
不同时期水氮调控下高粱根系生理特征
2024-05-11 00:00:00王坚强闫素仙李慧明李作一史红梅
山西农业大学学报(自然科学版) 2024年1期




摘要:[目的]水分和氮肥是影响干旱地区作物生长的关键因素。探究水氮互作对高梁根系的调控作用可以增加高粱产量,提高水肥利用效率。[方法]本文以2 个高粱品种晋杂17 和晋杂108 为试验材料,通过设置重度干旱胁迫(W1,土壤含水量25%~35%)、中度干旱胁迫(W2,土壤含水量45%~55%)和正常供水处理(W3,土壤含水量65%~75%)3 个水分处理及低氮(N1,尿素0 g·kg−1 土)、中氮(N2,尿素2 g·kg−1 土)和高氮(N3,尿素4 g·kg−1 土)3 个氮素处理,分析不同水氮处理对各生育期的不同品种高粱根系生理特征的影响。[结果](1)水分处理对高粱根系各指标影响显著高于氮素处理,重度干旱胁迫下各指标降幅显著。(2)高粱的根重和根系活力在相同水分条件下随着施氮量的增加而升高,在中氮水平变化显著。可溶性糖、可溶性蛋白及脯氨酸含量随着施氮量的增加逐渐积累,在中氮和高氮下积累较快。(3)水氮处理对晋杂108 抽穗期存在明显的互作效应,对晋杂17 则无显著影响。(4)在不同水氮处理下,晋杂108 根系的各项指标均高于晋杂17,具有较强的抗逆性。[结论]高粱根系各指标受供水量的影响较大。在干旱环境下,增施适量氮素可促进根系生物量,提高根系活力,维持较高的渗透调节功能,缓解水分胁迫对根系的损伤。不同高粱品种的水氮互作效应不同,晋杂108 的抗旱、抗瘠薄能力更强。
关键词:高粱; 根系; 水氮调控; 生理特征
中图分类号:S514 文献标识码:A 文章编号:1671-8151(2024)01-0024-10
水分和氮素是粮食生产需求量最大的两大自然资源。近年来,由于经济快速发展,人为干扰和自然因素导致干旱气候频繁出现,大气氮沉降明显增加[1]。粮食作物的氮肥施用量过大,氮肥利用效率低下等问题日益突出。基于这些现状,我国农业生产也越来越注重节水、控肥的有机结合。
干旱胁迫是限制作物生长发育和产量的主要因素,受人们广泛关注。与其他非生物胁迫一样,植物的水分胁迫引起体内发生氧化应激反应,活性氧显著增加。大量研究表明,植物中抗氧化酶的活性、渗透调节能力与对干旱等胁迫的耐受性有关[2]。在逆境条件下,作物通过积累大量渗透调节物质,使组织水势下降,维持细胞膨压,进而保证体内代谢活动的正常运行,增强抗逆性。无机离子如K+、Ca2+等,有机物质如可溶性糖、可溶性蛋白和脯氨酸等[3]均为渗透调节物质。邵艳军等[4]研究发现,高粱叶片水势较大,以可溶性糖和蛋白质为主要渗透调节物质,玉米则以K+和脯氨酸为主。可溶性糖在苗期为高粱生长发育中氮同化提供碳骨架α-酮戊二酸,而α-酮戊二酸在调控植物碳氮代谢方面起到重要作用[5]。脯氨酸干旱胁迫下会快速积累,不仅能够保持细胞膨压,还具有猝灭自由基,保护亚细胞结构和缓冲细胞氧化还原电位的作用[6]。Rajasheker等[7]对高梁全基因组进行系统分析,鉴定出48 个编码富含脯氨酸蛋白的相关基因,并发现这些基因在维持细胞结构及响应逆境胁迫方面发挥关键作用。水分胁迫还可能导致作物不同营养元素缺乏甚至失衡。其中,氮元素参与植物体内多种代谢过程,是植物正常生长发育所必需的关键元素。一定水平的氮可通过维持代谢活动来缓解干旱胁迫对植物的负面影响,有助于改善作物产量,提高籽粒蛋白质、脂肪等含量[8-10]。如有人发现提高氮素能通过增强NO3−、NH4+的同化和脯氨酸的合成等多种途径提高水稻对水分胁迫的适应[11]。合理的水氮配比还可增强土壤酶活性,改善土壤透气性,提高肥力[12]。Shuo 等[13]通过建立甜高粱水肥耦合模型,确立了最佳的水肥配比,显著增加甜高粱干物质积累和产量,并发现灌溉是影响水利用效率和施氮的主要因素。水分和氮素之间存在复杂的交互作用,深入探究和揭示水氮交互作用对植物影响有助于农业上实现“以肥调水,以水促肥”目的。
根系生长是一个动态过程,且根系结构易随环境因素的变化而变化[14]。大量研究表明,根系性状与植株的抗旱性和养分吸收能力直接相关[15]。如干旱胁迫下,持绿型高粱比普通高粱能保持较好的根系形态和较高的根系活力,保证水分的吸收[16]。在低氮条件下,小麦根系形态、根系活力和氮代谢关键酶活性对氮素吸收具有重要的调控作用[17]。李文娆等[18]发现干旱虽会抑制棉花幼苗的根生物量,但根冠比、总根长增加,较高的根长和表面积可促进植株对水分的吸收,以应对干旱对冠层生长的影响。干旱条件下补充氮肥会促进根系表面积显著增加,以此提高根系形态对逆境的适应性。丁红等[19]发现干旱胁迫处理下增施氮肥,40 cm 以下土层内花生的根长、根系生物量和根表面积均显著增加,促进根系伤流量,提高了花生产量。根系的渗透调节机制对植物适应不良环境具有重要意义。对甘薯不同生长期时期进行干旱胁迫,均会导致甘薯根长、根活力下降,纤维根和块根中的可溶性糖等渗透调节物质上升,且干旱胁迫时间越早,升高的幅度越大[20]。张楚等[21]通过低氮胁迫不同品种的苦荞发现苗期根系直径、表面积、根系活力显著下降,主根长、可溶性糖含量及游离脯氨酸含量显著升高,且不同的耐低氮性品种对低氮胁迫的响应存在明显差异。
水分和氮素既是影响旱地农业的主要胁迫因子,又是一对相互作用的因子[22]。然而,以往研究多数只关注水分或氮素的单一影响,有关二者的交互作用对高粱根系的影响迄今研究较少。山西地处半干旱地区,而高粱(Sorghum bicolor L.)作为北方重要的粮食作物,其根系发达,抗旱、耐盐碱、耐贫瘠,具有较强的抗逆性和较高的养分利用效率[23]。卢峰等对高粱幼苗进行不同水平的水分胁迫,发现不同梯度下根系受影响程度大于叶片[24]。因此,深入研究水氮互作对高粱根系生理特征的影响,对揭示高粱根系的抗逆性机制、提高水肥利用效率具有重要意义。本试验以前期筛选出的氮素利用率高的晋杂108 和氮素利用率较低的晋杂17 为试验材料,2 个品种均为抗旱型品种。通过分析不同水分条件下施氮量对高粱不同生长期根系的生理特征的影响,探索水氮供应的最佳效果,以期为干旱地区对高粱进行合理水肥管理,提高作物产量提供理论依据。
1 材料与方法
1. 1 试验材料
本研究选用氮素利用率不同的晋杂17 和晋杂108 高粱品种为试材,均由山西农业大学高粱研究所提供。试验于2021 年5 月-10 月在晋中市榆次区修文基地棚内进行。试验土壤取自表层0~20 cm土壤,基础肥力状况为:土壤有机质11. 07 g·kg−1,全氮0. 76 g·kg−1,有效磷11. 32 mg·kg−1,速效钾135. 10 mg·kg−1,pH 值8. 1。
1. 2 试验方法
试验针对水分和氮肥2 因素分别设3 个梯度,共计9 个处理。3 个水分水平:W1、W2 和W3 分别表示重度干旱胁迫(土壤含水量25%~35%)、中度干旱胁迫(土壤含水量45%~55%)和正常供水处理(土壤含水量65%~75%);3 个氮肥水平:N1、N2和N3 分别表示低氮(尿素0 g·kg−1土)、中氮(尿素2 g·kg−1土)和高氮(尿素4 g·kg−1土)。每个处理进行3 次重复。氮、磷、钾分别以尿素、过磷酸钙和硫酸钾为基肥施入,P2O5和K2O 施用量为0. 2 g·kg−1。本试验采取盆栽方式,高粱生长到4 叶期(播种后14 d)开始控制土壤含水量,土壤含水量的测定采用称重法。盆栽盆上口径23 cm,高18 cm,选取饱满、均匀的高粱种子,5 月19 日每盆播种10 粒,在三叶期定苗,每盆保留3 株。于苗期(播种后19 d)、拔节期(播种后39 d)、抽穗期(播种后67 d)进行取样调查测定根系形态、根系活力和各渗透物质含量。
1. 3 测定项目与方法
选取每处理具有代表性的植株进行取样,取样前先浇水使土壤变软,用清水冲洗干净,烘干称重,105 ℃杀青30 min,80 ℃烘干至恒重。剩余采集的新鲜根样放入液氮备用。采用氯化三苯基四氮唑法(TTC 法)测定根系活力[25];采用蒽酮比色法测定根系可溶性糖含量[26];采用考马斯亮蓝G-250 染色法测定根系可溶性蛋白含量[27];采用茚三酮显色法测定根系游离脯氨酸含量[28]。
1. 3 数据处理
用Excel 2007、Origin 9. 1 对数据进行处理并作图,SPSS 19. 0 软件进行单因素和双因素方差分析和差异显著性分析。
2 结果与分析
2. 1 不同时期水氮互作下高粱根系干重
由表1 可见,2 品种高粱的根系干重在正常供水(W3)和高氮(N3)条件下达到最大值。在不同时期各处理下晋杂108 的根系干重均高于晋杂17。低氮条件下,晋杂17 根系干重W1 和W2 比W3 减少25. 24%、12. 81%(苗期),13. 31%、6. 61%(拔节期),13. 85%、4. 34%( 抽穗期);中氮下减少18. 18%、17. 25%(苗期),8. 01%、8. 31%(拔节期),5. 28%、0. 62%( 抽穗期);高氮下减少19. 29%、19. 19%(苗期),9. 07%、6. 81%(拔节期),6. 25%、3. 76%(抽穗期)。晋杂108 根系干重W1 和W2 比W3 减幅4. 64%~16. 69%。在施氮条件相同时,水分的减少均可抑制根系干重的增加,W1 降幅显著且苗期受水分影响较大。在各生育期不同水分水平下,增施氮肥亦可促进根系干重的积累。在中度干旱胁迫下,晋杂17 与晋杂108 的根系干重在不同处理下最大增幅为2. 5%、2. 7%,增长不明显,且不同氮素水平下差异不大。在重度干旱胁迫(W1)下,晋杂17 根系干重在高氮(N3)与中氮(N2)下与不施氮相比分别增长−1. 36%、12. 79%(苗期),− 1. 23%、11. 15%(拔节期),3. 08%、7. 64%(抽穗期);晋杂108 增长4. 42%、5. 40%(苗期),0. 43%、4. 75%(拔节期),2. 54%、3. 71%(抽穗期)。可见,在重度干旱胁迫下适量的氮肥可以促进根系生物量的增长速度,但施氮过多会破坏根细胞膜功能,减缓甚至抑制根系生长,且在苗期和拔节期根系增长较快。
2. 2 不同时期水氮互作下高粱根系活力
根系活力主要指脱氢酶活性,根系活力直接决定地上部分的生长和营养状况。由图1 可见,总体变化趋势晋杂108 的根系活力高于晋杂17。在相同氮素条件下,2 品种各生育期W1 和W2 与W3 相比,下降幅度14. 91%~38. 0%、4. 98%~27. 71%。高粱根系活力均随着水分的减少而降低,W1 下降幅显著,2 品种各生育期降幅差异不大。在不同生育期,晋杂108 和晋杂17 的根系活力在相同水分条件下随着施氮量的增加而升高,但增幅不同。2 品种的根系活力在W1 和W3 下与W2 相比,增施氮肥增幅较小。晋杂108 在中度干旱胁迫(W2)下,N2 和N3 提高19. 63%、15. 57%( 苗期)、10. 35% 和8. 51%(拔节期)、11. 71% 和7. 23%(抽穗期),苗期根系活力在中氮条件下增幅最大。晋杂17 中度干旱胁迫下(W2),N2 和N3 提高14. 79% 和12. 54%(苗期)、16. 78% 和9. 55%(拔节期)、8. 12% 和11. 24%(抽穗期),拔节期N2 下增幅最大。可见,晋杂108 受中度干旱胁迫时,在苗期适量施用氮肥能显著提高根系活力;晋杂17 在拔节期受氮肥影响大。由此可知,不同高粱品种的根系活力对水氮互作的响应不同。
2. 3 不同时期水氮互作下高粱根系渗透调节物质含量
2. 3. 1 可溶性糖含量
由图2 可见,在相同氮素条件下,晋杂17 和晋杂108 各生育期在W1 和W2 下与W3 相比,增长幅度为9. 10%~28. 50%、3. 05%~19. 17%。高粱根系可溶性糖含量均随着水分的减少而增加,W1 下增长明显,2 品种各生育期增幅差异不大。此外,重度干旱胁迫(W1)时,晋杂17 根系可溶性糖含量N2和N3 比N1 增长51. 58%、30. 16%( 苗期),10. 80%、4. 53%(拔节期),9. 63%、5. 36%(抽穗期);W2 下增长52. 99%、30. 42%(苗期),9. 71%、6. 19%(拔节期),6. 13%、3. 32%(抽穗期);W3 下增长56. 71%、19. 81%(苗期),11. 29%、4. 89%(拔节期),6. 69%、3. 63%(抽穗期)。晋杂108 根系可溶性糖含量在相同供水条件下,N2 和N3 比N1 增加11. 76%~47. 55%(苗期),3. 20%~11. 41%(拔节期),3. 58%~6. 05%(抽穗期)。可见,在供水相同情况下,晋杂17 高粱根系的可溶性糖含量在苗期和拔节期受氮肥影响大,而晋杂108 则是苗期变化显著。此外,晋杂17 的可溶性糖含量在各种处理下均比晋杂108 高。
2. 3. 2 可溶性蛋白含量
随着生育期的推进,高粱根系可溶性蛋白含量在抽穗期达到最高(图3)。在不同生育期,晋杂108比晋杂17 的可溶性蛋白含量高。与高粱根系可溶性糖含量变化相似,在施氮量相同时,可溶性蛋白随着供水量的减少而显著增长,在W3 下与W1 比,抽穗期增幅达39. 75%~43. 97%,增幅最大。在相同水分条件下,高粱根系可溶性蛋白含量随施氮量的增加而增加。在抽穗期中度干旱胁迫时,晋杂17和晋杂108 在中氮、高氮下分别上升− 3. 81% 和7. 76%、20. 85% 和8. 01%,重度干旱胁迫时,晋杂17 和晋杂108 在中氮、高氮下分别下上升9. 01% 和7. 78%、5. 35% 和4. 84%。表明中度干旱胁迫下,高粱根系可溶性蛋白含量在高氮水平时增幅最大,重度干旱胁迫时中氮水平增幅较大。晋杂17 在拔节期和抽穗期受氮素影响极明显。
2. 3. 3 脯氨酸含量
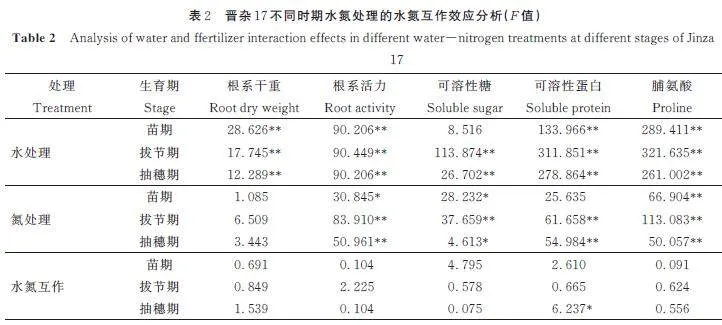
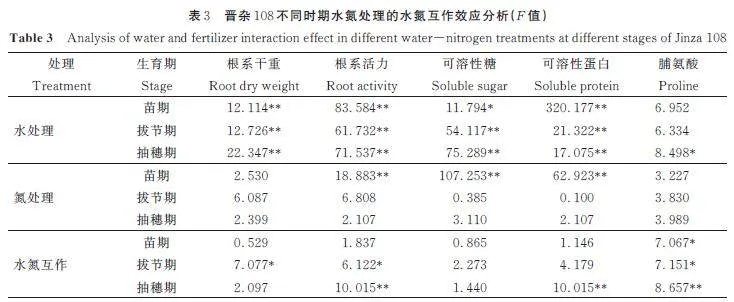
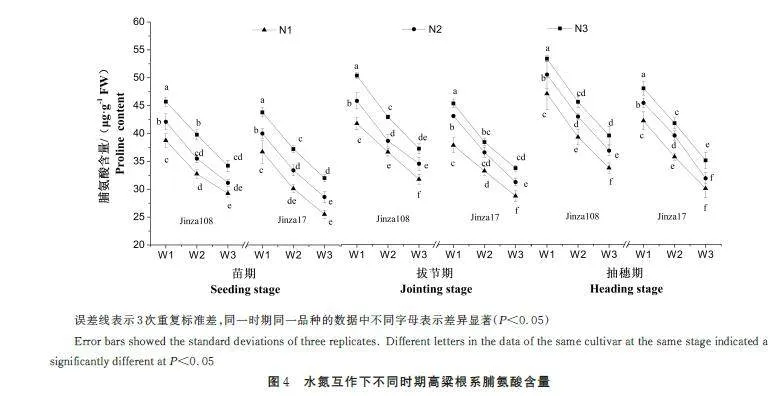
高粱根系的脯氨酸含量随着供水量的减少下整体增幅在33. 6%~43. 9%,与可溶性糖和可溶性蛋白含量相比增幅最大(图4)。这是因为植物细胞中脯氨酸含量一般较低,但受干旱胁迫时会快速积累,且积累越多,抗旱能力越强[6]。不同时期的水氮处理下,晋杂108 根系的脯氨酸含量较高。方差分析结果表明水氮互作效应对晋杂108 根系脯氨酸含量的影响达显著水平(表2~表3)。在相同水分条件下,增加施氮量可显著促进高粱根系脯氨酸的积累,尤其在重度缺水时。在重度干旱胁迫时,N2 和N3 水平下比N1,各时期晋杂108 的根系脯氨酸含量增长6. 11% 和9. 85%、8. 83% 和7. 82%、9. 29%和7. 33%;晋杂17 的根系脯氨酸含量增长10. 83%和11. 88%、8. 70% 和8. 01%、6. 01% 和10. 04%。晋杂17 在苗期和拔节期重度干旱胁迫时受氮素影响较大,而晋杂108 在抽穗期影响较大。施氮量一定时,高粱根系内脯氨酸含量随着供水量的增加而增加。在整个生长期,高粱根系的脯氨酸含量在抽穗期达到最大。
2. 4 各生理指标的水氮互作效应
水、氮单因素对不同时期高粱根系的各生理指标的影响不同。结果表明,水分处理对高粱根系生理指标的影响显著高于氮肥处理(表2~表3)。其中根系干重受水分变化的影响极显著,而氮素影响并不显著。这说明无论任何时期,水分都是高粱根系生长所需的关键因素。不同高粱对不同水氮处理的响应也存在差异。氮肥处理对晋杂17 拔节期和抽穗期的根系各项生理指标(除根系干重)影响都极为显著,而对晋杂108 则无显著影响。水氮互作效应对晋杂108 抽穗期的影响最为显著,而且越到高粱生育后期,水氮互作效应越强。
3 讨论
本研究发现,高粱根系干重、根系各渗透物质含量随着生育期的推进逐渐增加,在抽穗期达到最大,根系活力逐渐降低,在抽穗期最小。
水分和氮素是影响高粱生长发育和产量形成的重要的环境因子,两者相互影响,相互制约。干旱会降低土壤水势,加速根系死亡,抑制植物氮吸收,而高效的氮营养能通过维持代谢活动来缓解干旱胁迫对植物的负面影响[29]。本研究中高粱根系各指标均受水分处理的影响较大,尤其脯氨酸含量变化幅度达33. 6%~43. 9%。这与李广浩等[30]认为的水氮耦合效应中水为主导效应,氮为次效应的结论一致。氮对植物抗旱能力的调节与氮水平和干旱胁迫强度有关[31]。此外还发现在不同生育期,2高粱品种的根系干重和根系活力在相同水分条件下随着施氮量的增加而升高,在水分胁迫时中氮(尿素2 g·kg−1土)水平分别增长12. 79%、19. 63%(晋杂17 苗期),5. 40%、14. 79%(晋杂108 苗期),增加较快。受水分胁迫时高氮(尿素4 g·kg−1土)和中氮水平下与不施氮相比(以苗期为例,其余时期结果相似),根系可溶性糖含量(晋杂17 苗期增长51. 58%、30. 16%,晋杂108 苗期增长33. 42%、21. 36%)、可溶性蛋白含量(晋杂17 苗期增长11. 17% 和6. 71%、晋杂108 苗期增长6. 08% 和3. 10%)与脯氨酸含量(晋杂17 增长10. 83% 和11. 88%、晋杂108 增长6. 11% 和9. 85%),均上升显著。高粱根系在受干旱胁迫时,适量增加氮肥,可促进根系生长发育,提高根系的抗旱能力。这与前人研究结果相似[19,21,32-33]。但值得注意的是,相同水分条件下氮素对根系形态的影响目前存在争议。有研究认为在水分胁迫下增施氮素会造成玉米根部水势降低,抑制根的生长[34]。这可能是由于氮素条件不同和不同植物抗逆性不同造成的。
良好的根系形态和较高的根系生理活性是植物氮高效利用重要特征[35]。渗透调节物质含量可间接体现植物抗旱能力[36],其中脯氨酸能够保护细胞膜和细胞内酶系统,在干旱胁迫下,其含量增加会降低细胞水势,增强植物吸水能力。在各生育期水氮处理下,晋杂108 根系干重(5. 74~30. 77 g)、根系活力(55. 99~84. 16 μg·g−1·h−1)、可溶性蛋白含量(18. 03~20. 83 mg·g−1)和脯氨酸含量(33. 71~45. 72 μg·g−1)均高于晋杂17。说明晋杂108 对水分和氮肥的吸收能力更强,更能适应缺水缺氮环境,抗旱、抗瘠薄能力更强。在相同水分条件下,不同品种的高粱施氮效应不尽相同。晋杂17的施氮效应对于拔节期和抽穗期的根活力、可溶性糖含量、可溶性蛋白含量、脯氨酸含量影响显著,晋杂108 在苗期较显著。可能是因为不同基因型背景下,高粱对氮素的吸收和利用效率存在时间差异,其生理和分子机制还需进一步试验证明。不同品种的高粱对于水氮互作效应存在明显差异。水氮互作效应对晋杂108 抽穗期的影响最为显著,而且越到高粱生育后期,水氮互作效应越强。这是由于高粱生育后期生长需要大量水分和营养,使得水氮叠加效应逐渐显现。
4 结论
通过对不同水氮组合处理苗期、拔节期和抽穗期的高粱根系干重、根系活力和渗透调节物质的研究发现,高粱根系各指标受水分处理的影响大于氮素处理。在水分胁迫下,增施适量氮肥会促进高粱根系的生物量和根系活力的增长,促进可溶性蛋白、可溶性糖和脯氨酸的积累,有效缓减干旱胁迫的负面影响。此外,水氮对晋杂108 抽穗期的根系生长存在显著的互作效应。晋杂108 具有较强的抗旱和耐低氮能力。
参考文献
[1]陈晓娜,赵纳祺,段娜,等. 植物响应水分和氮添加的研究进展[J]. 温带林业研究, 2022, 5(1): 7-11.
Chen X N, Zhao N Q, Duan N, et al. Plant response to waterand nitrogen addition:a review [J]. Journal of TemperateForestry Research, 2022, 5(1): 7-11.
[2]万劲,汪安琳,陈嘉裔,等. 植物抗逆生理研究概述[J]. 江苏林业科技, 2013, 40(1): 42-45.
Wan J, Wang A L, Chen J Y, et al. Overview of plant stressresistance physiology research[J]. Journal of Jiangsu ForestryScience amp; Technology, 2013, 40(1): 42-45.
[3]韩志平,张海霞,张巽,等. 水分胁迫对黍子幼苗生长和生理特性的影响[J]. 中国农业气象, 2019, 40(8): 502-511.
Han Z P, Zhang H X, Zhang X, et al. Effects of water stress ongrowth and physiological properties of millet seedlings[J].Chinese Journal of Agrometeorology, 2019, 40(8): 502-511.
[4]邵艳军,山仑,李广敏. 干旱胁迫与复水条件下高粱、玉米苗期渗透调节及抗氧化比较研究[J]. 中国生态农业学报, 2006, 14(1): 68-70.
Shao Y J, Shan L, Li G M. Comparison of osmotic regulationand antioxidation between sorghum and maize seedlings undersoil drought stress and (water) recovering conditions [J].Chinese Journal of Eco-Agriculture, 2006, 14(1): 68-70.
[5]Yuan Y Z, Ou J Q, Wang Z Q, et al. Regulation of carbon andnitrogen metabolisms in rice roots by 2-oxoglutarate at the levelof hexokinase[J]. Physiologia Plantarum, 2007, 129(2):296-306.
[6]Ashraf M, Foolad M R. Roles of glycine betaine and proline inimproving plant abiotic stress resistance[J]. Environmental andExperimental Botany, 2007, 59(2): 206-216.
[7]Rajasheker G, Nagaraju M, Varghese R P, et al. Identificationand analysis of proline-rich proteins and hybrid proline-richproteins super family genes from Sorghum bicolor and theirexpression patterns to abiotic stress and zinc stimuli [J].Frontiers in Plant Science, 2022, 13: 952732.
[8]Nematpour A, Eshghizadeh H R, Zahedi M. Drought-tolerancemechanisms in foxtail millet (Setaria italica) and proso millet(Panicum miliaceum) under different nitrogen supply andsowing dates[J]. Crop and Pasture Science, 2019, 70(5):442-452.
[9]于肖,牛佳红,陈二影,等. 施氮与不同时期水分胁迫对谷子生长及生理生化特性的影响[J]. 山东农业科学, 2022, 54(1):61-67.
Yu X, Niu J H,Chen E Y, et al. Effects of nitrogen applicationand water stress at different stages on growth and physiologicaland biochemical characteristics of foxtail millet[J]. ShandongAgricultural Sciences, 2022, 54(1): 61-67.
[10]李昱,俞自仁,吴伟,等. 水肥耦合对甜高粱新品种中科甜2 号生物学特性和产量的影响[J]. 甘肃农业科技, 2017(10):15-21.
Li Y, Yu Z R, Wu W, et al. Effects of water and fertilizercoupling on biological characteristics and yield of sweetSorghum cultivar zhongketian 2 [J]. Gansu AgriculturalScience and Technology, 2017(10): 15-21.
[11]Zhong C, Cao X C, Hu J J, et al. Nitrogen metabolism inadaptation of photosynthesis to water stress in rice grown underdifferent nitrogen levels[J]. Frontiers in Plant Science, 2017,8: 1079.
[12]樊吴静,杨鑫,何虎翼,等. 水氮耦合对旱藕产量、生理特性和土壤理化性状的影响[J]. 西南农业学报, 2022, 35(5): 1069-1078.
Fan W J, Yang X, He H Y, et al. Effect of water and nitrogencoupling on the yield and physiology characteristics of ediblecanna and the physical and chemical properties of soil[J].Southwest China Journal of Agricultural Sciences, 2022, 35(5): 1069-1078.
[13]Shuo W, Chai C L, Liu H Q. Effects of water and nitrogenregulation on dry matter accumulation, yield and water use ofsweet Sorghum[J]. World Scientific Research Journal, 2021,7(5): 316-327.
[14]王岩,孙大为,李兆波,等. 不同品种高粱根系形态特征与产量形成的关系[J]. 园艺与种苗, 2011, 31(3): 84-86.
Wang Y, Sun D W, Li Z B, et al. Study on relationship ofroot morphological characters and yeild formation in differentSorghum varieties[J]. Horticulture amp; Seed, 2011, 31(3):84-86.
[15]Zhang Y J, Xu J N, Cheng Y D, et al. The effects of waterand nitrogen on the roots and yield of upland and paddy rice[J].Journal of Integrative Agriculture, 2020, 19(5): 1363-1374.
[16]王德权,周宇飞,陆璋镳,等. 水分胁迫下持绿型高粱根系形态及其活力研究[J]. 干旱地区农业研究, 2012, 30(2): 73-76, 130.
Wang D Q, Zhou Y F, Lu Z B, et al. Root morphology andactivity of stay green sorghum under water stress [J].Agricultural Research in the Arid Areas, 2012, 30(2): 73-76, 130.
[17]海闯北,彭超军,李艳,等. 高氮与低氮处理对不同氮效率小麦根系及相关生理特性的影响[J]. 麦类作物学报, 2022, 42(6): 697-703.
Hai C B, Peng C J, Li Y, et al. Effect of high and lownitrogen treatments on roots and related physiologicalcharacteristics of wheat with different nitrogen efficiency[J].Journal of Triticeae Crops, 2022, 42(6): 697-703.
[18]李文娆,范雨龙,冯士珍,等. 水氮耦合对棉花幼苗根冠生长和水分利用效率的影响[J]. 河南农业科学, 2017, 46(9):18-24.
Li W R, Fan Y L, Feng S Z, et al. Effect of coupling of waterand nitrogen on shoot and root growth and water use efficiencyof cotton seedlings[J]. Journal of Henan Agricultural Sciences,2017, 46(9): 18-24.
[19]丁红,张智猛,戴良香,等. 水氮互作对花生根系生长及产量的影响[J]. 中国农业科学, 2015, 48(5): 872-881.
Ding H, Zhang Z M, Dai L X, et al. Effects of water andnitrogen interaction on peanut root growth and yield [J].Scientia Agricultura Sinica, 2015, 48(5): 872-881.
[20]张海燕,汪宝卿,冯向阳,等. 不同时期干旱胁迫对甘薯生长和渗透调节能力的影响[J]. 作物学报, 2020, 46(11): 1760-1770.
Zhang H Y, Wang B Q, Feng X Y, et al. Effects of droughttreatments at different growth stages on growth and the activityof osmotic adjustment in sweet potato [Ipomoea batatas (L.)Lam.[J]. Acta Agronomica Sinica, 2020, 46(11): 1760-1770.
[21]张楚,张永清,路之娟,等. 低氮胁迫对不同苦荞品种苗期生长和根系生理特征的影响[J]. 西北植物学报,2017,37(7):1331-1339.
Zhang C, Zhang Y Q, Lu Z J, et al. Effect of low nitrogenstress on the seedling growth and root physiological traits ofFagopyrum tataricum cultivars with different low-N treatments[J]. Acta Botanica Boreali-Occidentalia Sinica, 2017, 37(7):1331-1339.
[22]张志远, 李玉庆. 我国水肥耦合研究热点及趋势探析[J]. 安徽农业科学, 2022, 50(5): 220-223, 227.
Zhang Z Y, Li Y Q. Analysis on the hotspots and trends ofwater-fertilizer coupling research in China[J]. Journal of AnhuiAgricultural Sciences, 2022, 50(5): 220-223, 227.
[23]李顺国,刘猛,刘斐,等. 中国高粱产业和种业发展现状与未来展望[J]. 中国农业科学, 2021, 54(3): 471-482.
Li S G, Liu M, Liu F, et al. Current status and futureprospective of Sorghum production and seed industry in China[J]. Scientia Agricultura Sinica, 2021, 54(3): 471-482.
[24]卢峰,张飞. 多水胁迫对高粱幼苗的生理影响[J]. 作物杂志,2015(4): 92-96.
Lu F, Zhang F. Physiological responses of Sorghum seedlingsunder excessive water stress[J]. Crops, 2015(4): 92-96.
[25]闫江艳,张永清,冯晓敏,等. 干旱胁迫及复水对不同黍稷品种根系生理特性的影响[J]. 西北植物学报, 2012, 32(2):348-354.
Yan J Y, Zhang Y Q, Feng X M, et al. Effect of droughtstress and rewatering on physiological characteristics of roots indifferent proso millet varieties[J]. Acta Botanica Boreali-Occidentalia Sinica, 2012, 32(2): 348-354.
[26]孔祥生,易现峰. 植物生理学实验技术[M]. 北京:中国农业出版社, 2008: 252-253.
Kong X S, Yi X F. Experiment technology of plant physiology[M]. Beijing: China Agriculture Press, 2008: 252-253.
[27]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社, 2000: 59-60.
Li H S. Principles and techniques of plant physiologicalbiochemical experiment[M]. Beijing: Higher Education Press,2000: 59-60.
[28]张志良,瞿伟菁. 植物生理学实验指导[M]. 3 版. 北京:高等教育出版社, 2003: 38-39.
Zhang Z L, Qu W J. Experimental guide for plant physiology[M]. 3rd edition. Beijing: Higher Education Press, 2003:38-39.
[29]Yuan Y H, Liu L, Gao Y B, et al. Comparative analysis ofdrought-responsive physiological and transcriptome inbroomcorn millet (Panicum miliaceum L.) genotypes withcontrasting drought tolerance [J]. Industrial Crops andProducts, 2022, 177: 114498.
[30]李广浩,赵斌,董树亭,等. 控释尿素水氮耦合对夏玉米产量和光合特性的影响[J]. 作物学报, 2015, 41(9): 1406-1415.
Li G H, Zhao B, Dong S T, et al. Effects of couplingcontrolled release urea with water on yield and photosyntheticcharacteristics in summer maize[J]. Acta Agronomica Sinica,2015, 41(9): 1406-1415.
[31]Fan J B, Zhang Y L, Turner D, et al. Root physiological andmorphological characteristics of two rice cultivars with differentnitrogen-use efficiency [J]. Pedosphere, 2010, 20 (4) :446-455.
[32]杜红霞,冯浩,吴普特,等. 水、氮调控对夏玉米根系特性的影响[J]. 干旱地区农业研究, 2013, 31(1): 89-94,100.
Du H X, Feng H, Wu P T, et al. Influence of water and Nfertilizer regulation on root growth characteristics of summermaize[J]. Agricultural Research in the Arid Areas, 2013, 31(1): 89-94, 100.
[33]孙婷, 张迪, 王冀川,等. 滴灌水氮运筹对南疆春小麦根系生长及产量的影响[J]. 干旱地区农业研究, 2020, 38(2):10-20.
Sun T, Zhang D, Wang J C, et al. Effects of drip irrigationand nitrogen application on root growth and yield of springwheat in Southern Xinjiang[J]. Agricultural Research in theArid Areas, 2020, 38(2): 10-20.
[34]邢换丽,周文彬,郝卫平,等. 水分胁迫下氮素增加对玉米生长的抑制作用[J]. 中国农业气象, 2020, 41(4): 240-252.
Xing H L, Zhou W B, Hao W P, et al. Inhibition of nitrogenincreasing on maize growth under water stress[J]. ChineseJournal of Agrometeorology, 2020, 41(4): 240-252.
[35]Tran T T, Kano-Nakata M, Takeda M, et al. Nitrogenapplication enhanced the expression of developmental plasticityof root systems triggered by mild drought stress in rice[J].Plant and Soil, 2014, 378(1): 139-152.
[36]司瑞,刘冰,朱钊岑,等. 西北干旱区甜高粱种植水肥配比模式研究[J]. 灌溉排水学报, 2021, 40(5): 54-61.
Si R, Liu B, Zhu Z C, et al. Optimal fertigation and irrigationfor sweet Sorghum production in arid regions in NorthwestChina[J]. Journal of Irrigation and Drainage, 2021, 40(5):54-61.
(编辑:郭玥微)
猜你喜欢
快乐作文(1.2年级)(2023年9期)2023-05-12 11:31:34
现代园艺(2022年7期)2022-11-19 10:18:42
古今农业(2022年2期)2022-08-15 01:39:52
青年文学家(2022年1期)2022-03-11 12:27:39
四川劳动保障(2021年3期)2021-06-09 07:08:56
疯狂英语·新读写(2021年2期)2021-02-25 08:58:46
湖南饲料(2019年5期)2019-10-15 08:59:10
现代园艺(2017年13期)2018-01-19 02:28:05
西南农业学报(2016年5期)2016-05-17 05:42:36
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:09
