
枸杞岛贻贝筏式养殖区附着藻类群落结构及其生物固碳量的估算
2024-05-13 08:35:02苏小艳郭浩宇吴嘉豪林觉润吴浩博张秀梅
浙江海洋大学学报(自然科学版) 2024年1期
田 阔,苏小艳,胡 扬,李 启,郭浩宇,吴嘉豪,周 倩,洪 晨,林觉润,吴浩博,张秀梅
(浙江海洋大学水产学院,浙江舟山 316022)
大型海藻具有净化海洋环境,提供初级生产物,维持海洋生物生命活动等重要作用,由其群落支撑构建的海藻场是维持岛礁生态系统稳定和生物多样性的重要生境[1]。因大型藻类生长固着的岩相基质不具有碳储藏的功能,大型藻类生态系统一直被排除在“蓝碳”系统之外。近年,已经有越来越多的证据指明该生态系统可以通过碳迁移、形成惰性有机物颗粒(LROD)等方式储碳,具备巨大的碳汇潜力[2-3]。
海洋碳汇是指长久封存的碳,长达1 000 a 以上,使其不再参与大气循环[4]。大型海藻作为重要的海洋初级生产者,具有很高的固碳效率,开展大规模的藻类养殖是实现联合国可持续性发展目标(SDGs)的有效手段之一[5]。目前,普遍认可的藻类碳汇方式包括海藻本身的生物固碳、海藻碎屑的浅海沉积、埋藏或深海迁移等。在马来西亚半岛养殖的张氏江蓠Gracilaria changii 中总碳、总有机碳和总无机碳的含量较高,其在吸收环境中的CO2具很高的应用价值[6]。不同藻类的固碳能力亦有不同,浙南沿岸的11 种大型海藻中,褐藻类的固碳能力最强,如海带Laminaria japonica、鼠尾藻Sargassum thunbergii、铜藻Sargassum horneri,其次是红藻,而羊栖菜Hizikia fusiforme、石莼Ulva lactuca 等固碳能力较弱[7]。枸杞岛潮间带海藻场的优势固碳藻类为鼠尾藻、刚毛藻、羊栖菜[8]。另外,环境因素也会影响藻类固碳效率。针对獐子岛礁区大型藻类碳汇能力的研究发现,叶绿素a、水深、硝酸盐和溶解氧都能促进大型藻类固碳,温度则会影响其固碳速率[9]。硇洲岛良好的生态环境也能够促进大型海藻高效固碳[10]。高流速会提高鼠尾藻的净初级生产力,增加固碳能力等[11]。大型藻类除自身固碳外,还可以通过被牧食或者利用海藻叶片或藻床复杂的空间结构吸引其他物种附着聚集,形成间接的固碳效应,如钩虾会选择固碳效率更好的绿藻啃食,并通过不同营养级食物链,快速传递藻类已固定的碳,对海洋渔业碳汇产生一定的影响[12]。
现有的藻类固碳研究中,往往侧重于具有优良固碳效率藻种的选育,或是环境因子对藻类固碳效率的影响研究。而种类群落多样性(种间相互作用)对其固碳能力的影响等相关研究较少。并且,藻类固碳量的多少主要还是取决于其本身累积的生物量。生物量与生物多样性之间的关系一直是生态学界争论的焦点,生物量与生物多样性是生态系统功能的重要驱动力。植物群落中植物多样性和生物量之间存在“驼峰”关系,这种驼峰多样性—生物量关系,通常归因于多种种间互补过程。在低生物量水平下,生物量和多样性通常是正相关,而在高生物量水平下,少数物种控制着资源,物种间竞争排斥,生物量与多样性呈负相关。在植物群落中,这种驼峰模型已经得到较多的扩展,而在藻类群落的研究中多样性与生物量间的关系还尚不明确[13]。
枸杞岛贻贝筏式养殖区是一个易受人为影响的生境,筏架区内水流缓慢,营养盐丰富,这为藻类的生长创造了良好的环境,养殖筏架上附着的藻类种类丰富,生物量可观。但目前,(1)该系统中藻类群落的多样性与生物量之间的关系尚不清楚;(2)筏式养殖区内藻类的生物固碳量也无可参考数据,这影响了对该区域的藻类资源利用。因此,本研究通过对枸杞岛贻贝养殖筏架附着大型海藻进行了为期一年的多样性本底调查,计算各站位的藻类多样性及其生物量;结合已公开的藻类含碳量数值,初步估算了枸杞岛贻贝养殖筏架区附着藻类的总体生物固碳量;并基于线性回归模型,讨论了藻类群落多样性与其生物量间的关系。研究结果为贻贝养殖区内的大型藻类固碳增汇提供了参考依据。
1 材料与方法
1.1 调查区域与样品采集
2021 年8 月、11 月,2022 年1 月、5 月对枸杞岛贻贝养殖筏架上的大型藻类群落进行了本底调查,养殖筏架区内共设置了12 个站位(图1)。
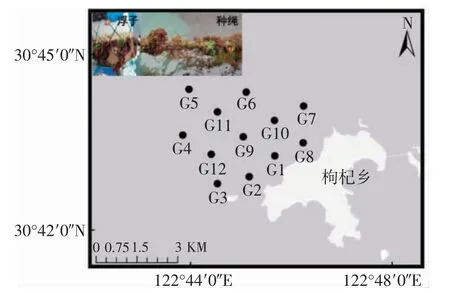
图1 采样站位示意图Fig.1 Diagrams of sampling stations
参考了先行研究的取样方法及《海洋调查规范》(GB/T 12763.6-2007)[14-17]。课题组于每站位,各取相同规格泡沫浮子1 个和苗绳60 cm,采集浮子及苗绳上所有藻类,低温避光保存运回实验室。将采集的藻类分类鉴定后,记录个体湿重、编号、烘干(80 ℃,>48 h)至恒重,逐一测量并记录干重。
1.2 数据分析
1.2.1 藻类群落物种多样性、优势种
(1)本研究采用α 多样性对群落组成进行了香农—威纳指数(Shannon-Wiener index,H′)、辛普森指数(Simpson index,D)、物种均匀度指数(Pielou index,J′)计算,具体公式分别如下:
式中:S 为海藻种类总数;Pi为出现种i 的生物量在出现种总生物量中占的比例大小,%。
(2)优势种采用相对重要性指数(IRI)计算,具体公式如下:
式中:Wi为第i 种海藻的生物量,g·m-2;W 为所有海藻种类的总生物量,g·m-2;F 为某一种类i 的出现站位数占采样总站位数的百分比,%;通常优势种即为优势指数大于0.02 的生物种类[16]。
1.2.2 回归模型构建
线性回归对于变量间复杂的关系可以直观呈现,且易于理解,这是我们本次研究采用的原因。我们将单个浮子上和苗绳上的藻类群落生物量(总干重)log10 转化后,作为响应变量,将多样性指数和物种数作为预测变量,构建线性回归模型(R 语言ggplot2)。通过P 值评估了预测变量与响应变量的显著性,R2为模型解释率。
1.2.3 大型藻类的含碳量估算
藻类直接吸收CO2通过光合作用转化固定碳,其固碳量的评估方法参考张继红等的核算方法[18-25]。收获上岸的大型藻类,相当于从水体中直接移出了碳,藻的碳汇贡献由藻的产量以及藻体的含碳量进行评估。公式如下:
式中:W(C)为藻体含碳量,%;不同海区同种藻类的W(C)并无显著性差异,其中藻类含碳量参考了张继红、权伟、周毅等[18,21,25-26]的研究方法,本研究取藻类平均含碳量为27.4%。
2 结果
2.1 藻类群落生物量及其优势种
经鉴定浮子上附着藻类共3 门79 种,绿藻14 种,红藻59 种,褐藻6 种。藻类组成以红藻类较多,绿藻类次之,褐藻类较少;春季藻类最多,秋季最少。优势种单个浮子附着的藻类生物量(干重g/个)为,春季34.81~374.24 g(mean±SD 179.5±101.5),夏季12.63~187.84 g(46.9±46.8),秋季20.68~91.10 g(42.0±18.0),冬季21.37~302.01 g(74.2±73.1)。浮子上各季度均以红藻为优势种类,尤其是膜状的带形蜈蚣藻Grateloupia turuturu 和椭圆蜈蚣藻Grateloupia elliptica 为主,小型红藻有密毛沙菜Hypnea boergesenii、大齿心藻Serraticardia maxima 等,秋冬季的优势藻种还有刚毛藻、裂片石莼Ulva fasciata (绿藻)、网地藻Dictyota dichotoma(褐藻)(表1,图2)。
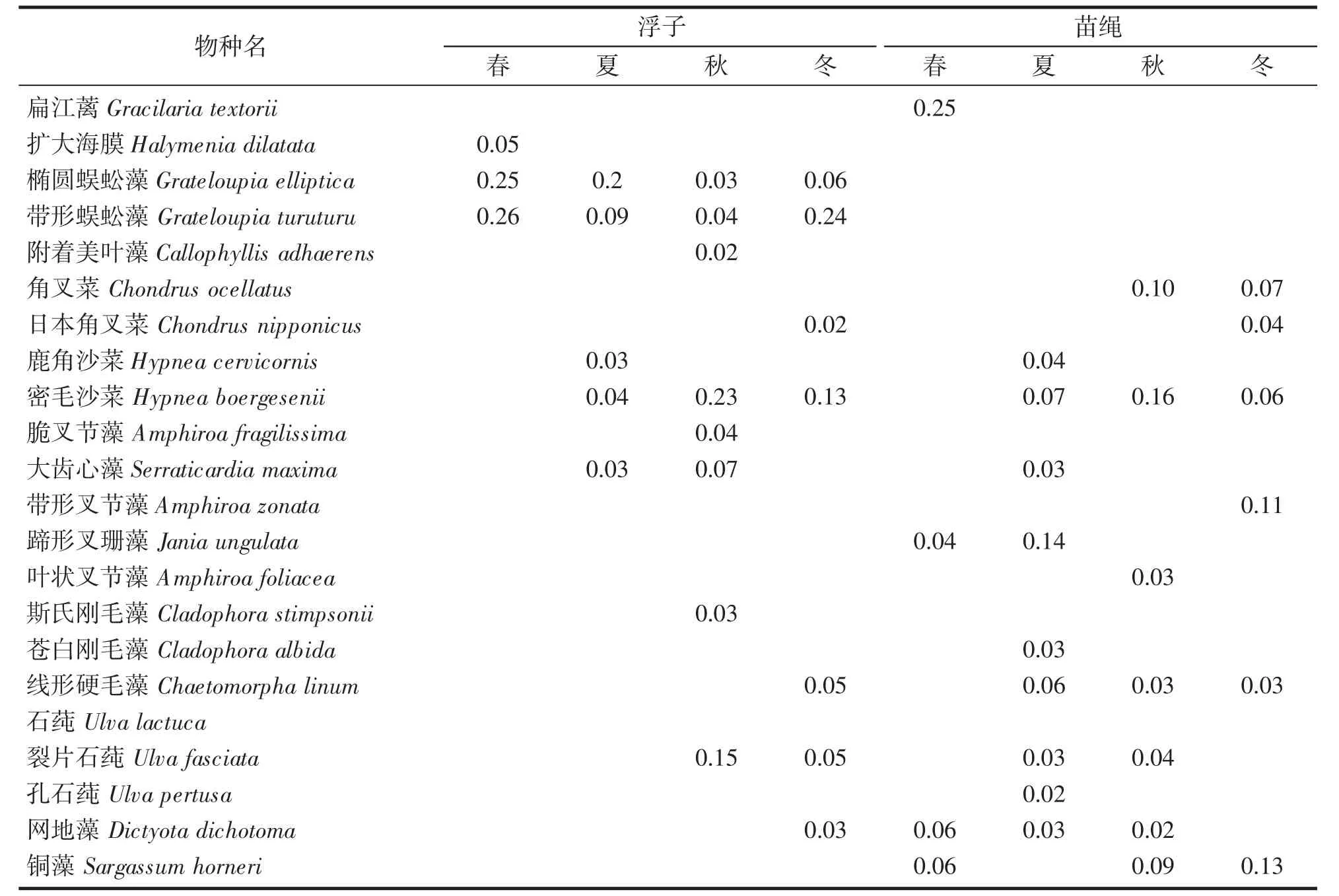
表1 浮子与苗绳上附着的优势藻种Tab.1 Dominant algae with a float attached to a seedling line

图2 贻贝养殖区的优势藻类Fig.2 Dominant algae in mussel culture areas
苗绳上附着藻类共3 门72 种,绿藻15 种,红藻55 种,褐藻4 种。春季最少,秋、冬季最多,主要以红藻为主;褐藻在秋冬季最多,春季最少,绿藻在春季种类有增加;苗绳附着的藻类生物量(干重g·m-1)为,春季28.43~121.67 g·m-1(mean±SD 77.3±29.8),夏季5.62~90.08 g·m-1(44.3±24.9),秋季2.05~84.07 g·m-1(31.9±22.1),冬季15.8~129.68 g·m-1(47.6±32.2)。苗绳上的优势种主要以密毛沙菜、蹄形叉珊藻Jania ungulata 和铜藻为主(表1,图2)。铜藻除夏季因繁殖脱落无法形成优势种以外,在其他季节都是优势种。线形硬毛藻Chaetomorpha linum、网地藻除个别季节外也是优势种。
2.2 藻类生物固碳量的估算
根据公式(5),计算了12 个站位浮子和苗绳上的藻类生物量,换算成单个浮子上附着的藻类生物量和苗绳上每米的藻类生物量。藻类群落整体含碳量为27.4%,估算了养殖筏架区的藻类生物固碳量。根据枸杞岛筏架的设计规格,浮绠长度为100 m,浮绠间距4.5 m,每行浮绠每一行挂80~120 个浮子[27]。枸杞岛现有贻贝养殖面积约1 000 hm2,按每450 m2,有浮子80~120 个,苗绳100 m 估算,则养殖区域现有浮子约1 778 664~2 667 996 个;苗绳长约2 223 333 m(1 000 m2/450 m2)。浮子上藻类群落的生物固碳量为春季87.5~131.2 t,夏季22.8~34.3 t,秋季20.5~30.7 t,冬季36.2~54.2 t。苗绳上的藻类的生物固碳量为春季47.1 t,夏季27 t,秋季19.4 t,冬季29 t。
2.3 藻类群落多样性
浮子藻类群落的各多样性指数的平均值冬季最高,秋季和夏季次之,春季最低(表2)。Shannon 指数波动范围0.46~2.40,最高值出现在秋季的G12 站位(2.40);Simpson 指数为0.22~0.87,最高值出现在冬季的G10 站位(0.87);Pielou 指数为0.22~0.78,最高值出现在冬季G6 站位(0.78)。
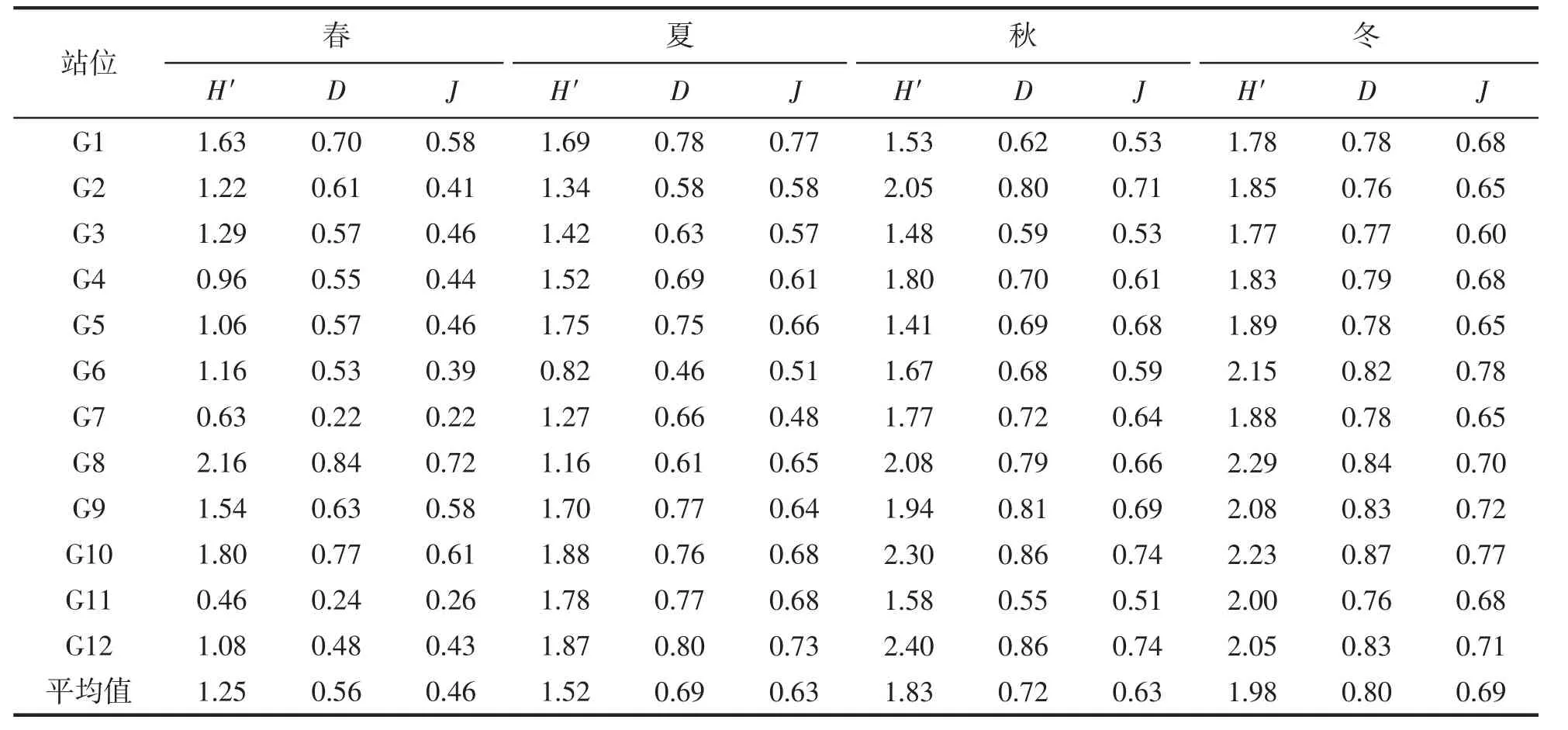
表2 各站位藻类群落的多样性指数(浮子)Tab.2 Diversity index of algae community at each station(float)
苗绳藻类群落的平均多样性指数在夏季最高,秋季和冬季次之,春季最低(表3)。春季G4 站位取样时,只有3 株铜藻,因此无法计算相关多样性指数。Shannon 指数波动范围0.35~2.33,最高值出现在夏季的G5站位(2.33);Simpson 指数为0.14~0.88,最高值出现在夏季的G5 站位(0.87);Pielou 指数为0.15~0.82,最高值出现在夏季G5 站位(0.82)。多样性指数最高值均出现在夏季的G5 站位,最低值均出现在春季的G10 站位。
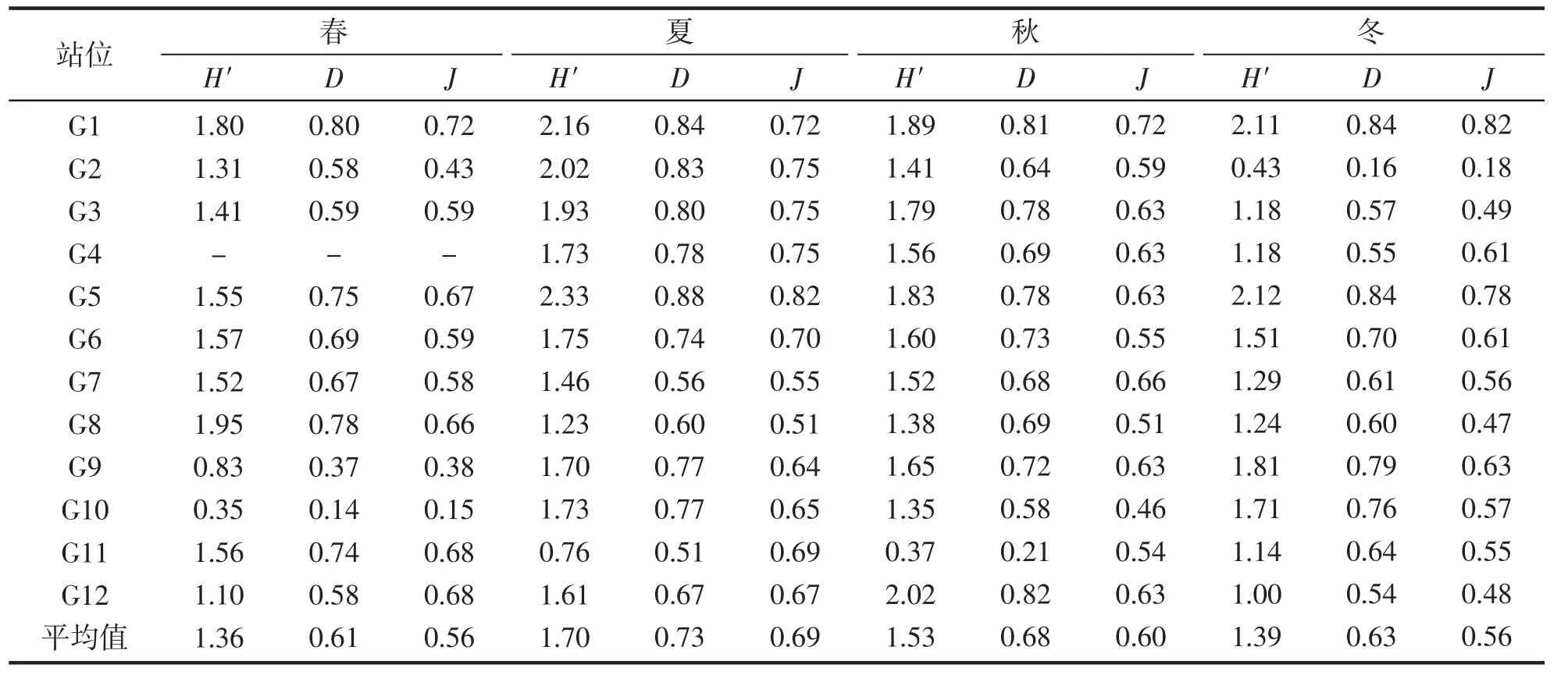
表3 各站位藻类群落的多样性指数(苗绳)Tab.3 Diversity index of algae community at each station(seedling rope)
2.4 群落多样性与生物量间的关系
浮子上藻类的多样性与生物量的关系见图3,春、秋、冬季藻类群落与生物量没有明显的线性关系。夏季,生物多样性与生物量呈显著相关(P<0.05,R2=0.267);均匀度指数与生物量呈极显著相关(P<0.01,R2=0.60)。
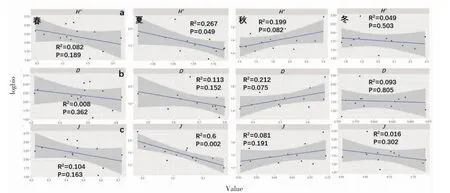
图3 藻类群落多样性与生物量的关系Fig.3 Relationship between algae community diversity index and biomass
苗绳上的藻类多样性与生物量关系见图4,春、夏、冬季藻类群落与生物量没有明显的线性关系。秋季,生物多样性与生物量呈显著正相关,多样性指数越高,生物量越高(P<0.01,R2=0.528)。辛普森指数与生物量也呈极显著正相关(P<0.01,R2=0.544)。冬季只有多样性指数与生物量呈显著负相关(P<0.05,R2=0.288)。
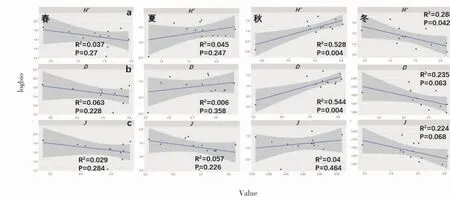
图4 藻类群落与生物量的关系Fig.4 Relationship between algae community diversity index and biomass
3 讨论
3.1 贻贝养殖区可移除碳量的估算
藻类生物固碳量的估算方法,采用的是生物量(干重)×含碳率,该方法是目前藻类固碳量研究中最常用的方法[18-25]。本文对采用同一固碳算法的,中国沿岸养殖藻类的每公顷固碳量数据进行了整理,山东省2016—2019 年养殖藻类的平均碳移除能力,海带约为34.99 t·hm-2、裙带菜37.39 t·hm-2、紫菜8.39 t·hm-2、江蓠为47.61 t·hm-2[23]。桑沟湾海草初级生产力的固碳贡献约为5 t·hm-2[28]。俚岛海带养殖固碳量为7 t·hm-2。莱州湾鼠尾藻海藻场的固碳量约为6 t·hm-2[29]。福州市开展的海带、紫菜、江蓠养殖藻类固碳量可达到14.82 t·hm-2[30]。广东省汕头南澳岛的龙须菜固碳量为6.7 t·hm-2[21]。本研究枸杞岛贻贝筏式养殖区的藻类春季最高固碳量约为0.18 t·hm-2。章守宇等[16]在2018—2019 年枸杞岛养殖筏架的附着藻类调查中,春季的藻类平均生物量35.08 t·hm-2,固碳量约1.89 t·hm-2(藻类干湿重比约为1/5)。这可能与年际间附着藻类的生物量差异有关。
养殖藻类采用单一栽培模式,每株植物都经过相同的标准栽培、维护和收获,养殖过程中的管理也趋于简单、高效,因此能以较低的成本获得更多的产量,这也是单一藻种养殖能获得较高生物量,单位面积内具有更大固碳潜力的原因。枸杞岛筏架养殖区的藻类群落多样性高,种质资源丰富,但没有开发出可单一栽培的藻种,藻类群落多以膜状、丝状、枝状红藻和珊瑚藻为优势种,这些藻多为一年生的小型藻类,会导致筏架区单位面积内生物量偏低,固碳潜力偏小。
3.2 多样性对生物量的影响
浮子的藻类群落多样性在秋冬季最高,3 个多样性指数的最高值分别出现在秋季、冬季的不同站位。这可能是浮子位于海水表层,更易受波浪、气温变动、人为因素(贻贝采收)等多方面因素影响,导致浮子上附着藻类的多样性指数分布呈现差异[16]。
夏季,浮子上附着藻类群落多样性与生物量呈显著负相关;其他季节无明显规律,物种数与生物量也无显著关系,同时该季节藻类的群落特征为生物量减少,多样性增加,这可能是群落演替所造成的,海膜类藻类消失降解,优势度降低,密毛沙菜等小型藻类优势度增加。另外,生物多样性与生物量的关系中,还存在着保险假说,物种多样性为环境变化提供了一种“保险”或缓冲,因为不同物种对这些变化的反应不同,从而导致更可预测的生态系统属性或功能[31]。在这个假设中,在某一特定时间对生物量具有功能冗余的物种随着时间的推移不再冗余[32]。因此,本研究中浮子上的附着藻类群落多样性与生物量之间在夏季呈负相关,也可能归因于物种冗余,其优势种均为红藻类。随着大型红藻类降解消失,冗余的小型红藻类增加,种间资源利用同质竞争激烈,导致生物量无法有效累积[12]。群落多样性越高,生物量反而越少,这可能也是自然附着藻类比单种养殖藻类固碳量低的原因之一。
苗绳上附着藻类群落多样性在夏季、秋季较高,多样性指数出现的最高值和最低值均为同季节、相同站位。藻类群落多样性与生物量在秋季时呈显著正相关,其他季节无明显规律,物种数与生物量无显著关系。春季藻类的群落特征为生物量全年最低,多样性也在减少。通常,只有在尚未达到平衡或遭受干扰的生态系统中,才会观察到多样性与生物量生产之间的正相关[33]。苗绳附着藻类的优势种以绿藻和红藻为主,在秋季铜藻也是优势种。多样性对生产力的积极影响更多的是由于物种间的互补性机制,包括对资源的有效利用、生态位分配、植食动物的促进和控制等因素[13]。因此,苗绳上附着藻类优势种的不同类型所能利用的空间、环境、营养盐等资源也不尽相同;物种间的互补性抵消了竞争的负面影响,群落整体处于生长发育阶段,表现出的多样性与生物量呈正相关[33]。在一年生的作物与树木和/或动物相关联的农林复合系统中,往往会具有这样的特征,即异质群落比单一栽培生物的生产力更高[34]。而冬季当群落生长到一定阶段,则与浮子上附着的藻类一样,产生了同质竞争,香农指数与生物量呈负相关。
已知的多样性较低的农业生态系统中,有限的物种之间,可以通过功能互补和促进,从而提高资源的利用度,提高生产力增加生物量[34-35]。一年生植物的田间试验表明,多样性对生产力的影响呈正相关,多样性的增加能使群落更有效地利用土壤养分[36]。农业群落中的“优势种”造成了多样性的丧失和大部分生物量的增加,适当的多样性补充,能提高整体作物的生物量[37]。
4 结论
综上所述,贻贝筏式养殖区的藻类群落多样性与生物量,在夏季的浮子上表现出显著的负相关,而在秋季的苗绳上则表现为正相关。藻类群落的多样性与生物量未呈现明显的驼峰关系,多样性的增加不能有效提高单位面积的生物量。此外,浮子与苗绳上的藻类优势种有差异,由此说明筏架养殖区不同水层和不同设施材质所形成的空间异质性,可以扩展附着藻类的生态位,这也为使用不同附着基质开展多种藻类养殖[22]提供了参考。因此,针对枸杞岛贻贝养殖筏架区,可以适度引入本地经济藻种,如裙带菜、铜藻等,采用经济藻类与贻贝混养、藻类轮种等方式,优化贝藻养殖模式,这将是有效的增汇手段之一。
猜你喜欢
检察风云(2024年10期)2024-05-24 17:38:09
当代水产(2021年8期)2021-11-04 08:49:00
船舶力学(2021年3期)2021-03-29 10:06:52
军事文摘(2020年20期)2020-11-16 00:31:40
烹调知识(2020年10期)2020-10-20 05:47:44
今日农业(2019年10期)2019-01-04 04:28:15
中国惯性技术学报(2018年4期)2018-11-08 10:56:40
环境科技(2016年2期)2016-11-08 12:18:22
浙江大学学报(工学版)(2016年2期)2016-06-05 09:20:51
水产养殖(2016年2期)2016-03-28 11:47:52
