
外源GABA对碱性盐胁迫下猴樟幼苗呼吸代谢相关酶活性的影响
2024-05-16 15:25任莉雯杨力行魏李彤周德玥周仪容韩浩章
安徽农学通报 2024年8期
任莉雯 杨力行 魏李彤 周德玥 周仪容 韩浩章


摘要 碱性盐胁迫影响植物的正常生长发育,γ-氨基丁酸(GABA)能有效缓解盐碱逆境对植物的伤害。本研究以猴樟幼苗为材料,共设5个处理,分别喷施0(对照)、30、50、100 和150 ?mol/L的GABA溶液,选择琥珀酸脱氢酶(SDH)、丙酮酸脱氢酶(PDH)、苹果酸脱氢酶(MDH)、柠檬酸合成酶(CS)和异柠檬酸合成酶(ICDH)指標,研究外源GABA处理对猴樟幼苗呼吸代谢相关酶活性的影响,为植物响应碱性盐胁迫的机理研究提供依据。结果表明,外源喷施30~50 ?mol/L浓度的GABA处理能明显提高碱性盐胁迫下猴樟幼苗叶片的SDH、PDH、MDH、CS和ICDH活性;100~150 ?mol/L浓度的GABA处理对呼吸代谢相关酶活性的促进作用不明显;随着外源GABA处理浓度的增加,猴樟幼苗的耐碱性盐胁迫能力综合评价值D呈先增加后降低的趋势,综合评价值D在30~50 ?mol/L浓度的GABA处理条件下达最大值,而100~150 ?mol/L浓度的GABA处理条件下D值低于对照。因此,30~50 ?mol/L浓度的GABA处理能促进碱性盐胁迫下的猴樟幼苗呼吸代谢能力。
关键词 GABA;碱性盐胁迫;呼吸代谢;猴樟;PDH活性
中图分类号 S792.23 文献标识码 A
文章编号 1007-7731(2024)08-0056-05
土壤盐碱化影响作物正常生长发育,最终影响作物产量和产品品质提升。碱性盐环境通常会造成渗透胁迫、氧化胁迫和离子毒害等,使植物光合作用受阻,养分吸收困难,甚至生长停止,最终导致植物组织早衰或死亡[1]。植物历经长期进化过程,具备了较复杂的碱性盐逆境应答系统。植物对盐碱地的逆境响应涉及多种生物化学和生理生化机制的调控,包括调节渗透压、积累渗透保护物质、调节离子吸收分配和转移、调节叶绿素组成以及维持细胞内膜脂状态等[2]。
猴樟(Cinnamomun bodinieri Levl.)为樟科樟属常绿阔叶树种,具有重要的药用价值、林用价值和油料加工价值,多分布于南方地区,在生产栽培过程中易受盐碱环境影响,可能导致其生长不良、抗性下降。秦嗣军等[3]发现,植物在应对非生物胁迫时,能量代谢路径会发生适应性变化,如水分胁迫使东北山樱幼苗通过调整磷酸戊糖(PPP)途径和交替呼吸(AP)途径比例的方式提高抗性。碱性盐胁迫下,猴樟组织内己糖激酶、乙醇脱氢酶、磷酸甘油酸激酶和磷酸甘油酸变位酶基因上调表达,草酰乙酸含量上升。植株呼吸代谢情况与其是否耐受碱性盐胁迫密切相关[4]。盐胁迫条件下,耐盐的燕麦品种产量明显高于盐敏感燕麦品种,在盐胁迫下盐敏感燕麦品种根系呼吸速率下降明显,抗氧化能力降低,营养供应不足,限制了根系生长及干物质的积累,最终导致产量下降[5]。
GABA为四碳非蛋白质类氨基酸,是植物体内重要的信号传导分子,参与种子萌发、氨基酸代谢、渗透调节、光合作用和活性氧调节等多种生理生化过程[6]。盐胁迫会抑制植物生长发育,呼吸代谢能为植物适应盐碱胁迫提供能量和小分子代谢物质,多种外源物质处理均能影响逆境胁迫下植物的呼吸代谢[5,7-8],碱低温驯化使高山植物绵头雪莲呼吸速率和糖酵解速率先降低后升高,抗氰呼吸途径加强,与糖代谢相关的关键酶基因表达量提高[9]。研究表明,外源GABA能够提高盐胁迫下菘蓝幼苗活性氧清除能力和抗氧化酶的活性[10],为植物幼苗生长发育提供氮素营养,促进叶绿素的合成,诱导产生细胞内多种氨基酸[11]。外源GABA也能够提高盐胁迫下西伯利亚白刺的光合能力[12],提高植物的抗逆能力,缓解非生物胁迫对植物的损害[13]。外源水杨酸(SA)处理可有效调控冬小麦糖酵解(EMP)代谢,促进果糖与丙酮酸积累与分解,提高植物的抗寒性[14]。赤霉素(GA3)处理提高了低温胁迫条件下玉米种胚中可溶性糖和可溶性蛋白含量,提高了淀粉酶和呼吸途径关键酶活性,提高了MDH、丙酮酸激酶(PK)、联合酶(G-6-PDH和6-PGDH)的活性,提高了EMP、三羟酸循环(TCA)、PPP途径的运转效率,提高了种子对低温的抵抗能力,实现在低温胁迫条件下种子的萌发[12]。植物体内的GABA由多胺代谢途径或谷氨酸脱羧途径合成,经由GABA转氨酶催化后,最终进入TCA代谢[15]。
糖是植物光合作用的主要产物,也是呼吸作用的底物,为植物细胞的构建和耗能提供碳骨架和能源,同时具有调节渗透、抵抗环境胁迫和影响生长发育等重要作用。在盐碱、低温和干旱等非生物胁迫下,植物体内的可溶性糖的含量会提高,而可溶性糖含量是植物抵抗非生物胁迫能力的重要体现。EMP、TCA、SDH、PDH、MDH、CS和ICDH是植物呼吸代谢过程中的关键酶。目前,关于GABA对植物碱性盐胁迫下呼吸代谢影响的研究较少,本研究基于前期研究结果,探索外源GABA处理对猴樟幼苗呼吸代谢相关酶活性的影响,为植物耐碱性盐胁迫的分子机理研究提供参考。
1 材料与方法
1.1 试验材料
试验材料选自宿迁学院樟属植物资源圃生长的1年生盆栽猴樟幼苗,2022年9月从同一母树上采种,低温层积后于2023年2月播种,2023年5月移栽至盆中,试验材料共60盆,每盆1株。栽培盆高22 cm,栽培基质比例为基地田园土∶草炭土=3∶1,土壤pH值为7,在玻璃温室中进行培养,进行正常水肥管理。
1.2 试验设计
试验共分5个处理,每处理有12盆猴樟幼苗。以前期研究为基础,2023年10月12日9:00用200 mmol/L Na2CO3溶液进行浇灌处理,用量为800 mL/盆;当日15:00,外源喷施浓度分别为0(对照)、30、50、100和150 ?mol/L的GABA溶液,用量为100 mL/盆,喷至叶面滴水为止,剩余溶液浇施于盆内,1 h后再喷施1次,用量为100 mL/盆。正常培养48 h后,检测猴樟幼苗叶片的SDH、PDH、MDH、CS和ICDH活性,进行3次生物学重复。
1.3 试验方法
采用SDH活性检测试剂盒(BC0955)测定SDH活性,每1 g植物组织在反应体系中每1 min消耗1 nmol 2,6-二氯酚靛酚定义为1个酶活力单位U;采用分光光度计法检测PDH活性,以每1 min吸光度值升高0.01为1个酶活力单位U;采用分光光度计法检测MDH活性,以每1 min吸光度值升高0.01为1个酶活力单位U;采用分光光度计法检测CS活性,以每1 min吸光度值升高0.01为1个酶活力单位U;采用分光光度计法检测ICDH活性,以每1 min吸光度值升高0.01为1个酶活力单位U[7]。外源GABA处理对碱性盐胁迫下猴樟幼苗呼吸代谢影响效果的综合评价采用隶属函数法[16],在所有测定指标中,SDH、PDH、MDH、CS和ICDH活性均与抗碱性盐胁迫能力成正相关,采用隶属函数公式(1)进行计算。
[U(Xab)=(Xab-Xamin)/(Xamax-Xamin)] (1)
式(1)中,[U(Xab)]为测定指标的隶属函数值,Xab为各处理的指标测定值,Xamin为各处理指标测定的最小值,Xamax为各处理指标测定的最大值。
1.4 数据统计与分析
采用Microsoft Excel 2019制表,用SPSS 21.0进行数据统计,用最小显著差数法(LSD)分析数据之间的差异性。
2 结果与分析
2.1 外源GABA处理对碱性盐胁迫下猴樟幼苗PDH活性的影响
PDH是分布在线粒体基质中由3个酶组合而成的复合体,负责催化丙酮酸脱羧,可生成乙酰辅酶A。使用不同浓度的外源GABA处理影响碱性盐胁迫下猴樟幼苗PDH活性(图1),其中30、50 ?mol/L浓度的外源GABA处理下碱性盐胁迫下PDH活性与对照组的差异有统计学意义(P<0.05),分别较对照提高了32.12%和77.58%;100、150 ?mol/L的外源GABA处理对PDH活性影响与对照组的差异无统计学意义(P>0.05)。因此,30~50 ?mol/L浓度的外源GABA处理对PDH活性的影响效果较好。
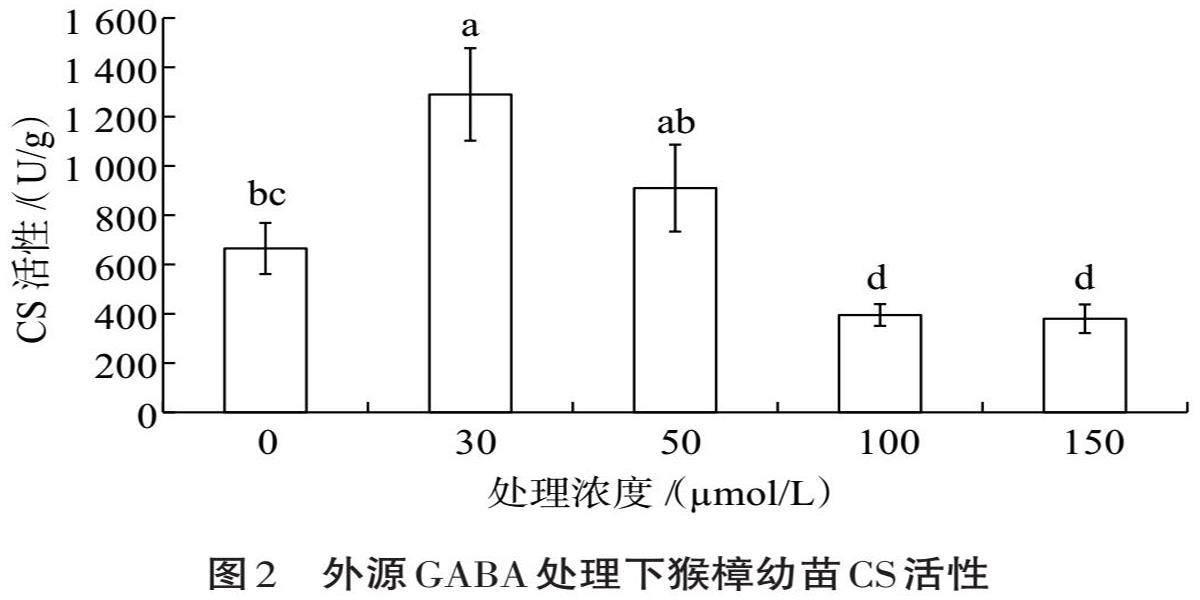
2.2 外源GABA处理对碱性盐胁迫下猴樟幼苗CS活性的影响
在三羧酸循环第一步反应中,CS能催化乙酰辅酶A的乙基与草酰乙酸的酮基结合,生成柠檬酰辅酶A。适宜浓度的外源GABA处理影响了碱性盐胁迫下猴樟幼苗CS活性(图2),随着GABA处理浓度的提高,CS活性呈先升高后下降的趋势;在30 ?mol/L时CS活性达最高值,与对照组差异有统计学意义(P<0.05),比对照组提高了94%,随后CS活性明显下降,其中50 ?mol/L外源GABA处理较对照提高了36.8%,但二者间差异无统计学意义(P>0.05),而100和150 ?mol/L外源GABA處理对CS活性影响低于对照,与对照组差异无统计学意义(P>0.05)。因此,30~50 ?mol/L的外源GABA浓度处理对CS活性的影响效果较好。
2.3 外源GABA处理对碱性盐胁迫下猴樟幼苗ICDH活性的影响
ICDH催化异柠檬酸转化成α-酮戊二酸,实现了TCA循环中第一次脱羧过程。适宜浓度的外源GABA处理影响了碱性盐胁迫下猴樟幼苗ICDH活性(图3),其中30 ?mol/L的外源GABA处理明显提高碱性盐胁迫下ICDH活性,与对照组差异有统计学意义(P<0.05),较对照提高了163.55%;随着GABA处理浓度的提高,ICDH活性迅速下降,其中50、100 ?mol/L浓度的GABA处理下ICDH活性低于对照,但二者间差异无统计学意义(P>0.05),150 ?mol/L外源GABA浓度处理时的ICDH活性较对照提高了15.89%,但二者间差异无统计学意义(P>0.05)。因此,30 ?mol/L浓度的外源GABA处理对ICDH活性的影响效果较好。
2.4 外源GABA处理对碱性盐胁迫下猴樟幼苗SDH活性的影响
SDH是TCA循环和线粒体电子传递链的关键酶,在细胞能量代谢中起重要作用,可以根据其活性来推测TCA循环的活跃程度。适宜浓度的外源GABA处理影响了碱性盐胁迫下猴樟幼苗SDH活性(图4),随着外源GABA处理浓度的提高,SDH活性呈先升高再下降趋势,其中50 ?mol/L外源GABA处理明显提高碱性盐胁迫下SDH活性,与对照组差异有统计学意义(P<0.05),比对照提高了61.79%,而100和150 ?mol/L外源GABA浓度处理下的SDH活性低于对照,但二者间差异无统计学意义(P>0.05)。因此,50 ?mol/L浓度的外源GABA处理对SDH活性的影响效果较好。
2.5 外源GABA处理对碱性盐胁迫下猴樟幼苗MDH活性的影响
MDH主要将呼吸代谢中的苹果酸脱氢形成草酰乙酸,草酰乙酸可继续作为受体参与TCA循环。适宜浓度的外源GABA处理影响了碱性盐胁迫下猴樟幼苗MDH活性(图5),随着外源GABA处理浓度的提高,MDH活性呈先升高再下降趋势,但与对照组的差异均有统计学意义(P<0.05),分别较对照提高了104.35%、143.04%、92.61%和70.43%。因此,30~150 ?mol/L浓度的外源GABA处理对MDH活性的影响效果均较好。
2.6 外源GABA处理对碱性盐胁迫下猴樟幼苗呼吸代谢影响效果的综合评价
综合评价值D表示不同浓度外源GABA处理对碱性盐胁迫下猴樟幼苗呼吸代谢影响的综合效果评价,D值范围为0~1,D值越接近1表示外源GABA处理对碱性盐胁迫下猴樟幼苗呼吸代谢的影响效果越好,植物耐碱性盐胁迫能力越强,反之则效果越差[17]。由图6可以看出,不同浓度外源GABA处理对碱性盐胁迫下猴樟幼苗呼吸代谢影响的综合效果不一致,其中30~50 ?mol/L浓度的外源GABA处理效果较好,D值分别为0.806和0.633,分别较对照提高了285.94%和203.33%,而100~150 ?mol/L浓度的外源GABA处理效果较差,D值分别为0.142和0.102,分别较对照降低了31.98%和51.21%。
3 结论与讨论
碱性盐胁迫影响植物的正常生长发育,γ-氨基丁酸(GABA)能有效缓解盐碱逆境对植物的伤害,本研究以猴樟幼苗为材料,研究外源GABA处理对猴樟幼苗呼吸代谢相关酶活性的影响,为植物响应碱性盐胁迫的机理研究提供参考。
本研究结果表明,呼吸代谢在植物响应碱性盐过程中发挥重要作用,包括供应能量、合成中间产物等多个方面,抗碱性盐胁迫能力强的植物品种的呼吸代谢越旺盛。外源化学药剂处理参与并促进植物呼吸代谢过程,适宜浓度的GABA处理均能促进猴樟幼苗适应碱性盐胁迫,提高种苗抗性和生长质量,但不同浓度GABA处理的影响效果不一致。结合隶属函数法的结果来看,30 ?g/L外源GABA和50 ?g/L外源GABA处理效果最好,D值分别为0.806和0.633,而100~150 ?mol/L外源GABA浓度处理效果较差,D值分别为0.142和0.102。因此,在猴樟幼苗期叶面喷施30~50 ?mol/L外源GABA可明显提高猴樟幼苗的抗盐碱能力。
在本试验中,叶面喷施适宜浓度的GABA可明显提高猴樟幼苗在碱性盐胁迫下的SDH、PDH、MDH、CS和ICDH活性,提高植物在碱性盐胁迫下的能量供应能力和草酰乙酸等小分子物质含量,提高植物抗逆性,这与吴繁琦等[18]对甘草、贾璐婷[19]对东北山樱的研究结果一致。
参考文献
[1] TAROLLI P,LUO J,PARK E,et al. Soil salinization in agriculture:mitigation and adaptation strategies combining nature-based solutions and bioengineering[J]. iScience,2024,27(2):108830.
[2] LIAO J M,XIA P G. Continuous cropping obstacles of medicinal plants:focus on the plant-soil-microbe interaction system in the rhizosphere[J]. Scientia horticulturae,2024,328:112927.
[3] 秦嗣军,吕德国,李志霞,等. 水分胁迫对东北山樱幼苗呼吸等生理代谢的影响[J]. 中国农业科学,2011,44(1):201-209.
[4] HAN H Z,ZHANG L H,LI S H,et al. Transcriptome and metabolome integrated analysis reveals the mechanism of Cinnamomum bodinieri root response to alkali stress[J]. Plant molecular biology reporter,2023,41(3):470-488.
[5] 张碧茹,米俊珍,赵宝平,等. 外源γ-氨基丁酸缓解燕麦幼苗盐碱胁迫的生理效应[J]. 麦类作物学报,2024,44(2):222-229.
[6] 韩多红,王恩军,张勇,等. 外源γ-氨基丁酸对盐胁迫下菘蓝幼苗活性氧和抗氧化系统的影响[J]. 北方园艺,2021(6):111-116.
[7] 贾邱颖,吴晓蕾,冀胜鑫,等. γ-氨基丁酸对番茄嫁接苗耐盐性的生理调控效应[J]. 植物营养与肥料学报,2021,27(1):122-134.
[8] 王馨,闫永庆,殷媛,等. 外源γ-氨基丁酸(GABA)对盐胁迫下西伯利亚白刺光合特性的影响[J]. 江苏农业学报,2019,35(5):1032-1039.
[9] 常菲菲,杨志杰,冉春霞,等. 外源γ-氨基丁酸缓解番茄幼苗热胁迫的生理生化机制[J]. 植物生理學报,2023,59(11):2039-2051.
现代农业科技,2023(24):88-91.
[10] 朱育贤,施诗,岂子雁,等. 褪黑素对碱性盐胁迫下猴樟呼吸代谢相关酶活性的影响[J]. 现代农业科技,2023(24):88-91.
[11] 罗立娜,李绍鹏,吴凡,等. 隶属函数法筛选多效唑对香蕉除芽促长效果的最佳剂量[J]. 分子植物育种,2018,16(12):4079-4085.
[12] 王青青,胡艳丽,周慧,等. 根皮苷对平邑甜茶根系TCA循环酶的影响[J]. 中国农业科学,2012,45(15):3108-3114.
[13] 陈晓晶,徐忠山,赵宝平,等. 盐胁迫对燕麦根系呼吸代谢、抗氧化酶活性及产量的影响[J]. 生态学杂志,2021,40(9):2773-2782.
[14] 陆雅宁,王晓静,杨佳慧,等. 低温驯化对绵头雪莲呼吸代谢及相关基因表达的影响[J]. 分子植物育种,2023,21(10):3171-3179.
[15] 赵欣,孟德义,苍晶,等. 外源SA对低温胁迫下冬小麦糖酵解代谢的影响[J]. 麦类作物学报,2022,42(8):1031-1038.
[16] 任晓松,王子沐,焦健,等. GA3处理对低温胁迫条件下玉米种子呼吸代谢的影响[J]. 生态学杂志,2020,39(3):847-854.
[17] SHELP B J,BOZZO G G,TROBACHER C P,et al. Hypothesis/review:contribution of putrescine to 4-aminobutyrate (GABA) production in response to abiotic stress[J]. Plant science:an international journal of experimental plant biology,2012,193/194:130-135.
[18] 吴繁琦,王桂荣,白钰,等. γ-氨基丁酸对甘草种子萌发及呼吸代谢的影响[J]. 种子,2023,42(4):18-24.
[19] 贾璐婷. 水涝胁迫下东北山樱根系GABA代谢特征及其生理效应研究[D]. 沈阳:沈阳农业大学,2018.
(责编:张 蓓)
基金项目 江苏省自然科学基金项目(BK20231362);江苏省高等学校大学生创新创业训练计划项目(2023XSJ144Y);宿迁市自然科学基金项目(K202316);宿迁学院创新团队项目(2021td05)。
作者简介 任莉雯(2003—),女,江苏扬州人,从事樟属植物逆境分子生物学研究。
通信作者 韩浩章(1978—),男,山东青岛人,硕士,副教授,从事樟属植物逆境分子生物学研究。
收稿日期 2024-02-15
