1 黄鹂无齿鲹的胚胎发育特征
鱼类胚胎发育可大致分为8 个阶段,包括受精卵期、胚盘期、卵裂期、囊胚期、原肠胚期、神经胚期、器官形成期和孵化出膜期。但不同鱼种在各发育阶段的命名和过程描述上存在一定差异。如毕加索小丑鱼(Amphriprion percula)、云南光唇鱼(Acrossocheilus yunanensis)和美洲鲥(Alosa sapidissima)的胚胎发育较为典型,可明确区分出8 个发育阶段[10-12]。而锦鲤(Cyprinus carp)、棘头梅童鱼(Collichthys lucidus)、金钱鱼(Scatophagus argus) 、翘嘴鲌(Culter alburnus)等[13-17]则缺失了胚盘期的描述,这可能是由于胚盘期时间较短导致的。另外,云南盘鮈(Discogobio yunnanensis)和台湾泥鳅(Paramisgurnus dabryanus)亦未详细描述胚盘期和孵化期的特点,仅描述了6 个发育阶段[18-19]。此外,以斑马鱼(Danio rerio)为例,其发育阶段的命名存在个别差异,如增加了体节期和眼囊期等[20]。这些差异一方面反映了鱼类胚胎发育过程中存在一定的物种特异性,但总体发育规律是高度一致的。研究发现,发育时间长短更多的与鱼卵的黏性程度相关,黏性卵发育时间较长,非黏性卵较短,这一点也不完全对应于淡水鱼和海水鱼的区分[10,21-28]。因此,尽管存在细微差异,鱼类胚胎发育的整体发育顺序和过程是高度同质的。后续研究需关注这些差异形成的内在机制,以期对不同物种发育规律有更深入理解。
黄鹂无齿鲹为广盐性鱼类,其受精卵仅能在海水条件下正常发育。通过室内人工繁育观察发现,黄鹂无齿鲹胚胎发育可分为8 个阶段:受精卵期、胚盘期、卵裂期、囊胚期、原肠胚期、神经胚期、器官分化形成期和出膜期。这一发育模式与其他硬骨海水鱼类如军曹鱼(Rachycentron canadum)[29]、黄 斑 篮 子 鱼(Siganus oramin)[30]、豹纹鳃棘鲈(Plectropomus leopardus)[31]等高度相似。值得一提的是,尽管上述研究均对胚胎发育中胚盘隆起阶段进行了记录与描述,但其对该阶段的命名仍存在差异,如军曹鱼研究中,其将受精卵发育至胚盘隆起阶段合称为合子期,而在对豹纹鳃棘鲈的研究中,则将胚盘隆起阶段归属于卵裂期等。这可能是由于鱼类胚胎发育胚盘期时间较短引起的命名差异,实际上,鱼类从受精卵期发育至胚盘形成阶段经历了细胞质及细胞核从植物极流向动物极并最终形成盘状胚盘的过程,是鱼类胚胎发育中首个细胞的形成过程。因此,本研究将胚盘期作为黄鹂无齿鲹发育过程中独立的一个时期进行描述。黄鹂无齿鲹胚胎发育所需时间为18.5 h,其发育模式及周期均类似于豹纹鳃棘鲈的胚胎发育,属于典型的浮性卵类型。黄鹂无齿鲹胚胎出膜后仍无法直接摄食,约需72 h 才能开始获取外源性营养。同时出膜个体的游泳能力较弱,通常漂浮于水体上层。待鳔完全形成(约96 h)后才能自由控制体位。
3.2 胚胎发育的速率及积温
鱼卵的大小对其发育进程有重要影响,一般以卵径表示其大小。殷名称[32]的研究表明,鱼卵大小对其生长发育有重要生态意义,总体来看,卵径越大,发育速率越慢。以黄鹂无齿鲹为例,其卵径较小,所需有效积温为470.8~503.8 °C·h,而在相近条件下统计了2 种鲹科鱼类、4 种石斑鱼类,包 括 卵 形 鲳 鲹[30]、高 体 鰤(Seriola dumerili)[33-34]、棕点石斑鱼(Epinephelus fuscoguttatus)[35]、斜带石斑鱼(E.coioides)[36]、鞍带石斑鱼(E.lanceolatus)[37]及豹纹鳃棘鲈的卵径、发育温度、胚胎发育所需时间及有效积温(表2)。可见黄鹂无齿鲹卵径较卵形鲳鲹、高体鰤更小,其胚胎发育速率明显更快,在相近的温度条件下(22.0~26.0 °C),高体鰤胚胎经35 h 10 min 后孵化出膜[33]。而卵形鲳鲹的胚胎发育所需时间更是达到41 h 27 min[38]。而与礁栖性的石斑鱼属相比,在卵径、温度相近的条件下,斜带石斑鱼孵化时间要比黄鹂无齿鲹更长,但其他鲹科鱼类如卵形鲳鲹、高体鰤的胚胎发育所需时间则远远高于石斑鱼属鱼类,这可能与鮨科(Serranidae)鱼类和鲹科鱼类的种属特异性有关。

表2 7 种海水鱼类受精卵的直径、胚胎发育过程的温度与发育所需时间Tab. 2 Fertilized egg diameter, temperature and time needed of embryonic development in seven fish species
黄鹂无齿鲹与4 种鲹科鱼类相比较,其前中期生长速率较快,黄鹂无齿鲹卵径小,使得卵裂期、囊胚期、原肠期以及神经胚期等时期的发育时间相对缩短,从而整个胚胎发育的速率相对加快,各鲹科鱼类神经胚在胚胎发育过程中形成的时间见表3。

表3 5 种鲹科鱼类的神经胚形成时间Tab. 3 Formation time of neurula of five species in Carangidae
3.3 黄鹂无齿鲹的仔鱼生长特征
在传统的鱼类早期生长阶段理论中,划分不同发育阶段的方式尚未统一,其划分依据包括卵黄囊的消耗殆尽、由内源性营养转向外源性营养输入、脊索弯曲等。本研究依据卵黄囊的消失,将仔鱼期划分为前仔鱼期(0~3 dph)和后仔鱼期(4~20 dph),并根据脊索的完全弯曲、各鳍条的出现判断后仔鱼期发育是否结束。黄鹂无齿鲹仔鱼初孵时平均全长 (1 520±19) μm,其卵黄囊3 dph即被消耗殆尽,在相近条件下,较同为鲹科鱼类的高体鰤发育时间明显缩短[33],黄鹂无齿鲹仔鱼发育进入后仔鱼期;同属内源性营养供给的油球于6 dph 后被仔鱼完全吸收。15 dph,脊索开始向上弯曲,各鳍逐渐成型。20 dph,脊索完全弯曲,各鳍出现且鳍条完备,后仔鱼期发育结束。
本研究就相近条件下,对比了5 种鲹科鱼类[33-34,39-41]的仔鱼阶段发育特征(表4),发现在4种鲹科鱼类中,黄鹂无齿鲹初孵仔鱼体型最小,但其仔鱼期的发育速率较其他同为鲹科的鱼类快,20 dph 时即完成了仔鱼期的发育,这可能与其独特的摄食方式有关,有研究者提出观点,初孵仔鱼的体型越大,其口裂也越大,其开口摄入饵料的时间也越早。黄鹂无齿鲹吻部长而宽大,没有咽齿,仅依赖可伸缩的吻部吸取食物进行摄食,因此尽管黄鹂无齿鲹初孵仔鱼体型较小,但其在开口摄食阶段仍然可获得相对更多的养分,促进鱼体各部位的分化发育。黄鹂无齿鲹鱼苗的早期生长呈现出生长速率缓慢的前期和快速增长的后期这种典型的 “异速生长” 模式,在0~9 dph 的平均生长速率仅为194 μm/d,但在10~20 dph 的平均生长速率达到680 μm/d。这一发育规律与某些其他鲹科鱼类的观察结果一致[33]。这种两段式生长策略被认为是鱼类在进化过程中适应自然生态环境的重要方式之一,可提高脆弱苗种在早期关键阶段的适应性和存活率。9 dph 前黄鹂无齿鲹仔鱼体表色素以点状和散生的枝状黄褐黑色素为主,但随后体色迅速加深,10 dph 起以大量黑黄色块为主要特征,颜色分布趋于均匀,整体呈现黄黑色调。这提示了体表色素沉积量和模式均与个体生长存在高度协调性。
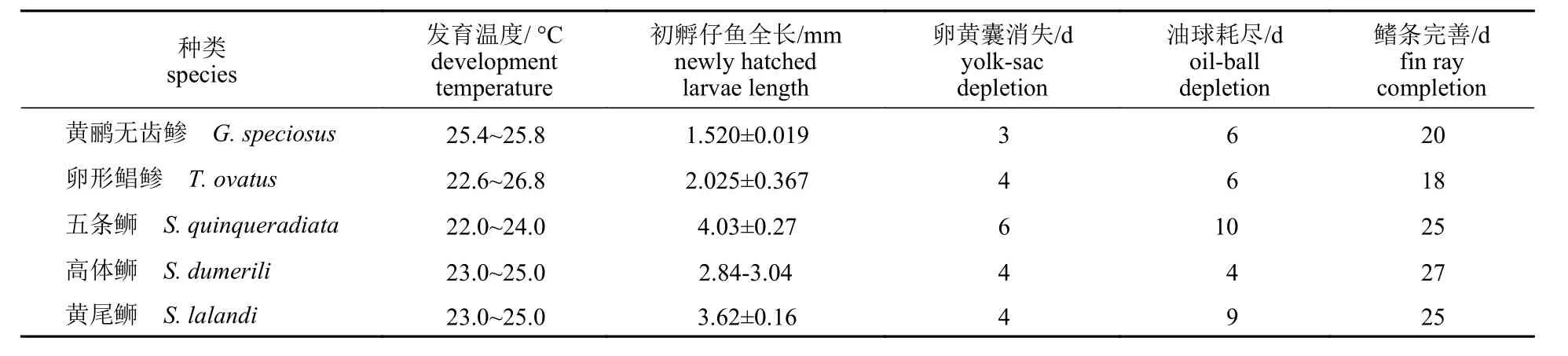
表4 5 种鲹科鱼类仔鱼阶段发育特征Tab. 4 Main characteristics of larvae stage of five species in Carangidae
本研究发现黄鹂无齿鲹与卵形鲳鲹的胚胎及胚后发育特征比较接近。卵形鲳鲹是我国广东、海南海洋牧场的主养品种,黄鹂无齿鲹将来的开发和产业发展,可以借鉴卵形鲳鲹成熟产业链关键环节的技术和管理。
(作者声明本文无实际或潜在的利益冲突)
猜你喜欢
小学生优秀作文(低年级)(2024年3期)2024-04-07
科普童话·百科探秘(2023年5期)2023-06-19
渔业研究(2022年5期)2022-10-28
北京支部生活(2022年4期)2022-05-25
雪豆月读·中年级(2021年12期)2021-01-03
奥秘(2018年9期)2018-09-25
浙江理工大学学报(自然科学版)(2016年5期)2016-09-15
幼儿100(2016年30期)2016-02-28
广东海洋大学学报(2015年4期)2016-01-13
集美大学学报(自然科学版)(2015年4期)2015-02-28
