
羊肉中呈膻物质的形成机制及降膻方式的研究进展
2024-05-08 03:00:54杨东松赵文秀罗玉龙罗瑞明侯艳茹
食品科学 2024年8期
杨东松,高 爽,袁 红,赵文秀,罗玉龙,罗瑞明,侯艳茹,*
(1.宁夏大学食品科学与工程学院,宁夏 银川 750021;2.宁夏大学动物科技学院,宁夏 银川 750021)
羊肉含有丰富的蛋白质、维生素和铁、锌、硒等微量矿物质元素,同时具有肉质细嫩、易消化、低脂肪和低胆固醇等优点,深受广大消费者的喜爱[1-2]。2022年,我国猪牛羊禽肉总产量高达9 227万 t,其中羊肉产量为525万 t,位居世界第一,但在全国猪牛羊禽肉总产量中仅占约5.7%[3-4]。同样,全世界范围内羊肉产量也仅占世界肉类总产量的4.5%,远低于其他肉类[5]。羊肉产量低与消费量不足密切相关,而羊肉本身具有的物种风味是导致羊肉消费量不足的主要原因之一,并已成为影响消费者可接受程度和制约羊肉开发的限制性因素。Wasserman等[6]研究发现,评定人员难以辨别分离脂肪的羊肉和其他肉类,将羔羊脂肪加入到犊牛肉中,评定人员将犊牛肉误判为羔羊肉的概率显著增加,进一步表明羊肉的物种风味是区别于其他畜禽肉类风味的关键风味特征。国外学者将羊肉这种不良的风味特征描述为“动物味”“腐败味”“金属味”“谷仓味”“粪臭味”“樟脑味”等[7],而我国普遍用“膻味”来描述羊肉的特征性风味。
膻味是由各种挥发性组分相互混杂,综合作用下而形成不良风味的总和,受到众多因素的影响和控制,具有构成复杂、来源广泛、含量极微、效果显著和消减难度大等特点,故探明羊肉膻味的来源、物质组成及形成机制已成为羊肉品质提升研究的热点,如何降低羊肉的膻味强度也成为羊肉开发的关键共性技术难题之一。因此,了解羊肉膻味的来源及形成机制,探究羊肉膻味强度的影响因素,总结当前的降膻方式,对于扩大羊肉及其制品的消费市场,促进养羊产业提质增效,推动羊肉质量和产量的协同发展具有重要的现实意义。
1 羊肉呈膻物质的来源及其形成机制
羊肉膻味并非由单一物质形成,而是由多种化合物相互作用的结果,其组成和比例的不同会造成膻味强度的差异。针对羊肉膻味,近些年国内外学者进行大量的研究探索羊肉膻味的来源和物质组成。目前,已报道的呈膻物质有4-烷基支链脂肪酸、硬脂酸、吲哚类化合物、酚类化合物和含硫化合物等[8-9],但各途径的形成机制尚未完全揭示,有待进一步深入探讨。
1.1 4-烷基支链脂肪酸
4-烷基支链脂肪酸中4-甲基辛酸(4-methyloctanoic acid,MOA)、4-乙基辛酸(4-ethyloctanoic acid,EOA)和4-甲基壬酸(4-methylnonanoic acid,MNA)是沉积在脂肪组织中的膻味脂肪酸,被认为是羊肉膻味的主要贡献者[10]。虽然4-烷基支链脂肪酸在肉羊不同组织部位的含量均不高,但具有较强的挥发性,其含量的变化将直接影响羊肉膻味的强弱[11]。在3 种特征支链脂肪酸中,MOA的浓度最高,且感知阈值低,对羊肉的膻味起主要作用,EOA对羊肉膻味的作用次之,MNA浓度最低,变化范围较大,对膻味的影响相对较小[12]。气味活性值(odor activity value,OAV)是指化合物浓度与其气味阈值的比值,通常情况下OAV≥1的组分被认为对总体风味有直接作用,而0.1≤OAV<1的组分被认为对总体风味具有修饰作用[13]。Watkins等[9]对绵羊脂肪组织中4-烷基支链脂肪酸的OAV进行测定,发现MOA和EOA的OAV分别为4.7~26.5和3.1~28.3,而MNA的OAV最低(0.0~0.1),这进一步表明MOA和EOA对羊肉膻味具有贡献作用。但值得注意的是,MNA和MOA、EOA之间呈较强的正相关性,其组合效应可能会对羊肉膻味产生叠加作用,即便MNA在肉羊组织的沉积量低,与MOA和EOA相互作用后可能会给羊肉膻味带来独特的变化[14-15]。此外,4-烷基支链脂肪酸在自然界以S构型和R构型两种形式存在,并且同一分子的对映异构体之间具有不同性质,所呈现的膻味强度也存在差异。Heinsman等[16]研究发现,肉羊组织中MOA主要以R构型的形式存在,其膻味强度是对映异构体S构型的5~10 倍。
关键呈膻物质4-烷基支链脂肪酸的合成路径主要有以下几种:1)饲粮中碳水化合物经过瘤胃微生物发酵后会产生大量的丙酸,丙酸是支链脂肪酸合成的重要前体物。丙酸通过瘤胃上皮进入门静脉,而后被运送到肝脏代谢。当丙酸的浓度超过肝脏的代谢阈值时,剩余的部分转化为丙二酰辅酶A和甲基丙二酰辅酶A[17],甲基丙二酰辅酶A与以丙酸为前体的乙酰辅酶A从头合成后可形成单甲基脂肪酸(MOA和MNA)[18]。在脂肪酸的从头合成过程中,因脂肪酸合成酶(fatty acid synthase,FASN)和乙酰辅酶A羧化酶(acetyl CoA carboxylase,ACC)的底物混杂特性也可产生支链脂肪酸。FASN的产物(直链脂肪酸和支链脂肪酸)取决于参与反应的底物,通常情况下FASN可利用丙二酰辅酶A合成直链脂肪酸,也可利用甲基丙二酰辅酶A代替丙二酰辅酶A生成甲基支链脂肪酸;ACC可催化丙二酰辅酶A的合成,利用丙酰辅酶A和丁酰辅酶A分别合成甲基丙二酰辅酶A和乙基丙二酰辅酶A,进而产生甲基支链脂肪酸和乙基支链脂肪酸[19]。乙基支链脂肪酸的生物合成途径可能是以丁酸替代丙酸作为前体,乙基丙二酰辅酶A替代甲基丙二酰辅酶A作为引物,其合成途径还有待进一步深入的研究[20]。另有研究认为,由于肉羊体内FASN负责转运的乙酰辅酶A/丙二酰辅酶A转移酶具有结构特异性,促使甲基支链更容易与中短链直链脂肪酸结合,导致组织中沉积的支链脂肪酸相较于其他物种更为丰富[21]。此外,丙二酰辅酶A脱羧酶是支链脂肪酸合成的关键酶,提高其活性可降低丙二酰辅酶A的利用率,增加羊体对甲基丙二酰辅酶A的利用率进而增加支链脂肪酸的合成量[22]。2)瘤胃微生物在脂肪酸的分解代谢中起着至关重要的作用,同样与组织中特殊支链脂肪酸的形成密切相关。饲料中的糖类物质经瘤胃微生物发酵可生成低级奇数碳羧酸,进而生成甲基丙二酰辅酶A为4-烷基支链脂肪酸的合成提供底物;也可利用支链氨基酸及相应的支链羧酸作为碳骨架底物合成4-烷基支链脂肪酸,经小肠吸收后沉积于肉羊脂肪组织中,对羊肉风味产生影响[23-24]。3)羊体组织中4-烷基支链脂肪酸的合成除了受瘤胃的发酵功能和微生物的分解作用影响外,还可能受羊体内脂肪酶的影响,促使长链脂肪酸降解,为4-烷基支链脂肪酸的合成提供条件。
1.2 3-甲基吲哚和4-甲基苯酚
在放牧条件下采食牧草的反刍动物畜产品具有明显的特征风味,也就是俗称的“田园风味”,其中包括含硫化合物、酚类化合物和吲哚类化合物等,而3-甲基吲哚(粪臭素)和4-甲基苯酚(对甲酚)是“田园风味”的主要贡献者[25]。
3-甲基吲哚是由反刍动物的瘤胃微生物对L-色氨酸的转氨基与脱羧作用而形成并沉积在羊肉的风味物质,对羊肉风味产生负面影响[26]。Young等[27]采用气相色谱-嗅觉法检测羊肉的气味化合物,发现3-甲基吲哚是唯一具有粪臭味的化合物,同时也发现3-甲基吲哚与放牧模式下饲养肉羊的特征性风味密切相关。肉羊在放牧饲养模式下进食大量牧草,当中的易发酵碳水化合物含量低,可溶性蛋白质含量较高,导致牧草中高浓度的蛋白质在瘤胃微生物和植物蛋白质水解协同作用下降解产生氨基酸和肽类,经降解产生的氨基酸和肽类含有大量的L-色氨酸,并在瘤胃微生物作用下通过转氨基和脱羧作用下生成3-甲基吲哚[28]。羊体内的3-甲基吲哚一部分被氧化成极性衍生物,与葡糖醛酸、谷胱甘肽或硫酸盐结合形成水溶性化合物随尿液排出体外[29];另一部分由肝脏代谢,若3-甲基吲哚的浓度超过肝脏的代谢阈值时,3-甲基吲哚会进入全身血液循环并沉积在羊脂中,进而对羊肉膻味产生影响(图1)[30-31]。

图1 色氨酸为前体物合成3-甲基吲哚的生物转化途径(BioRender.com绘制)Fig.1 Biotransformation pathway for synthesizing 3-methylindole using tryptophan as the precursor (created by BioRender.com)
4-甲基苯酚是瘤胃微生物对由酪氨酸的转氨基与脱羧作用产生的中间体——对羟基苯乙酸再次脱羧而形成的,具有较低的阈值,同样经血液循环沉积于脂肪组织中,对羊肉风味产生负面影响(图2)[25]。4-甲基苯酚的形成主要取决于反刍动物所进食的饲粮,并与放牧活动和肉品pH值密切相关[21]。有研究报道,在放牧条件下肉羊体内的4-甲基苯酚主要有两种来源。一种是饲粮中粗蛋白经瘤胃发酵产生酪氨酸,酪氨酸经过转氨基和脱羧作用而形成;另一种由瘤胃对饲粮中香豆酸酯和其他植物酚类物质的降解产生[32],其含量与放牧时间呈正相关关系,与肉中最终的pH值呈负相关关系[33]。
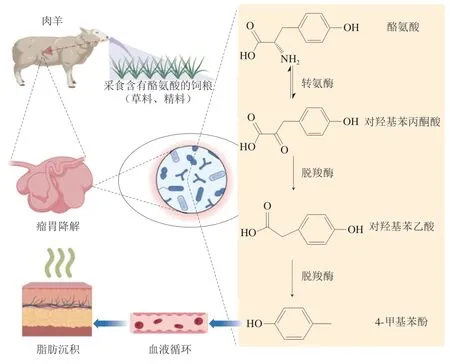
图2 酪氨酸为前体物合成4-甲基苯酚的生物转化途径(BioRender.com绘制)Fig.2 Biotransformation pathway for synthesizing 4-methylphenol using tyrosine as the precursor (created by BioRender.com)
1.3 硬脂酸
羊肉膻味除了来源于脂肪组织中的4-烷基支链脂肪酸,硬脂酸也是重要的呈膻物质之一,其经血液循环后沉积于羊脂中可使得羊肉的膻味增强,对羊肉风味产生影响。Sanudo等[34]研究发现,威尔士山羊和美利奴羊的膻味强度与硬脂酸的含量呈正相关关系。同时,脂肪酸的组成在很大程度上取决于瘤胃脂质降解和生物氢化程度,羊采食饲粮中的营养成分会影响瘤胃菌群作用下降解的游离脂肪酸,进而影响硬脂酸的合成[35]。经牧草饲养的肉羊其脂肪组织含有较高浓度的亚麻酸和硬脂酸,而以精料饲养的肉羊脂肪组织中含有较高浓度的油酸和亚油酸[36]。有研究指出,油酸、亚油酸和亚麻酸是羊肉中重要的风味物质前体物[37],而三者与硬脂酸的合成也密切相关。在放牧条件下饲喂牧草的肉羊生物氢化的主要脂肪酸底物是以糖脂和磷脂为主的亚麻酸,亚麻酸的氢化过程一般是先异构化形成顺-9,反-11,顺-15-18:3,再氢化为非共轭的反-11,顺-15-18:2,进而氢化形成硬脂酸;以精料饲养的肉羊生物氢化的主要脂肪酸底物是以亚油酸形成的三酰基甘油,其氢化过程由亚油酸转化为顺-9,反-11-18:2,然后进一步氢化为反-11-18:1,最后合成硬脂酸[38-39](图3);油酸在瘤胃蛋白分解梭状芽孢杆菌(Clostridium proteoclasticum)的作用下可转化为反-11-18:1,最终经还原酶氢化为硬脂酸[40]。由此可见,脂肪酸代谢产物的变化也与瘤胃细菌密切相关。溶纤维丁酸弧菌(Butyrivibrio fibrisolvens)可氢化多不饱和脂肪酸,参与形成共轭亚油酸和亚油酸的过程,蛋白分解梭状芽孢杆菌参与油酸、亚油酸转化为硬脂酸的过程[41];蛋白溶解梭菌(Butyrivibrio proteoclasticus)的数量与瘤胃中硬脂酸的生成量呈负相关关系[42]。
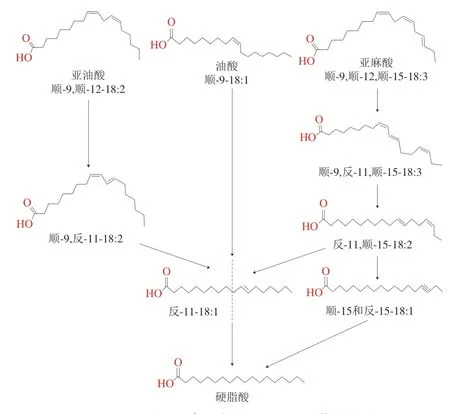
图3 亚油酸和亚麻酸合成硬脂酸的生物氢化途径Fig.3 Biohydrogenation pathway for the synthesis of stearic acid from linoleic acid and linolenic acid in the rumen of ruminants
1.4 氨基酸
4-烷基支链脂肪酸除了来源于脂肪酸的从头合成,还来源于支链氨基酸的降解[15],因此支链氨基酸的含量也会影响羊肉的膻味强度。支链氨基酸可为瘤胃细菌合成支链脂肪酸提供前体物(图4),如缬氨酸、亮氨酸和异亮氨酸可在支链氨基酸转移酶的作用下生成相对应的支链羧酸(2-甲基丙酸、2-甲基丁酸和3-甲基丁酸),通过支链α-酮酸脱氢酶的催化作用生成支链脂酰辅酶A,再经合成酶的作用形成低分子质量的挥发性支链脂肪酸,为呈膻物质4-烷基支链脂肪酸的合成提供支链碳架结构[43]。同时,丙酰辅酶A是缬氨酸和异亮氨酸的降解产物,而丙酰辅酶A经ACC羧基化可合成甲基丙二酰辅酶A,进而产生甲基支链脂肪酸,对羊肉膻味产生影响[44]。刘旺景等[45]利用代谢组学技术对不同膻味水平小尾寒羊肾周脂肪的代谢组学特征进行分析,进一步证实了4-烷基支链脂肪酸的合成与支链氨基酸的代谢通路密切相关。此外,芳香族氨基酸(酪氨酸和色氨酸)对“田园风味”化合物的形成也起到了重要作用,其中色氨酸是生成3-甲基吲哚的重要前体物,而酪氨酸是生成4-甲基苯酚的重要前体物[31]。含硫氨基酸(胱氨酸、半胱氨酸和蛋氨酸)在一定条件下也可转化为硫化物和硫醇,经瘤胃发酵后可导致挥发性含硫风味化合物和“田园风味”化合物在羊体内积累,加剧羊肉膻味[7]。
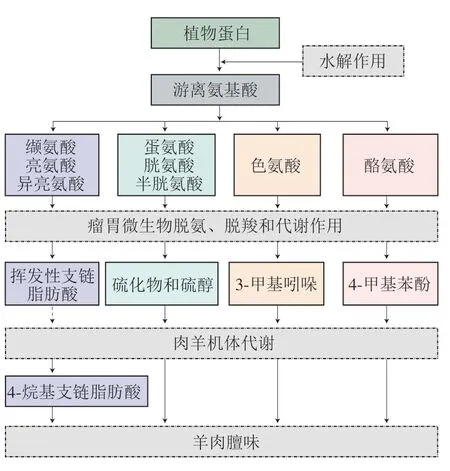
图4 以氨基酸为前体物形成羊肉膻味物质的合成路径(BioRender.com绘制)Fig.4 Synthetic pathway for the formation of mutton odor substances using amino acids as precursors (created by BioRender.com)
2 影响羊肉呈膻物质形成的因素
2.1 品种
品种受遗传基因的调控和影响,其基因差异性可能是由机体内的脂肪酸组成和代谢遗传控制所造成的,是影响羊肉膻味强度的主要因素之一[46]。有研究报道,肉羊的基因差异性可能会导致基因表达、脂肪酶活性、去饱和或链延伸酶活性的不同,进而影响脂肪酸的组成和沉积量[47]。Momen等[48]也观察到,在相同的饲养条件下,3 种伊朗绵羊品种的尾部脂肪和内脏脂肪的脂肪酸合成潜力、组成和沉积量均不同,其中棕榈酸、亚麻酸、共轭亚油酸和硬脂酸差异显著。说明不同品种的肉羊其机体对风味化合物的代谢能力存在差异,膻味脂肪酸含量可能也会有差别。Alves等[49]对达马拉肥尾羊、杜泊羊和美利奴羊尾部脂肪中甲基支链脂肪酸的含量进行了比较,观察到达马拉肥尾羊尾部脂肪中甲基支链脂肪酸的含量高于杜泊羊和美利奴羊。丁艳艳等[50]对小尾寒羊、滩羊、同羊和蒙古羊皮下脂肪组织的4-烷基支链脂肪酸进行了测定,发现4 个品种MOA和MNA的含量存在差异,其中MOA的含量按照小尾寒羊、滩羊、同羊、蒙古羊的顺序依次减少,MNA的含量按照同羊、滩羊、小尾寒羊、蒙古羊的顺序依次减少。Salvatore等[51]对5 种基因型杂交羊背部皮下脂肪的4-烷基支链脂肪酸进行测定,结果发现无角多赛特与美利奴羊的杂交羊中MOA和MNA的含量高于其他4 种基因型的杂交羊,推测可能是无角多赛特与美利奴羊的杂交羊存在特定的基因影响了羊体中MOA和MNA水平,进一步表明膻味水平存在一定的遗传效应。同时,纯种羊与低膻味的肉羊品种进行杂交,其体内膻味物质的含量也存在差异。张志超等[52]对相同舍饲条件和年龄的澳寒羊(澳洲白羊与小尾寒羊的杂交后代)和小尾寒羊皮下脂肪中4-烷基支链脂肪酸的含量进行测定分析,结果发现澳寒羊中MOA和EOA的含量低于小尾寒羊,表明羊肉的膻味强度可通过遗传选择和育种技术进行改善。
2.2 年龄和性别
羊机体中脂肪酸含量、组成和膻味物质的沉积量随年龄而变化,羊肉的膻味强度也随着羊年龄的增长不断增强[53]。研究发现,年龄的增长与羊体内的4-烷基支链脂肪酸、十七烷酸和硬脂酸等膻味脂肪酸的含量呈正相关关系[54-55]。欧慧敏等[11]对呼伦贝尔羊不同生长阶段背部皮下脂肪中4-烷基支链脂肪酸的含量进行测定,结果显示MOA、MNA和EOA的含量均按育肥后期(6 月龄)、育肥中期(5 月龄)、断奶期(2.5 月龄)和新生期(1~2 d)顺序依次减少,其中MOA和MNA的含量在育肥中期至育肥后期的增速最快,EOA的含量在断奶期至育肥中期的增速最快,表明年龄因素对羊肉膻味的形成和沉积具有重要影响。Salvatore等[51]探究了年龄对澳大利亚绵羊的脂肪组织中MOA和MNA含量的影响,发现22 月龄绵羊MOA的含量低于8 月龄的羔羊,与其他研究结果相悖。由此可见,羊机体内膻味物质的含量不能作为羊年龄的判断依据,屠宰日期和饲粮类型等其他因素均可影响呈膻物质的含量。但有众多研究表明,羊年龄与羊机体内呈膻物质的沉积量呈正相关关系。因此,选择适宜年龄的肉羊进行屠宰对羊肉膻味的改善仍具有重要的意义。
相关研究发现,性别也是影响羊肉膻味物质含量的重要因素。随着雄性羔羊年龄的增长,睾酮激素也会随之提高,公羊与母羊、去势羊之间的膻味强度的差异显著,而母羊与去势羊之间的膻味强度差异不显著[56]。由于去势可增加肉羊机体血液中卵泡刺激素和促黄体生成素水平,降低促性腺激素和雄性激素睾酮浓度,进而减轻了羊肉膻味[57]。Gkarane等[58]使用定量描述分析和感官评价得出,公羊与去势羊相比,公羊肉的不良特征风味得分更高,推测与公羊的睾酮水平有关,睾酮水平可在一定程度上影响蛋白质和脂质的代谢,导致其不良特征风味的得分高于去势羊。Sutherland等[59]研究发现,在12 周屠宰的公羊和去势羊中鉴定出的总脂肪酸几乎相同,MOA和MNA无显著差别;而在30 周屠宰的公羊总脂肪酸比去势羊高1.5 倍,MOA和MNA也分别增加了13 倍和4 倍,并且MOA和MNA在12 周屠宰的去势羊样本中最低,在30 周屠宰的公羊样本中最高。由此可见,公羊的膻味强度高于母羊和去势羊,成年羊的膻味强度高于羔羊。年龄和性别效应及交互作用都会导致肉羊体内脂肪酸组成、挥发性风味化合物以及呈膻物质含量的差异,特别是成年公羊体内的4-烷基支链脂肪酸高于同龄去势羊或未成熟的羔羊,这可能与公羊体内的高浓度的睾酮激素及代谢水平密切相关。
2.3 组织部位
由于呈膻物质4-烷基支链脂肪酸合成的组织特异性,导致肉羊不同部位的膻味脂肪酸的含量存在差异。Brennand等[60]对肉羊不同组织部位的4-烷基支链脂肪酸进行定量分析,结果显示皮下脂肪组织样本(臀部、胸部和肩部)4-烷基支链脂肪酸的含量整体显著高于肾周脂肪组织样本和肌肉组织样本,其中臀部和肩部皮下脂肪MOA的含量高于胸部皮下脂肪,臀部和胸部皮下脂肪MNA的含量高于肩部皮下脂肪,而肩部皮下脂肪EOA的含量高于胸部和臀部皮下脂肪。Berthelot等[61]研究表明,在饲粮中添加丙酸,肉羊皮下脂肪组织4-烷基支链脂肪酸的增幅大于内脏脂肪,推测的原因是皮下脂肪组织对丙酸的利用能力更强,导致其皮下脂肪组织的4-烷基支链脂肪酸的沉积量更高。刘旺景等[62]在日粮中添加沙葱及其提取物,并对小尾寒羊不同部位的4-烷基支链脂肪酸的沉积量进行比较和分析,发现MOA、MNA和EOA的沉积量按照尾部脂肪、背部皮下脂肪、大网膜脂肪及肾周脂肪的顺序依次减少。综上所述,羊体不同组织中呈膻物质的合成潜力和沉积量具有部位差异性,其体内膻味脂肪酸在皮下脂肪组织中的沉积量普遍高于其他脂肪组织,可能是由于皮下脂肪组织易受饲粮的影响,其脂肪酸合成比例高于其他脂肪组织,或者是由2 种脂肪酸的沉积方式(脂肪酸的从头合成和外源脂肪酸的摄取)在脂肪组织之间的差异而造成上述的结果[63]。
2.4 营养及饲养体系
羊肉呈膻物质的合成、沉积与羊机体对脂肪的利用密切相关,而不同的营养(饲粮中能量水平、蛋白质、矿物质、维生素和不饱和脂肪酸的含量)及饲养体系(放牧和舍饲)都会导致肉羊机体内脂肪酸的含量和组成存在差异,进而对羊肉膻味产生影响[64]。饲粮经反刍动物的瘤胃吸收后,通过瘤胃的微生物作用降解生成挥发性脂肪酸、氨基酸及氨类等物质,随后被微生物利用合成其他产物并经机体代谢和吸收沉积于组织中,进而产生不同的挥发性风味物质[65]。以谷物为基础的饲粮会导致羊肉中一些支链脂肪酸、醛和内酯的积累,而以牧草为基础的饲粮会导致羊肉中的酚类、萜烯、吲哚和硫化物的积累。有研究报道,与采食牧草的肉羊相比,饲喂精料的肉羊膻味更加强烈[15],其原因可能是以谷物为基础饲喂的肉羊中含有较高浓度的单不饱和脂肪酸和多不饱和脂肪酸,在贮存过程中易受脂质氧化和蛋白质氧化的影响,导致其脂肪组织中支链脂肪酸的增加;也可能是高能量的谷物饲粮在瘤胃中产生了更多的丙酸和丁酸,使贮存在脂肪组织中奇数碳链脂肪酸和支链脂肪酸的含量增加[7]。但也有报道指出,肉羊进食混合苜蓿、盐灌木等草料其羊肉膻味反而会更强烈[66]。由于在放牧模式下,肉羊采食不同的草料类型如白三叶草、紫花苜蓿、羽衣甘蓝和油菜等,其肉品风味也会有所不同,其中的营养物质(如蛋白质和碳水化合物的含量)及特定的化学组分(如羽衣甘蓝和油菜中含有的硫代葡萄糖苷)均会对羊肉膻味造成不同程度的影响[67-68]。由此可见,肉羊在天然放牧时的行为习性和采食远比舍饲复杂,其风味具有更大的可变性。不同地域条件、刈割季节、成熟度和施肥条件的同一类型牧草都会对羊肉风味产生不同的影响,如嫩草中含有高浓度的可溶性蛋白质及氮肥的超量使用,都会产生更多的呈膻物质,对羊肉风味造成负面影响。
饲粮的物理特征(体积和密度)的差异会影响肉羊组织中支链脂肪酸的含量。有研究报道,饲粮经过碾压、研磨等加工后,能够降低瘤胃发酵的乙酸盐与丙酸盐的比率,而高浓度的丙酸盐可导致肉羊脂肪组织中亚麻酸和支链脂肪酸的浓度升高[69-70],对羊肉风味产生不良影响。另外,饲喂期的长短也会对羊肉膻味造成影响。De Brito等[71]比较了不同类型的饲粮对羊肉风味的影响,发现不同饲粮之间没有明显的感官差异,但在他们的实验中,饲喂期只有49 d,这可能是由于饲喂时间较短使得挥发性风味化合物还未在羊脂中蓄积。Rousset-Akrim等[72]研究发现,与饲喂90 d屠宰的羔羊相比,饲喂215 d的肉羊风味更浓重,这进一步表明饲喂期的增长会导致羊肉的风味强度也增强。
3 羊肉降膻方式
3.1 遗传降膻法
营养及饲养体系在指导羊肉生产中发挥直接作用,但肉羊的遗传潜力在一定程度上也决定了其生产的肉品质量。品种、性别和年龄等遗传因素均可影响脂肪酸的组成和沉积量,对羊肉的膻味强度产生影响。Kaffarnik等[12]研究显示,美利奴羊、法国伊尔羊与美利奴羊杂交羊和新西兰羊杂交羊皮下脂肪中4-烷基支链脂肪酸的含量存在差异,其中美利奴羊4-烷基支链脂肪酸的含量最高,法国伊尔羊与美利奴羊杂交羊和新西兰羊杂交羊的含量较低。丁武[73]对波尔山羊与关中奶山羊的杂交后代羊进行研究,结果发现获得的杂交后代羊其肌肉嫩度、肉色及膻味均到改善,进一步表明选育膻味强度较低的品种进行杂交繁育,是降低羊肉膻味强度的手段之一。Deighton等[74]研究发现,绵羊瘤胃中输注外源性的粪臭素会导致肌间脂肪粪臭素的沉积水平存在显著差异,存在差异的原因可能与粪臭素相关的代谢基因有关,故筛选羊肉膻味遗传因子有助于利用遗传调控方式选择和繁育低粪臭素的肉羊品种。综上,膻味物质的含量可通过遗传学手段进行调控,发挥杂交优势,选育膻味强度低的品种是改善羊肉膻味长效而又稳定的措施之一。但目前膻味化合物的遗传力数据鲜有报道。因此,在实际繁殖育种的过程中,可利用先进的基因测序技术进一步发掘羊肉膻味相关的遗传因子,探索膻味的遗传机制,确定膻味性状相关的遗传基因并解析遗传差异,以期建立良种繁育体系,从而达到降低膻味基数、选育低膻味水平品种的目的。
3.2 饲粮降膻法
通过改变饲粮中营养成分可调控羊肉呈膻物质的物质组成及含量,从而影响羊肉的膻味强度。刚虎军[75]在日粮中添加不同梯度的甘草,发现当甘草添加量占日粮质量的40%时可降低新疆多浪羊皮下脂肪组织中癸酸、辛酸、乙酸、MOA和EOA的含量,具有较好的降膻效果。这可能是由于甘草在弱碱性的环境下生长,在弱碱性条件下呈膻物质被减弱,也可能是甘草中的主要活性成分——甘草酸经瘤胃形成的代谢产物可降低膻味脂肪酸沉积的底物,进而减少呈膻物质的合成。另有研究表明,利用百合科葱属植物及其提取物的活性成分可阻滞瘤胃微生物的生物氢化作用,影响瘤胃微生物菌群的组成和脂肪酸的代谢能力,进而抑制4-烷基支链脂肪酸的合成相关酶及基因表达,从而降低4-烷基支链脂肪酸在羊体的沉积量。Liu Wangjing等[76]研究发现,日粮中添加沙葱黄酮类化合物可降低小尾寒羊背最长肌中硬脂酸、MOA等膻味物质的含量,有助于减轻羊肉膻味。丁赫[77]采用宏基因组学和代谢组学技术进一步证实了沙葱及其提取物可影响绵羊瘤胃微生物区系,改变瘤胃微生物菌群的组成,减少了4-烷基支链脂肪酸的沉积量,具有较好的降膻效果。此外,中草药因具有多功能性、无抗药性和毒副作用小等特点,有学者选取多种中草药材进行配伍,将其添加于饲粮中以减轻羊肉膻味。罗燕[78]选用苍术、肉桂、茴香、陈皮和山楂等药材按照比例进行配伍组成中草药添加剂,以粗品粉状的形式添加于日粮中,发现中草药添加剂在羊体内发挥协同作用后可降低癸酸、硬脂酸和总脂肪酸的含量,推测的原因是中草药添加剂可影响羊机体脂肪代谢的基因表达,降低FASN的活性,进而调控脂肪酸的组成和含量,以达到减轻羊肉膻味的目的。
相关研究指出,羊采食牧草在机体衍生出的3-甲基吲哚和4-甲基苯酚会引发支链脂肪酸结构的变化,加剧由支链脂肪酸引发的膻味[79],故通过饲粮中的一些活性成分抑制“田园风味”化合物的形成和积累对于降低羊肉的膻味强度具有重要意义。Girard等[80]研究发现,驴食草(Onobrychis viciifolia)中缩合单宁(condensed tannins,CTs)的含量与肉羊肾周脂肪中粪臭素的含量呈负相关关系。Schreurs等[81]采用不同类型的草料进行体外发酵发现,饲喂具柄线蕨(Leptochilus pedunculatus)形成粪臭素的浓度仅为饲喂白三叶草的7%,而饲喂地中海岩黄芪(Hedysarum coronarium)形成的粪臭素浓度为饲喂白三叶草的53%。这是由于地中海岩黄芪当中的单宁浓度相比于具柄线蕨较低,进一步研究表明CTs对粪臭素的抑制作用存在量效关系,其浓度越高,对粪臭素的抑制作用越显著。推测的原因是单宁可与蛋白质形成络合物,调控参与蛋白质降解的瘤胃微生物,降低色氨酸的浓度以抑制粪臭素的形成。另有研究表明,添加淀粉或葡萄糖以增加体外培养中易发酵碳水化合物的含量,可使由色氨酸衍生的粪臭素和吲哚的含量减少至原来的1/4~1/2,进一步表明调控色氨酸对粪臭素的转化率可减少粪臭素的含量[82]。同时,羊体组织中“田园风味”的形成与进食的饲粮及瘤胃对饲粮蛋白降解的能力密切相关,通过影响参与代谢的微生物(细菌、原生动物和真菌)及瘤胃中粪臭素的合成,可调控瘤胃对饲粮蛋白的降解,减少“田园风味”化合物在羊体的积累,同样是减少呈膻物质形成的途径之一[31]。
3.3 微生物降膻法
微生物降膻法是将一种或多种菌株制成发酵剂,接种到羊肉及其制品中并进行发酵处理,进而达到降低羊肉膻味的效果,因其具有安全、温和且降膻效果良好等特点,近年来已成为羊肉降膻技术的研究热点[83]。彭健斌等[84]筛出肉葡萄球菌(Staphylococcus carnosus)、植物乳杆菌(Lactobacillus plantarum)和戊糖片球菌(Pedicoccus pentosaceus)3 株发酵菌种,将其制成发酵剂对羊肉进行发酵处理,结果发现MOA、MNA和EOA 3 种膻味脂肪酸的降解率均可达到60%~80%,其中肉葡萄球菌对膻味脂肪酸的降解能力最强,戊糖片球菌次之,植物乳杆菌最弱。Holko等[85]选取嗜酸乳杆菌(L.acidophilus)和双歧杆菌(Bifidobacterium)制备益生菌混合发酵剂应用于羊肉香肠中,经过感官评价分析发现,该混合发酵剂可减轻羊肉香肠的膻味。此类降膻方法主要是利用微生物释放的脂肪酶对膻味脂肪酸的降解能力改变膻味物质的构型及其存在形式,如乳酸菌产生的脂肪酶对羊肉脂肪具有水解作用,菌体分解释放的蛋白酶和酯酶可在一定程度上降解膻味脂肪酸[86];另一方面,蛋白质经蛋白酶水解后产生的肽、氨基酸和脂肪分解产生的脂肪酸可以通过美拉德反应和氧化降解形成风味物质,降低呈膻物质的相对含量,从而达到既降低羊肉的膻味强度又不破坏其食用品质的目的[84]。但目前对降膻菌株的研究较为有限,发掘出更多具有降膻作用的菌株并探究其作用机理是微生物降膻技术未来需要进一步研究的重点。
3.4 物理化学降膻法
羊肉膻味主要来源于特征支链脂肪酸,具有一定的挥发性,采用高温处理方式结合真空急骤蒸发原理可使羊肉中的膻味脂肪酸得以挥发,进而达到减轻羊肉膻味的目的。另有研究表明,羊肉通过多阶段漂洗可减少总脂肪和游离脂肪酸的含量,降膻效果良好。顾仁勇等[87]采用漂洗法对羊肉进行脱膻处理,其工艺条件经优化后发现,温度40 ℃、pH 8.2、肉与水的比例1∶7、漂洗5 次的工艺组合能降低羊肉的膻味强度。Rhee等[88]将羊肉绞碎后采用pH值为8.2的清水、漂洗次数为4 次、漂洗周期为15 min、肉与水的比例为1∶4的组合条件对其进行降膻处理,结果发现该漂洗条件可显著降低羊肉中乙酸、辛酸、癸酸、MOA、MNA和EOA等膻味脂肪酸的含量。但该类方法受限于产品形态及体积,较适用于糜状产品,其他形态的产品呈膻物质的渗透和扩散效率低,降膻效果差[24]。综上,漂洗时间、漂洗温度和漂洗次数与羊肉的膻味强度呈负相关关系,产品体积与羊肉的膻味强度呈正相关关系。值得注意的是,此类方法的降膻效果不稳定,并且会破坏羊肉的风味和营养价值,降低羊肉的食用口感。
通过添加吸附剂和酯化反应的物质也可有效减轻羊肉的膻味。β-环糊精作为一种应用广泛的吸附剂,是由α-1,4-连接的吡喃葡萄糖亚基组成的环状低聚物,具有疏水分子形成包合物的能力,其分子对支链脂肪酸具有较高的亲和力,可与膻味脂肪酸结合,当中的有机物能被β-环糊精分子洞的疏水空腔所包埋形成非挥发性复合物,从而达到降膻的目的[89-90]。同时,依据羊肉呈膻物质的化学特性,利用特定的化学物质与羊肉膻味物质进行酯化还原反应形成芳香类化合物,同样是降低羊肉膻味的途径之一。例如醇类和酚类物质可与羧酸类膻味物质发生酯化反应生成有机酸酯、脂肪酸酯等物质,将膻味脂肪酸转化为酯类增香物质以降低膻味强度[91]。但该方法通常会受到相关食品法规的约束,需严格控制用量,若用量不当会导致羊肉产生其他的异味,并可能发生化学物质残留损害羊肉食用品质的情况。
3.5 食材降膻法
羊肉在进行预处理及加热过程中,通常会添加香辛料以达到降膻的效果,如大蒜中蒜氨酸可分解许多硫化物,能掩蔽羊肉在加热过程中释放的挥发性硫化物。八角、花椒、丁香等香辛料中富含的芥子苷类、酮类化合物,在羊肉加热过程挥发中具有浓烈的增香效果,可掩盖肉中的膻味[91],但羊肉经冷却贮藏后膻味又会呈现[92],说明利用香辛料进行降膻处理只能起到暂时性的掩蔽作用,呈膻物质没有消失或其化学成分的结构并未改变。王振东[93]选取胡萝卜、白萝卜、绿豆和板栗等传统烹饪羊肉所用食材,并将其制备成食材汁分别对羊肉进行腌制和煮制,结果发现食材汁可降低羊肉中亚麻酸和硬脂酸等膻味脂肪酸的含量,并随着食材汁添加量的增加,其降膻效果越显著。推测原因是食材汁中芳香性化合物对羊肉的膻味具有一定的掩盖作用,也可能是其空间结构可吸附呈膻物质,在一定程度上包埋了羊肉中的呈膻成分;另一方面,食材汁中的生物碱、有机醇、有机酸等成分可与呈膻物质发生化学反应,降低其挥发性,进而减轻了羊肉膻味。
4 结语
羊肉膻味是影响消费者购买的限制性因素,导致其消费总量远低于其他肉品的主要原因之一。4-烷基支链脂肪酸、3-甲基吲哚、4-甲基苯酚、硬脂酸和氨基酸等呈膻物质的含量均会对羊肉的膻味强度造成影响,从其形成机制总结和分析可以得出:1)饲粮中的碳水化合物经瘤胃微生物发酵后可产生大量的丙酸,添加丙酸能够显著增加肉羊不同组织部位4-烷基支链脂肪酸的沉积量。同时,4-烷基支链脂肪酸的合成与瘤胃发酵活跃性呈显著性关系,故减少瘤胃中丙酸的生成量,抑制4-烷基支链脂肪酸合成的关键酶活性如丙二酰辅酶A脱羧酶,可降低甲基丙二酰辅酶A或乙基丙二酰辅酶A参与合成支链脂肪酸的几率,进而调控4-烷基支链脂肪酸的合成。2)3-甲基吲哚和4-甲基苯酚主要来源于瘤胃对蛋白质的发酵。肉羊采食不同的草料会导致其体内的3-甲基吲哚和4-甲基苯酚的含量存在差异,这些差异与草料中的粗蛋白的浓度以及瘤胃对蛋白质的降解能力密切相关,故降低肉羊体内“田园风味”化合物的积累时,利用瘤胃对蛋白质的降解能力十分重要。3)亚麻酸、亚油酸、油酸、支链氨基酸及支链羧酸是呈膻物质的重要前体物。因此,改变它们的供应模式可影响呈膻物质的合成,进而调控呈膻物质在羊体的沉积量,同样是降低羊肉膻味的途径之一。
近年来国内外对羊肉降膻技术的研究已不断深入并取得新的进展,涉及物理、化学以及生物等领域且方法多样,但目前改善羊肉膻味最有前景的途径还是营养调控和遗传育种。在这两种途径中,营养调控对羊肉膻味改善的效果更为直观,是所有降膻方式中最高效、便捷和经济的方式,而遗传育种方式则更为长效且稳定。但目前对羊肉膻味基因表达的研究成果较少,对膻味相关基因的挖掘还有很大的空白,未来需进一步探究呈膻物质的候选基因、代谢通路和分子调控机制。同时,当前的降膻方式大多采用单一的降膻技术,且无法被广泛应用于工业化生产中,具有较大的局限性。因此,亟待从分子结构层面对呈膻物质进行更全面和深入的研究,并以此为理论依据结合多种降膻技术,使其互补并发挥协同增效作用,以获得经济高效、安全温和和应用广泛的新型降膻技术。在实现膻味消减、提高风味的同时,最大限度保存羊肉的营养价值和风味口感,以期从根本上解决羊肉的膻味难题,推动羊肉消费市场的发展。
猜你喜欢
百科知识(2023年1期)2023-05-30 10:48:04
饮食科学(2019年5期)2019-11-21 13:17:53
西南石油大学学报(自然科学版)(2018年6期)2018-12-26 01:00:18
饮食保健(2018年23期)2018-12-20 01:06:08
食品与健康(2015年3期)2015-09-10 07:22:44
实用手外科杂志(2015年2期)2015-08-28 09:50:56
饲料博览(2015年4期)2015-04-05 10:34:14
北京航空航天大学学报(2014年1期)2014-12-19 08:58:38
食品工业科技(2014年15期)2014-03-11 18:17:49
食品科学(2013年19期)2013-03-11 18:27:27
