
重组新城疫病毒rL-RVG通过p53-YAP1-ACSL4通路诱导肺腺癌细胞铁死亡
2024-05-10 10:11:00何瑛珏田仪督贡克文严玉兰
医学研究生学报 2024年1期
何瑛珏,李 洋,田仪督,贡克文,严玉兰
0 引 言
肺癌已成为全球癌症患者的首要威胁,导致的死亡人数比乳腺癌、前列腺癌、结直肠癌的总和都要多,每天大约350例患者死于肺癌,肺癌的患者比例也在持续上升,目前肺癌已成为全球第一大癌症[1]。肺癌一般分为小细胞肺癌和非小细胞肺癌,其中非小细胞肺癌的患者比例更为突出,按其病理形态又可分为:肺鳞癌、肺大细胞癌及肺腺癌等。肺腺癌是此类肺癌中最常见的类型,且具有高度的异质性和恶性[2]。除手术及放化疗外,抗癌药物触发凋亡使细胞死亡也是杀死癌细胞的经典方法[3]。然而,细胞凋亡治疗肿瘤的效果有限,随着癌细胞耐药性的逐年上升,许多新的细胞死亡过程被发现。铁死亡作为一种全新的细胞死亡机制开始被认识,它的主要表现形式就是由于铁的代谢而导致的致死性的脂质过氧化产物的积聚[4]。近期研究表明,铁死亡在肺癌的发生发展具有至关重要的意义[5],通过诱导肺癌细胞铁死亡可能会被视为一种抗肺癌治疗的新策略。
新城疫病毒(newcastle disease virus,NDV)具有高度的特异性,它能够通过营养剥夺、吞噬铁蛋白等机制诱导肿瘤细胞铁死亡[6]。研究证明NDV可通过上调胶质瘤细胞中p53的表达和抑制Xc-系统诱导脑胶质瘤细胞铁死亡[7]。而稳定表达狂犬病毒糖蛋白的重组新城疫病毒(recombinant newcastle disease virus with stable expression of rabies virus glycoprotein, rL-RVG)不仅可以发挥NDV的溶瘤效应,还可以发挥狂犬病毒糖蛋白的N受体阻断效应,它的抗癌能力显著优于NDV[8]。本课题组前期实验已经验证rL-RVG在抑制人肺腺癌细胞增殖过程中可能发挥类似铁死亡诱导剂Erastin作用,但rL-RVG导致肺腺癌细胞铁死亡的相关机制尚不明确,故本研究旨在探讨重组新城疫病毒rL-RVG诱导人肺腺癌细胞铁死亡的部分机制研究。
1 材料与方法
1.1 材料人肺腺癌细胞系A549、PC9,人正常支气管上皮细胞BEAS-2B(中国科学院上海生物科学研究院细胞中心);rL-RVG和NDV(哈尔滨兽医研究所);RPMI 1640培养基和胎牛血清(Gibco 公司);脂质过氧化( MDA) 检测试剂盒(碧云天生物公司);活性氧检测试剂盒(江苏凯基生物);BCA 蛋白定量试剂盒(南京福麦斯生物技术有限公司);兔抗YAP1、ACSL4抗体(Boster公司);鼠抗p53(CST 公司);鼠抗β-actin 抗体、羊抗鼠和羊抗兔二抗(武汉三鹰生物技术有限公司);SiRNA-YAP1、SiRNA-ACSL4(吉玛基因);CCK8试剂盒、RNA快速提取试剂盒、反转录试剂盒、SYBR qPCR Master Mix(均南京诺唯赞生物科技有限公司)。
1.2方法
1.2.1 细胞的培养与分组在恒温恒湿的环境下(37 ℃,5% CO2,100%的湿度)培养人肺腺癌细胞A549和PC9。待细胞长至80%~90%时,进行传代和铺板等处理。用病毒来感染细胞,将人肺腺癌细胞 A549、PC9 分为对照组(NC组)、NDV组(MOI=5)、rL-RVG组(MOI=5)、Erastin组(5 μmol/L)、rL-RVG(MOI=5)+ Erastin(5 umol/L)处理组、NAC组(5 μmol/L)、rL-RVG(MOI=5)+NAC(5 μmol/L)处理组、Si-NC对照组、SiRNA-YAP1干预组、SiRNA-YAP1+rL-RVG处理组、SiRNA-ACSL4干预组、SiRNA-ACSL4+rL-RVG处理组进行培养用于后续实验。
1.2.2CCK-8法检测细胞增殖能力在96孔板上接种细胞(1000个/孔),加入100 μL含10%胎牛血清的RPMI1640培育基,置于恒温恒湿的培养箱中培养,显微镜下观察细胞状态,贴壁后按照不同的病毒感染复数(MOI=1、3、5、10)进行处理,继续在细胞培养箱里进行培育,每个处理组都设置3个复孔。在0、24、48、72 h 4个时间段进行处理,每孔加入10 μL CCK8试剂,避光培养1 h后使用酶标仪在450 nm波长处对样本进行吸光度检测,以确定不同时间段样本的细胞活性。公式如下:
细胞活力(%)=(测量值-空白值)/(对照值-空白值)×100%
1.2.3细胞划痕实验检测迁移率根据不同实验组将细胞悬液接种于6孔板中,每组设置3个复孔;当细胞贴壁增殖汇合至 70%~80%时弃去板内原培养基,用PBS清洗3次,病毒感染后以10 μL枪头从孔板的底部开始沿着每个孔的中心横和纵方向画出一条直线。将6孔板放置于细胞培养箱中继续培养,显微镜下分别拍照记录0 h、24 h直线宽度,计算细胞迁移率。公式如下:
细胞迁移率(%)=(0 h宽度-24 h宽度)/0 h 宽度×100%
1.2.4Transwell细胞侵袭实验Matrigel基质胶经过(1∶10)的稀释,把它均匀地平铺于Transwell的各个小室,置于37 ℃的环境2 h时使其变成凝胶状。使用NDV、rL-RVG感染肺腺癌A549、PC9细胞24 h后,将细胞中的培养基更换成无血清培养液,饥饿12 h消化细胞后用含 5%血清的培养基重悬,细胞密度控制在5×105/mL,向上室中加200 μL上述细胞混合液,下室加入800 μL含20%血清的细胞培养基,继续放置在37 ℃细胞培养箱,孵育24 h,倒掉Transwell小室中的培养液,PBS清洗3次,用4%的多聚甲醛溶液固定,经过0.5 h的处理,将其脱离,使用0.1%的结晶紫溶液染色10 min后取出,并用PBS清洗小室中残留的染液,最后使用棉签轻轻地擦拭掉小室上层中没有穿透进入小室底的细胞,尽量减少对小室外观的破坏,以便更准确的分离和检测。在200倍倒置相差显微镜下记录视野中的细胞数并进行统计分析。实验均3次重复。
1.2.5ROS的检测流式细胞术:根据剂盒产品说明书,将不同处理组细胞置于6孔板(1 ×106/孔)中培养过夜。胰酶消化离心收集细胞后PBS洗涤2次,37 ℃下悬浮于 200 mL DCFH-DA工作溶液中约30 min,离心收集细胞后继续用PBS洗涤两次,用400 μL PBS重悬,使用流式细胞仪在485 nm的激发波长下检测细胞内ROS的水平。对于每组样品,评估大约20 000个细胞。设置3组重复进行。
ROS荧光强度测定:6孔板中接种细胞密度达80%左右,加入病毒感染24 h。将DCFH-DA荧光探针以1∶1000的比例稀释,放置于37 ℃、5% CO2的培养箱内,经过30 min的遮光处理,再经过PBS洗涤,通过荧光显微镜观察ROS荧光的强弱,进行相应的数据统计。
1.2.6MDA水平检测根据说明书要求,首先配置0.37%的TBA贮存液,然后按照比例配制新鲜的MDA检测工作液。在1.5 mL离心管中加入0.1 mL稀释成不同浓度的标准品和待测样品,随后加入0.2 mL MDA检测工作液,混匀后,99 ℃ 金属浴加热15 min,冷却至室温,1000×g室温离心10 min。取200 μL上清液加入到96孔板中,酶标仪在532 nm测定吸光度。通过标准曲线计算出样品溶液中的MDA浓度后,以单位重量的蛋白含量(BCA法测量) 来计算最初样品中的MDA含量(μmol/mg)。各处理组均设置3组重复。
1.2.7Western blot检测蛋白表达蛋白提取:将接种细胞后的6孔板置于冰上,加入适量裂解液于冰上裂解细胞,收集裂解液于4 ℃,12 000×g离心10 min后收集上清液。部分上清液以BCA蛋白浓度检测试剂盒检测各组总蛋白浓度。按照计算出的蛋白浓度取适量上清液加入4×Load Buffer,放于100 ℃水浴锅内水浴5 min,冷却至室温后-80 ℃保存。
Western blot:制胶后,取10 μL的上样量,恒压80V电泳30 min,待 marker的分层条带跑至分离胶下1~2 cm后,调整电压至120 V继续电泳1 h左右;200 mA恒流湿转120 min将蛋白转移至PVDF膜,将膜置于无蛋白酶快速封闭液中15 min;封闭后的条带浸泡在稀释后的一抗中置于4 ℃冰箱孵育过夜;TBST洗膜3次,每次10 min;再加入羊抗兔二抗、羊抗鼠二抗室温孵育1 h;继续TBST洗膜3次,每次10 min;最后置于ECL发光显影液内3 min;凝胶成像仪拍照保存。
1.2.8细胞总RNA提取及实时荧光定量 PCR根据诺唯赞RNA快速提取试剂盒说明书提取细胞总的RNA,使用超微量分光光度计测定所提取的RNA浓度,然后以1000 ng总量按照反转录试剂盒说明书将RNA逆转录为cDNA,-80 ℃保存。荧光定量PCR仪进行实时荧光定量PCR,以 β-actin为内参,2-ΔΔCt法分析各目的基因mRNA相对表达水平。
1.2.9Si-RNA干预将装有干粉状态的Si-RNA的EP管瞬时离心后加入适量的的DEPC水(无菌)溶解Si-RNA ,使其浓度为20 μmol/L,根据需要分装并存储至-20 ℃冰箱储存。首先,取无菌1.5 mL EP管3个分别标记Si-NC 、Si-YAP1、Si-ACSL4,每管加入100 μL无血清培养基和6 μL的lipofectamine2000并轻轻摇匀配制成a液体,静置3 min。再取无菌1.5 mL EP管3个分别标记Si-NC 、Si-YAP1、Si-ACSL4,每管加入100 μL无血清培养基和8 μL的Si-RNA混匀配制成b液体,室温下放置5 min。将配好的a液与b液体相混合,室温放置20 min形成混合物。在实验组中每组加入100 μL上述混合物后置于细胞培养箱继续培养24 h,干预后的细胞可用于提取蛋白或提取RNA等。

2 结 果
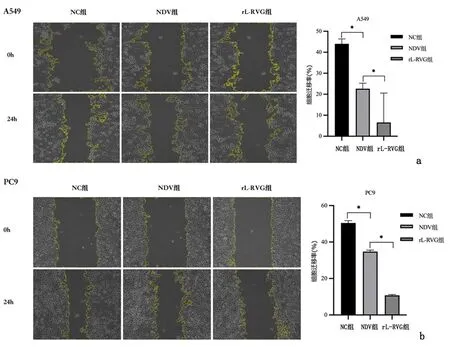
2.1 重组新城疫病毒rL-RVG感染后对肺腺癌细胞生长、迁移及侵袭能力及细胞形态的影响采用CCK-8法检测细胞活性,病毒感染时间设定为24 h时,不同MOI值的rL-RVG病毒感染A549、PC9及BEAS-2B细胞,A549及PC9细胞存活率在rL-RVG的MOI=5时与MOI为0、1、3相比差异均有统计学意义(P<0.01),与MOI=10相比较差异无统计学意义(P>0.05),通过对BEAS-2B细胞生存率分析可知,MOI=5时对正常组织细胞不存在毒害作用,细胞存活率97.67%,故选择MOI=5作为后续实验病毒感染复数。MOI=5时,NDV及 rL-RVG 对肺腺癌细胞活力抑制作用随时间增加而增强,且rL-RVG的抑制作用更强 (P<0.01),见图1。细胞划痕结果提示rL-RVG组细胞迁移率明显低于NC组 (P<0.01),见图2。Transwell实验结果也证明与NC组相比,NDV及均抑制肺腺癌细胞侵袭能力,且rL-RVG抑制作用更强(P<0.01),见图3。在光学显微镜下观察细胞状态,我们发现,NC组细胞数量增加,胞体丰满,形态良好,而NDV组和rL-RVG组的细胞数量减少,细胞出现皱缩形态不佳,其中rL-RVG组变化更为明显,见图4。上述结果显示,NDV及rL-RVG均能抑制肺腺癌细胞A549、PC9的生长、迁移及侵袭,且rL-RVG对肿瘤细胞的抑制作用较NDV更强。

a:A549细胞;b:PC9细胞
a:A549细胞迁移及数目统计分析;b:PC9细胞迁移及数目统计分析
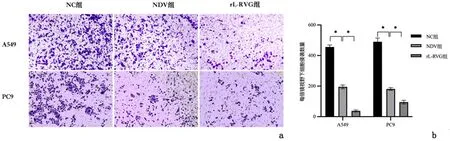
a:各组细胞侵袭情况(结晶紫染色 ×200);b:各组细胞侵袭数量统计分析
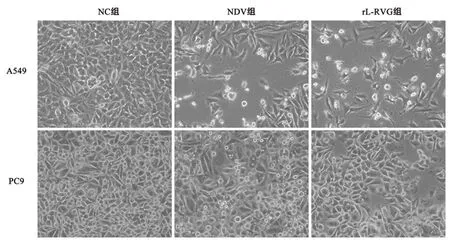
图4 光镜下观察各组细胞形态变化
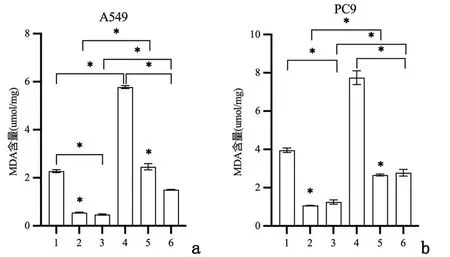
2.2重组新城疫病毒rL-RVG感染肺腺癌细胞后ROS及MDA水平变化流式细胞术检测铁依赖性的ROS含量,图5提示在A549和PC9细胞中,NDV、rL-RVG和铁死亡诱导剂Erastin均能增加ROS含量,其中rL-RVG作用强于Erastin;rL-RVG+Erastin组ROS含量大于rL-RVG组和Erastin组。相反,铁死亡抑制剂NAC可以减少ROS产生,rL-RVG可以挽救NAC造成的ROS减少。同时,ROS荧光强度测定也出现类似结果,见图6。用酶标仪法检测细胞内MDA含量,结果提示经过NDV、rL-RVG和Erastin处理的细胞中MDA含量明显增加(P<0.01 ),且rL-RVG组MDA含量最高;NAC可以减少MDA产生,rL-RVG可以一定程度上挽救NAC造成的MDA减少(P<0.01),见图7。
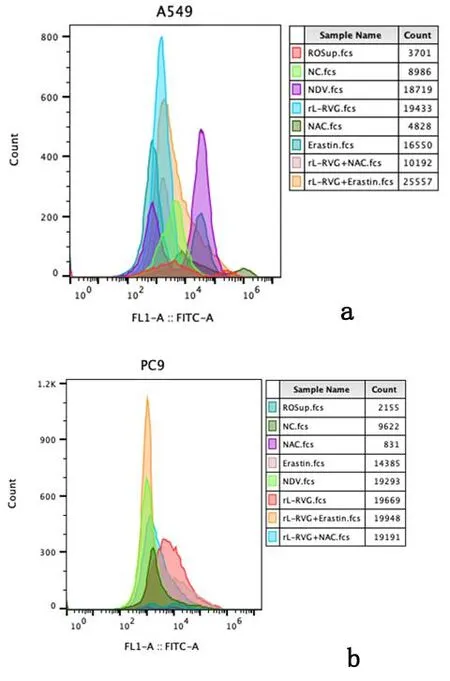
a:A549细胞;b:PC9细胞

a:A549细胞内ROS荧光强度及统计分析情况;b:PC9细胞内ROS荧光强度及统计分析情况
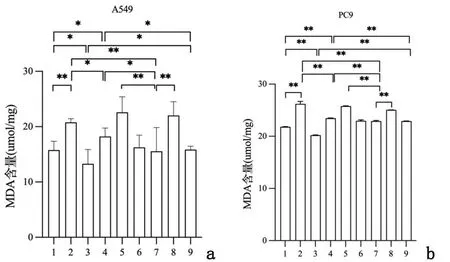
1:NC组;2:rL-RVG组;3:NAC组;4:NDV组;5:rL-RVG+Erastin组;6:rL-RVG+NAC组;7:Erastin组;8:NDV+Erastin组;9:NDV+NAC组
2.3重组新城疫病毒rL-RVG感染后铁死亡相关分子p53、YAP1、ACSL4的表达水平为进一步探究p53-YAP1-ACSL4通路在铁死亡中的作用,通过Western blot 证明rL-RVG感染后肺腺癌细胞P53、ACSL4和YAP1蛋白表达均上调(P<0.01),见图8。Erastin促进P53、YAP1、ACSL4表达,见图9,NAC抑制P53、YAP1、ACSL4表达且rL-RVG可以一定程度地减弱NAC的抑制效果,见图10。接着,利用Si-RNA分别对细胞中的ACSL4、YAP1进行敲减,通过实时荧光定量PCR检测目的基因mRNA的相对表达量,与Si-NC组相比,Si-ACSL4组中mRNA表达量大幅下降(P<0.01),证实ACSL4敲减成功。Si-ACSL4+rL-RVG组mRNA水平较Si-ACSL4组增高(P<0.01),但仍比Si-NC组要低(P<0.01),见表1。Western blot说明Si-ACSL4组中ACSL4蛋白表达水平较Si-NC组降低(P<0.01),再次证明敲减ACSL4是成功的。相应的,rL-RVG可以提高Si-ACSL4组中ACSL4的蛋白表达水平(P<0.01),但仍比Si-NC组中低(P<0.01),见图11。上述结果提示rL-RVG可以上调ACSL4的表达。同样地,我们用Si-RNA敲减YAP1,结果说明rL-RVG可以诱导肺腺癌细胞中YAP1在转录水平表达量上调,见表2,敲减YAP1的同时可以降低ACSL4的蛋白表达量,rL-RVG可以同时上调YAP1和ACSL4的蛋白表达水平,见图12。
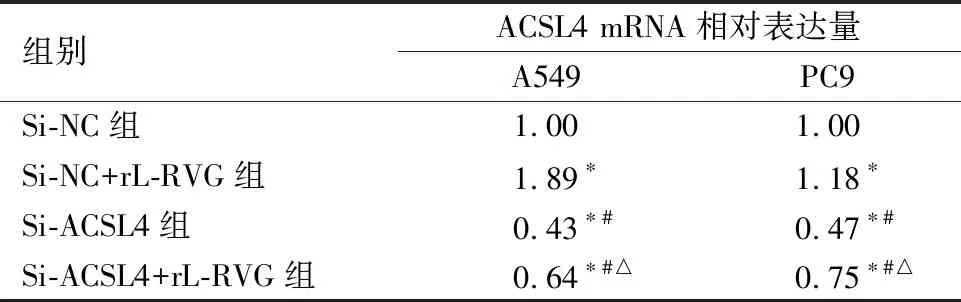
表1 Si-RNA干预后ACSL4的mRNA相对表达量

表2 Si-RNA干预后YAP1的mRNA相对表达量
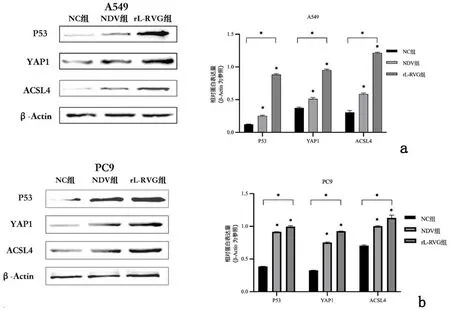
a:A549细胞;b:PC9细胞

1:NC组;2:NC+NAC组;3:rL-RVG+NAC组;4:rL-RVG组

1:Si-NC组;2:Si-ACSL4组;3:Si-ACSL4+rL-RVG组;4:Si-NC+rL-RVG组
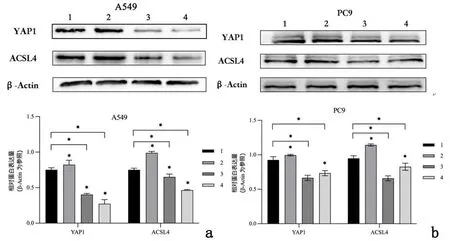
1:Si-NC组;2:Si-NC+rL-RVG组;3:Si-YAP1+rL-RVG组;4:Si-YAP1组
2.4敲减YAP1及ACSL4后对ROS及MDA水平的影响通过流式细胞术检测敲减YAP1及ACSL4后细胞中ROS积聚水平,与Si-NC组相比,Si-YAP1和Si-ACSL4组ROS水平均下降,rL-RVG可以在一定程度上挽救敲减YAP1及ACSL4所导致的ROS的减少(P<0.01),但较Si-NC组ROS水平仍显著下降(P<0.01),见图13。同样的,MDA含量的变化和上述结果相符合,见图14。
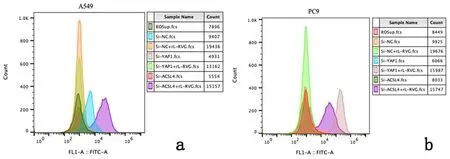
a:A549细胞;b:PC9细胞
1:Si-NC组;2:Si-YAP1组;3:Si-ACSL4组;4:Si-NC+rL-RVG组;5:Si-YAP1+rL-RVG组;6:Si-ACSL4+rL-RVG组
3 讨 论
铁死亡(Ferroptosis)作为一种新型的调节性细胞死亡(regulated cell death, RCD),与细胞凋亡、坏死和自噬等传统的RCD有着本质区别,主要涉及铁稳态、脂质代谢和谷胱甘肽代谢的改变[9]。铁死亡的本质是ROS积累诱导脂质过氧化,并经铁离子催化后发生代谢异常,脂质不断积累,破坏细胞内氧化还原稳态,攻击生物大分子,从而引起细胞死亡[10]。由于强烈的膜脂质过氧化和氧化应激的发生而导致质膜选择性通透性丧失[11],造成的线粒体嵴减少或消失,线粒体外膜破裂和线粒体膜浓缩是铁死亡的主要细胞学变化特征[12]。目前,人们已经对铁死亡进行深入探索,并且已经证实它与多种疾病的发生发展相关,其中与肿瘤之间的关系密切[13],多种信号通路也被发现参与铁死亡调节[14],因此铁死亡成为近年来肿瘤领域的研究热点之一。其中Hippo途径不仅能够有效地调节和整合上游信号而且还能够利用转录共激活因子YAP来指导细胞基因转录与生物学行为,对于细胞的生长发育、存活、增殖、迁移,组织器官大小的调节和肿瘤的形成和发展都有着重要的影响[15]。最近的一项研究表明,YAP1可以促进铁死亡[16]。
铁死亡的发生依赖于Hippo通路的活性,抑制Hippo通路或者激活YAP均可以促进铁死亡[17]。此外,p53和YAP1均在细胞周期和铁死亡中发挥调控作用,YAP1可直接结合TP53基因启动子上调p53表达,导致肝细胞癌化疗过程中的凋亡。反过来,P53可以与YAP1启动子结合上调其表达,建立一个正反馈回路[18]。相关研究表明p53-YAP1轴在调节结肠癌铁死亡中发挥了一种新的肿瘤抑制作用[19]。由于YAP1靶向多个铁死亡调控因子包括ACSL4[20],所以我们推测p53-YAP1-ACSL4轴在诱导铁死亡的发生过程中起到重要作用。
溶瘤病毒具备高度的特异性,能选择性地感染、大量复制并选择性杀伤肿瘤细胞。除了直接和局部的抗肿瘤活性外,还可以诱导有效、全身、持久的抗肿瘤免疫[21]。溶瘤病毒治疗肿瘤是安全且有效的[22]。新城疫病毒(new castle disease virus, NDV)是一种具有溶瘤作用的鸡瘟溶瘤病毒,对人类正常细胞无致病性。而安全、高免疫活性、稳定表达狂犬疫苗糖蛋白的重组新城疫病毒rL-RVG具有更强的扩散性,其抗肿瘤作用远远强于NDV[23]。但目前尚未有相关文献报道rL-RVG对肿瘤细胞铁死亡的影响。故本研究旨在探索rL-RVG是否能诱导肺腺癌细胞铁死亡,并进一步探索其相关机制。
在这项研究中,我们首先使用了两种病毒rL-RVG和NDV对肺腺癌 A549和PC9 进行感染,结果发现NDV和 rL-RVG都有抑制肺腺癌细胞的增殖、迁移和侵袭的效应,而rL-RVG的效力比NDV更明显。其次我们发现rL-RVG可以与Erastin相媲美,在抑制肺腺癌细胞增殖过程中可能发挥类似铁死亡诱导剂作用,促进细胞内ROS积聚,诱导肿瘤细胞发生铁死亡。进一步研究其相关机制后发现rL-RVG感染肺腺癌细胞后p53、YAP1及ACSL4的蛋白表达量均增加,同时Si-RNA的实验结果也提示YAP1及ACSL4参与调控铁死亡的发生,且ACSL4是YAP1的下游靶点。这些发现共同提示重组新城疫病毒rL-RVG可以通过激活p53-YAP1-ACSL4通路诱导人肺腺癌细胞铁死亡,这可能为临床治疗肺腺癌提供新的思路,使rL-RVG有望成为有吸引力的抗癌剂。
猜你喜欢
核科学与工程(2022年3期)2022-10-18 01:25:14
广州化工(2020年6期)2020-04-18 03:30:20
现代矿业(2018年9期)2018-10-16 09:37:02
中成药(2018年7期)2018-08-04 06:04:18
湖南畜牧兽医(2016年1期)2016-06-05 08:37:51
西南军医(2016年3期)2016-01-23 02:17:47
兽医导刊(2015年7期)2016-01-04 11:59:56
中国当代医药(2015年17期)2015-03-01 02:03:36
天然气与石油(2014年2期)2014-07-16 11:01:18
西南军医(2014年5期)2014-04-25 07:42:32
