
黄土丘陵区刺槐人工林林龄增加土壤微环境变化对凋落物分解的影响
2024-05-06 03:13:14张晓曦胡嘉伟刘凯旋王丽洁王羿人陈灵素董雨欣
生态学报 2024年7期
张晓曦,胡嘉伟,刘凯旋,王丽洁,胡 伟,田 爽,王羿人,陈灵素,董雨欣
1 延安大学 生命科学学院,延安 716100 2 中国科学院东北地理与农业生态研究所黑土区农业生态院重点实验室,哈尔滨 150081
黄土高原是我国生态环境最为脆弱的地区之一。20世纪中叶以来,当地陆续营造了大规模的人工林,在固持水土和修复受损生态环境等方面发挥了关键作用[1—2]。然而,刺槐Robiniapseudocacia等外来树种人工林大量消耗了当地土壤水分,导致黄土高原的造林面积已经接近水分承载力极限[3—4]。因此,如何保证现有林地长期稳定存在、并发挥其生态功能显得尤为重要。目前,黄土高原多种人工纯林均面临着不同程度的衰退风险[1—2]。除土壤水分损耗、林木和林下植被发育欠佳以及生物多样性偏低等限制人工林稳定性的常见因素外[5—6],植被对土壤的养分归还特性也是影响其长期稳定的重要因素之一[7]。凋落物分解是森林植被向土壤归还矿质元素、输入有机物和能量的核心环节,直接关系着林地土壤肥力维持、土壤结构的形成和碳库稳定[8—9]。因此,了解人工林植被恢复过程对凋落物分解的影响趋势及其潜在机制有助于从物质循环角度预测人工林稳定性的可能发展趋向,对维持人工林的可持续性具有重要的理论和实践意义。
大量研究指出,凋落物的分解受其自身理化特性和物种组成[9—11]、分解者的群落和功能特征[12—14]以及分解环境[15]的共同作用,且已就上述因素改变对凋落物分解的影响做了较为深入的探讨。然而,现有研究尚少关注植被改变、特别是人工林植被恢复过程中土壤微环境因素的变化及其对林地凋落物分解的影响[16—17]。与天然植被演替过程相比,人工纯林的土壤环境变化往往存在一定的特殊性。首先,单一树种对土壤养分的选择性吸收和归还可能导致凋落物分解环境中关键养分匮乏或失衡[1—2, 18]。其次,林地长期生长过程中,其郁闭度和地被物的改变显著影响与凋落物分解相关的光照、湿度和温度等物理环境[19—20]。更重要的是,植被生长、凋落物归还和林下环境的变化共同驱动了林地土壤中微生物的群落组成、结构及其潜在功能的改变[20—21]。鉴于土壤微生物在植物残体脱落后能够迅速侵入并在相当程度上参与凋落物的分解,其改变必然显著影响林地凋落物的分解过程[22]。然而,前述土壤微环境的变化通常是由建群树种特性塑造;且较之受到广泛关注的长期自然植被演替,相对短期的人工林植被恢复过程可能尚不足以引起土壤环境的急剧改变。因此,特定人工林的植被恢复过程中,林地土壤环境的具体改变如何影响林地凋落物的分解特征尚有待进一步研究加以阐明。
刺槐是黄土高原人工造林中应用最为广泛的树种之一。维系当地刺槐人工林的物质循环及其所控制的林分稳定性,使其在未来持续发挥生态、社会和经济效益显得尤为重要。因此,本文以不同恢复年限的刺槐人工林以及油松(Pinustabuliformis,典型的难分解凋落物)和白三叶(Trifoliumrepens,易分解凋落物)为对象,在排除林地凋落物性质变化以及区域尺度气候干扰的条件下,研究凋落物分解特征以及土壤理化性质、土壤微生物群落结构特征和9种凋落物分解酶的活性随林龄的变化规律,并分析土壤微环境变化影响凋落物分解的可能机制。研究结果可为进一步理解人工林植被恢复过程中林地土壤微环境变化对人工林系统物质循环功能的影响趋势,据此为后续营林管理措施的制定和实施提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于陕西省延安市安塞区纸坊沟流域(109°13′—109°16′E, 36°30′—37°19′N),当地地处黄土高原丘陵沟壑区,平均海拔约1200 m。气候分类属于暖温带半湿润半干旱季风气候过渡区,年均降水量549.1 mm,集中于7—9月,蒸发量1463 mm,年平均气温8.8 ℃,年日照总时数2145 h。土壤以黄土母质上发育的黄绵土为主。植被类型属森林草原带,历史上植被严重破坏后自20世纪70年代开始水土保持综合治理并恢复植被,现有乔木林以人为引入的刺槐为主,灌草地主要以柠条Caraganakorshinskii、沙棘Hippophaerhamnoides、白刺花Sophoradavidii以及狗尾草Setariaviridis、达乌里胡枝子Lespedezadaurica、白羊草Bothriochloaischaemum和铁杆蒿Artemisiagmelinii等蒿属植物等为主[7]。
1.2 凋落物样品采集和处理
本项目的前期工作表明,林龄增加过程中,供试林分中刺槐叶片自身的养分特征显著改变,且其林下植物出现明显演替[7]。为避免上述变化以及林地土壤特性和林地原生凋落物间长期互作所产生的“主场效应”影响实验结果[16—17],本研究统一选用在研究区域不存在分布的油松和白三叶的凋落物为对象,以单纯研究林地土壤环境变化对凋落物分解的影响趋势。上述凋落物的养分、难分解物和分解抑制物含量(表1)分别高于和低于供试林分林下植被演替各阶段的优势种及不同树龄刺槐凋落物上述指标对应的极值(见文献[7]),前期研究也已证实两者的可分解性能够完全覆盖供试林地中涉及到的所有凋落物可分解性的最高和最低值(供试林地凋落物的分解速率k为0.296—0.968,相同分解环境下油松凋落物的k低于0.283,而白三叶凋落物的k高于1.716,未发表数据),视为典型的难分解凋落物和易分解凋落物。
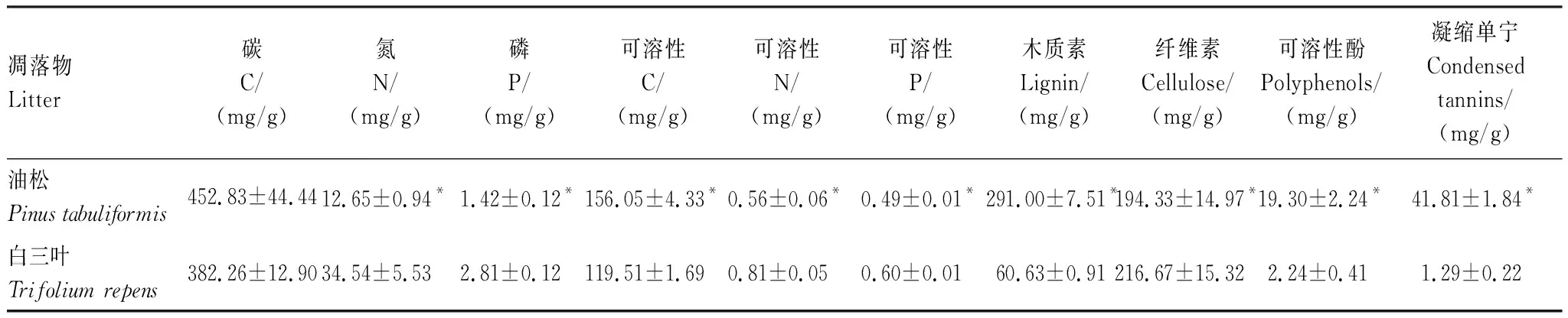
表1 供试凋落物的基质质量Table 1 The substrate quality of the tested litters
于秋末在延安市宝塔区分别选择生长良好的油松群落和白三叶草地,在前者中随机选择20株油松收集其树体上宿存的枯死针叶,随后在实验室避光风干至恒重,并在来源于每株油松的针叶中随机选取等量完整针叶后均匀混合;对于白三叶草地而言,在其内随机建立0.5 m×0.5 m的小样方20个,并在距离地表1 cm处对所有白三叶枯死残体进行刈割、运回实验室风干处理,并在来源于所有样方的白三叶凋落物中抽取等量后均匀混合。另外,对于两种凋落物的风干样品均随机选取3个子样本在65℃下烘干至恒重,据此建立两种凋落物风干质量和烘干质量的比例关系,在后续表述中所涉及的所有“质量”均以此为基础换算为烘干质量。处理完成后,以10.00 g/袋的规格将两种凋落物分解置入大小为10 cm×15 cm,网眼直径为0.5 mm的分解袋中,每种凋落物分别制备120袋。
1.3 样地布设及凋落物分解试验
在研究区选择立地条件和初始造林密度相似、试验布设时林龄分别为10、20、33 a和43 a的刺槐人工林作为研究样地。样地分布海拔介于1117—1245 m,坡度23°—28°,均位于阴坡/半阴坡的中上部,土壤均为黄绵土。试验布设时,林地实际郁闭度分别为0.60、0.64、0.34和0.33。鉴于当地地形相对破碎,未能对每个林龄的林地选择林下环境可重复性足够的多个林分,因此,本研究在前述研究样地内各分别设置3 个面积为20 m×20 m,且尽量间隔10—20 m以上的实验样方,编号后作为重复样方,以在最大程度上保证样方间的本底条件相近且重复间相对独立。
于样品采集翌年3月5日,在每块研究样地的每个实验样方内均随机选择20个位置,并去除其上的植物和凋落物层。随后分别将每份分解袋(10袋油松凋落物和10袋白三叶凋落物)置于各实验样方预定位置的土壤表面。分解袋数量2倍于试验所需,以避免可能存在的动物活动导致的损失。使用聚氯乙烯U形钉将分解袋固定于地表使之与土壤表面紧密接触,以此时间为基准开展凋落物分解试验。
分解过程中,综合考虑季节性天气条件对分解的影响、凋落物的瞬时分解速率和采样便捷程度,分别在分解进行第108、201、389、506天和第592 天分五次回收凋落物。每次回收时,均在各林龄林地的每个实验样方内、同种凋落物的所有分解袋中随机选择1袋,取出凋落物、以毛刷清理干净后在65℃下烘干至恒重保存。待所有样品回收完成后,将每个固定编号实验样方内5次回收的同种凋落物用于拟合1个分解过程,即每个林龄-凋落物组合共拟合3个分解过程(分别取自3个实验样方)作为重复。同时,在分解试验前、中和后期(即分解第108、389天和第592天)采样时,分别在采样前后各3 d和采样当天回收凋落物时(12:00—14:00)使用红外测温仪在各实验样方内随机选择10个位点测定土壤表面温度。在后续分析中,每个样地的表面温度均以上述70个数据的均值作为观测值,以排除不同位点和采样时随机天气过程的影响。同时,在采样当天对每个实验样方内所有测温位点处的表层土壤(0—10 cm)进行等量取样,使用灭菌工具迅速充分混合均匀后以四分法取足量置于冰盒内转移至实验室,并分取3个子样品。前2个子样品在检测前短暂保存在-80℃和4℃中,分别用于土壤微生物群落组成和结构特性分析、凋落物分解相关酶活性以及土壤含水量测定。另1个子样品风干后过1 mm筛,用于土壤速效养分含量和pH分析。在后续分析中,自每个林龄林地的3个实验样方中获取的上述数据视为重复。
1.4 指标测定
采用重铬酸钾外加热法测定凋落物及其分解残留物的有机碳含量,采用靛酚蓝比色法和钒钼黄比色法测定其全氮和全磷含量。采用烘干法测定土壤含水量,采用玻璃电极法测定土壤pH,并分别采用靛酚蓝比色法-紫外分光光度法和磷钼蓝比色法测定土壤的硝铵态氮和有效磷含量[23]。
使用比色法测定土壤中与凋落物分解和养分转化密切相关的漆酶、锰过氧化物酶、木质素过氧化物酶、多酚氧化酶、β-葡糖苷酶、纤维二糖酶、蛋白酶、N-乙酰氨基葡糖苷酶和碱性磷酸酶的活性[7]。另外,参照Zheng等[24]的方法对酶活性数据进行Z-score标准化后求和,分别得到总酶(含所有9种酶)活性和木质纤维素酶(含漆酶、锰过氧化物酶、木质素过氧化物酶、β-葡糖苷酶和纤维二糖酶)活性。
对于回收的新鲜土壤样品,委托陕西轩宸生物科技有限公司完成高通量测序分析。简单而言,采用DNeasy PowerSoil Kit试剂盒根据说明书进行样品总DNA提取,利用琼脂糖凝胶电泳和NanoDrop ND-1000分光光度计检验DNA完整性并进行定量。随后利用条形码特异性引物扩增细菌16S rRNA V4-V5区和真菌ITS1基因。PCR反应条件为在98℃预变性2 min后进行25个循环的98℃变性15 s,55℃退火30 s和72℃延伸30 s,最后在72℃延伸5 min。扩增产物使用Agencourt AMPure磁珠进行纯化并使用PicoGreen dsDNA Assay Kit试剂盒进行定量。随后,采用TruSeq Nano DNA LT Library Prep Kit试剂盒制备测序文库,并使用Agilent High Sensitivity DNA Kit试剂盒进行质检。检测合格的测序文库使用Illlumina Novaseq 6000测序仪进行2×300bp的双端测序。原始下机数据经质量筛查、双端序列配对连接获取样本有效序列,使用QIIME软件去除嵌合体序列。最后,调用UCLUST比对工具按97%的序列相似度对质控后的序列进行归并和OTU划分。获取的代表序列在QIIME软件中与Silva数据库和UNITE数据库的模板序列相比对,获取每个OTU所对应的分类学信息,基于上述数据计算每个样品菌群的Chao1和Shannon指数,并以相对丰度5%已鉴定菌门和属作为优势类群进行分析。
1.5 数据处理和统计分析
凋落物回收后将其残留质量转化为残留率,并使用Olson模型拟合凋落物的分解过程并据此计算凋落物的分解速率k(式1):
k=-ln(mt/m0)/t
(1)
式中,t为分解时间(a),mt和m0分别为t时刻和未分解凋落物的质量(g)。
采用重复测量方差分析检测林龄、时间(采样季节)以及两者间交互效应对土壤化学和生物指标影响的显著性,采用三因素重复测量方差分析检测林龄、凋落物类型、分解时间及三者交互效应对凋落物养分释放影响的显著性。在前述分析中,当Mauchly球形检验未通过时使用Greenhouse-Geisser法对检验结果进行校正。当交互效应显著时,进一步分析林龄因素的简单效应,多重比较校正采用Bonferroni法,显著性检验水平为α=0.05。上述分析使用SPSS 23.0软件完成。
使用非度量多维尺度分析(NMDS)配合置换多元方差分析(PERMANONA)检测不同林龄和时间(采样季节)间林地土壤真菌(或细菌)群落结构差异及其显著性。在后续分析中,使用NMDS排序图中第1轴和2轴的数值反映微生物的群落结构。上述分析使用R软件完成。
最后,使用Pearson相关分析检测凋落物在采样时瞬时分解速率及其与土壤理化性质指标以及土壤微生物群落结构、多样性以及总酶活性之间的相关性,显著性检验水平为α=0.05。使用冗余分析(RDA)检测土壤理化性质指标与土壤优势微生物属的相对丰度以及后者与9种土壤酶活性之间的关系。以上分析分别使用OriginPro 2021b软件和Canoco 5软件完成。使用OriginPro 2021b软件绘图。
2 结果与分析
2.1 凋落物分解特征变化
两种凋落物的质量和碳含量随分解持续降低,而油松凋落物的氮以及两种凋落物的磷则均呈现富集-释放趋势(图1)。凋落物的分解特征受到林龄、分解时间、凋落物种类以及上述因素间的交互作用的显著(P<0.05)或极显著(P<0.01)影响。随林地林龄增加,油松凋落物在刺槐林地中的分解速率呈先降后增趋势(表2,P<0.01),在各分解时间,其碳释放率普遍呈先降后增趋势(图1,P<0.05,仅涉及林龄的表述均为简单效应检测结果,下同),氮、磷释放率不具备明确变化规律,但在分解第502天和第592天时其同样呈现类似趋势(图1,P<0.05和<0.001)。白三叶凋落物在刺槐林地中的分解速率(表2,P<0.001)和碳氮磷释放率则均随林龄增加呈显著提高趋势(图1,P<0.05)。

图1 不同林龄刺槐林地中凋落物的分解及碳氮磷释放Fig.1 Decomposition and C, N and P release of the litters in R. pseudocacia plantations with different stand age数据采用平均值/标准误(误差线)表示,重复测量方差分析结果以各因素的F值及显著性表示,*,P<0.05;**,P<0.01;***,P<0.001;Pt:油松凋落物,Tr:白三叶凋落物

表2 不同林龄刺槐林地凋落物分解特征Table 2 Decomposition characteristics of the litters in R. pseudocacia plantations with different stand age
2.2 林地理化环境特征变化
除硝态氮和有效磷含量以及pH外,林地土壤理化性质普遍受到林龄、分解时间以及两者交互作用的显著影响(表3)。具体而言,林地土壤的表面温度在任一分解时间均随林龄增加呈先降后增趋势(P<0.001),而土壤湿度持续降低(P<0.05)。另外,林地土壤的硝铵态氮含量在分解第108天和第389天随林龄增加显著提高(对应试验布设当年夏季和翌年春末,P<0.001),其pH值在分解第108天随林龄增加显著降低(P<0.05),土壤有效磷含量在分解第592天同样随林龄增加呈显著降低趋势(对应试验布设翌年冬初,P<0.05)。

表3 不同林龄刺槐林地土壤理化性质差异Table 3 Differences in the physical and chemical properties of soil in R. pseudocacia plantations with different stand age
2.3 林地土壤微生物群落特征变化
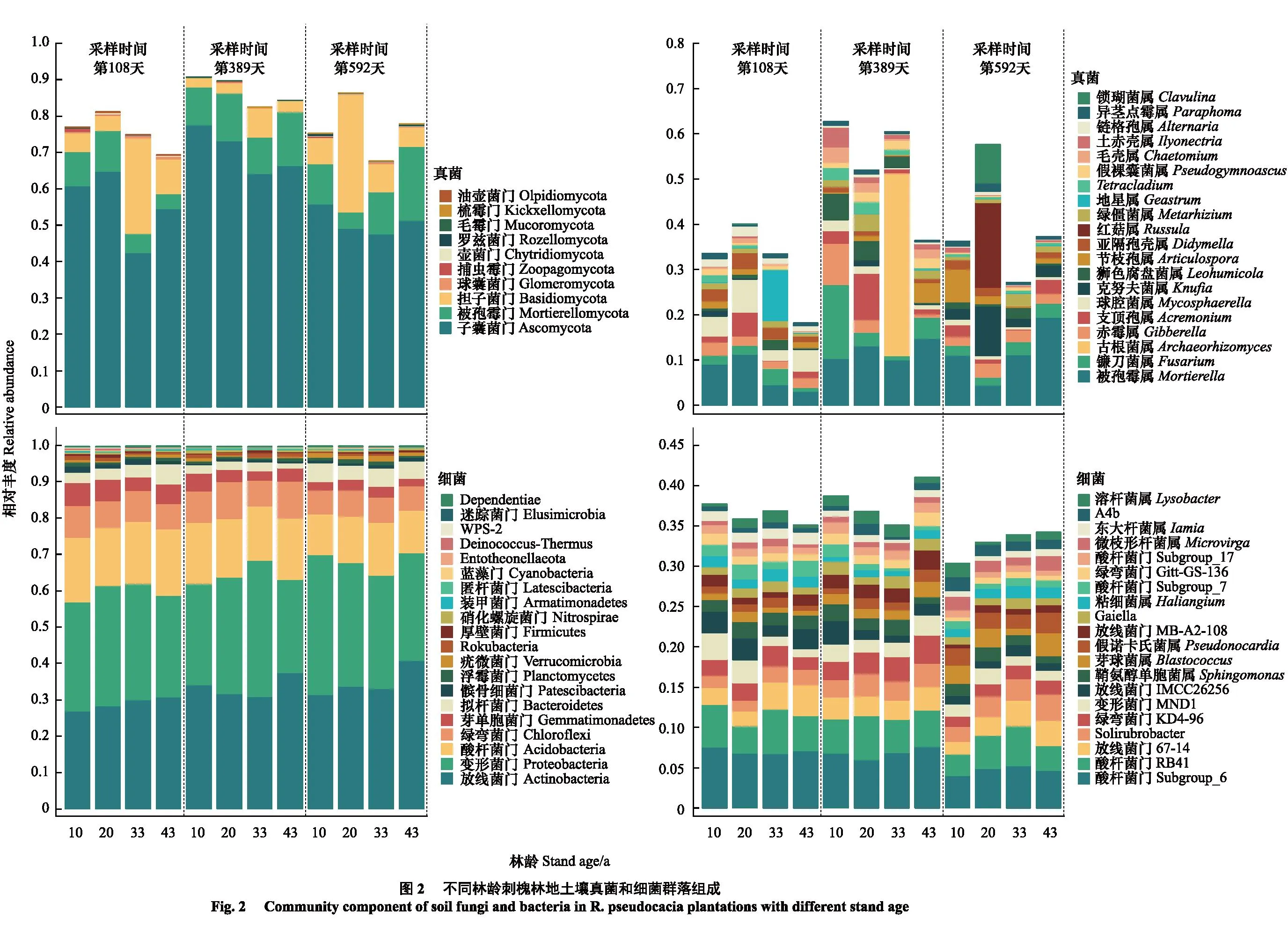
门水平上,土壤真菌中的子囊菌门Ascomycota、担子菌门Basidiomycota和被孢霉门Mortierellomycota以及细菌中的放线菌门Actinobacteria、变形菌门Proteobacteria和酸杆菌门Acidobacteria是10—43 a林地土壤中的共同优势菌门(图2)。其中,担子菌门相对丰度在分解第108天和第389天均随林龄增加呈先增后降趋势(P<0.05和<0.001),被孢霉门丰度在分解第389天随林龄增加显著降低,而在分解第592天则呈先降后增趋势(P<0.05和<0.001)(图2);土壤细菌中(图2)的放线菌门丰度在分解第108天和第389天随林龄增加显著提高(P<0.05),变形菌门相对丰度在分解第389天随林龄增加呈先增后降趋势(P<0.001),在分解第592天则呈降低趋势(P<0.01)。
具体至属水平时,真菌中仅被孢霉属Mortierella是所有林地土壤中共同的优势属,其余属则随林龄增加呈现出明显的演替现象(图2),而细菌中酸杆菌门的Subgroup6和RB41类群是多数林地土壤中的共同优势属(图2)。其中,被孢霉属的相对丰度在分解第108天随林龄增加显著降低(P<0.05),而在分解第592天呈先降后增趋势(P<0.01);镰刀霉属Fusarium和赤霉属Gibberella仅在分解第389天为优势属,且其丰度随林龄增加显著降低(P<0.01),球腔菌属Mycosphaerella和地星属Geastrum,古根菌属Archaeorhizomyces、支顶孢属Acremonium和狮色腐盘菌属Leohumicola以及克努夫菌属Knufia、红菇属Russula、锁瑚菌属Clavulina和节枝孢属Articulospora分别仅在分解第108天、389天和第592天为优势菌属,且均仅在特定林龄林地中具有优势地位。细菌中,Subgroup6类群在分解全程均为优势菌属,但在分解第389天时其相对丰度呈提高趋势(P<0.05),而在分解第592天时仅在33 a林地中具有优势地位;RB41类群仅在分解第389天和第592天时为优势菌属,在分解第389天时其相对丰度随林龄增加呈先降后增趋势(P<0.05),在第592天时则呈相反趋势(P<0.01)。
从α多样性角度分析,林地土壤真菌的丰富度总体仅受到分解时间的显著影响(图3,P<0.001),而其多样性同时受到林龄(图3,P<0.05)、分解时间(P<0.001)以及两者交互作用(P<0.001)的显著影响。其中,其物种丰富度和多样性均在分解第108天时随林龄增加显著降低(P<0.05),其物种多样性在分解第389天呈先增后降趋势(P<0.05)。林地土壤细菌的丰富度和多样性(图3)均受到分解时间以及林龄和分解时间交互作用的显著影响(P<0.001或0.05)。其中,土壤细菌的物种丰富度在分解第108天时随林龄增加呈先降后增趋势(P<0.001),而在分解第389天时显著提高(P<0.05);其物种多样性在分解第389天时随林龄增加呈先降后增趋势(P<0.001)。另外,NMDS分析结果表明(图3),林龄和采样季节对林地土壤真菌和细菌的群落结构均产生显著影响(P<0.001,置换多元方差分析)。
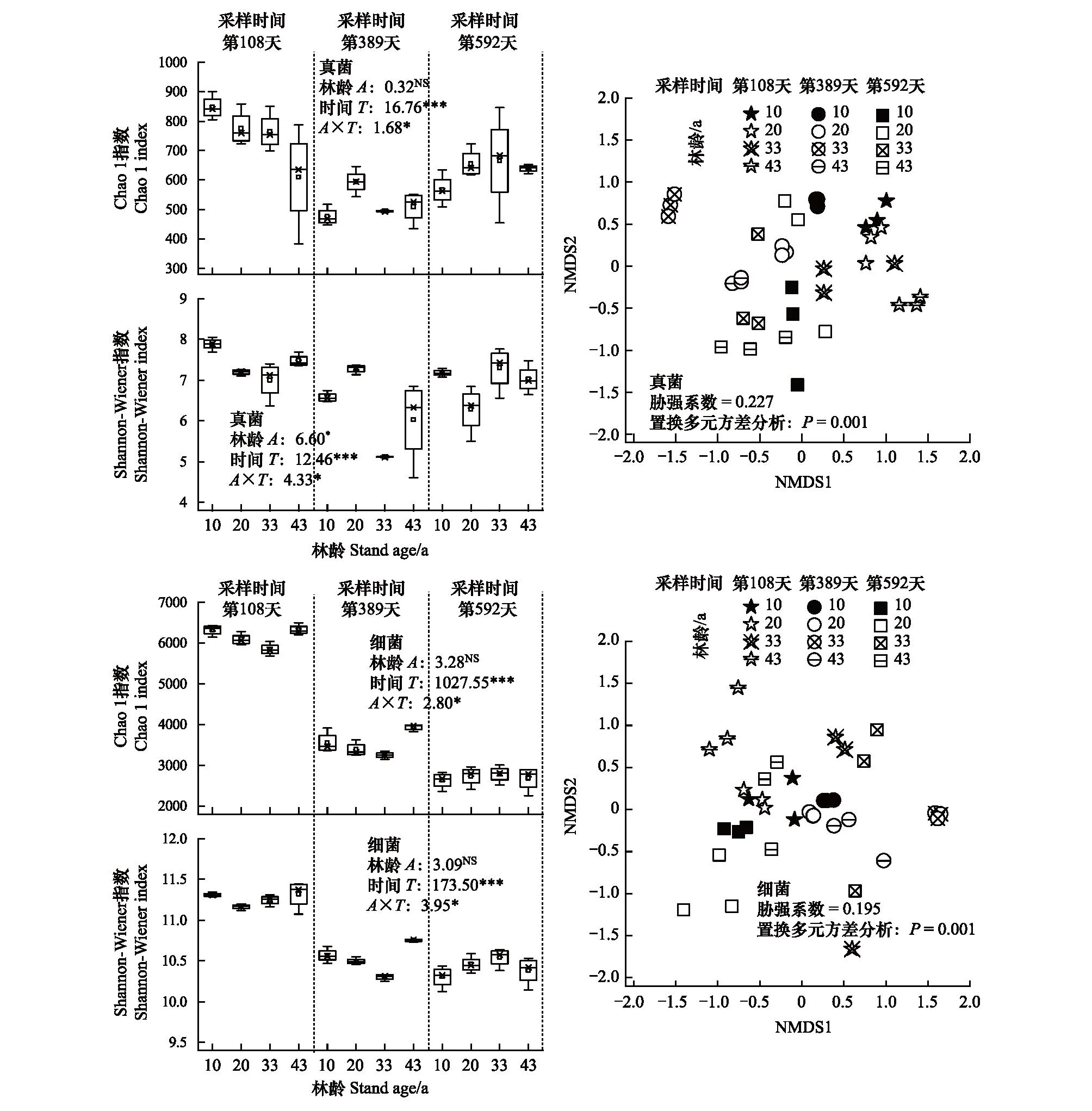
图3 不同林龄刺槐林地土壤微生物的α多样性和菌落结构差异(基于NMDS分析)Fig.3 The α diversity indices and community structure difference (base on NMDS analysis) of soil microbes in R. pseudocacia plantations with different stand ageNMDS:非度量多维尺度分析 non-metric multidimensional scaling;NMDS 1和NMDS 2分别为排序图中两轴的值
2.4 林地土壤酶活性变化
本研究中涉及的土壤酶活性均受到林龄、采样时间及其交互作用的极显著影响(图4,P<0.001)。在上述酶发挥作用的主要阶段(酶活性最高的时期),漆酶和锰过氧化物酶活性随林龄增加显著降低,木质素过氧化物酶、葡糖苷酶和纤维二糖酶活性则呈相反趋势;土壤蛋白酶活性和多酚氧化酶活性随林龄增加呈先降后增趋势,而碱性磷酸酶和乙酰氨基葡糖苷酶活性则呈现先增后降的变化规律。进一步将上述酶综合为总酶活性和木质纤维素酶活性时(图4),两者均受林龄、分解时间及其交互作用的显著影响(P<0.001),两者均在分解第108天时随林龄增加呈先降后增趋势,但在分解第389天和第592天时则呈显著提高趋势(P<0.001)。

图4 不同林龄刺槐林地土壤酶活性Fig.4 The activity of soil enzymes in R. pseudocacia plantations with different stand age数据采用平均值±标准误(误差棒)表示,不同字母表示林龄间差异显著(简单效应),P<0.05;采样时间: 第……天
2.5 凋落物分解与土壤环境因素的关系
以土壤环境指标测定结果以及与其采样测定时间对应的瞬时分解速率进行Pearson相关分析(图5),结果表明油松凋落物分解速率仅与土壤总酶活性、真菌群落结构(NMDS1轴数值,提取自图3)和土壤铵态氮含量呈显著正相关(R=0.34—0.38,P<0.05),而白三叶凋落物分解速率与土壤总酶和木质纤维素酶活性、细菌(NMDS1轴,提取自图3)和真菌(NMDS2轴,提取自图3)群落结构、温度和铵态氮含量呈显著正相关,而与土壤湿度和pH呈显著负相关(R=0.34—0.82,P<0.05)。同时,上述土壤环境因素指标间以及与其余土壤因素间存在复杂的相关关系。

图5 凋落物瞬时分解速率与土壤环境因素间的关系Fig.5 The relationships among the instantaneous decomposition rate of litter and soil environmental factors*:Pearson相关性分析结果达到显著水平(P<0.05);k:凋落物瞬时分解速率;EAt:土壤总酶活性;EAlc:土壤总木质纤维素酶活性;BN1:土壤细菌NMDS分析的轴1值;BN2:土壤细菌NMDS分析的轴2值;FN1:土壤真菌NMDS分析的轴1值;FN2:土壤真菌NMDS分析的轴2值;BCi:土壤细菌Chao1指数;BSi:土壤细菌香农指数;FCi:土壤真菌Chao1指数;FSi:土壤真菌香农指数;Tem:土壤温度;Mois:土壤湿度;pH:土壤pH值;NH4:土壤铵态氮含量;NO3:土壤硝态氮含量;AP:土壤有效磷含量
进一步的RDA分析结果表明,本研究涉及到的6种土壤理化性质仅解释了12个优势土壤菌属相对丰度变化的25.17%,其中土壤温度和铵态氮含量与Subgroup6、RB41、球腔菌属和地星属丰度呈正相关,硝态氮和有效磷含量以及土壤湿度与古根菌属、镰刀菌属、狮色腐盘菌属和被孢霉属含量呈正相关,而pH与克努夫菌属、赤霉属和节枝孢属丰度呈正相关。另外,以上12个优势属的丰度解释了9种主要土壤酶活性变化的63.92%,其中球腔菌属和地星属丰度与漆酶和锰过氧化物酶活性呈正相关,Subgroup6、RB41、赤霉属、支顶孢属和镰刀菌属丰度与多酚氧化酶丰度呈正相关,狮色腐盘菌属、古根菌属和被孢霉属丰度与乙酰氨基葡糖苷酶活性呈正相关,克努夫菌属和节枝孢属丰度则主要与其他5种酶活性呈正相关。
3 讨论
研究结果表明,刺槐林龄的增加显著改变了其林下土壤的温湿度、养分有效性和pH,以及与之关系密切的微生物和酶活性,这与前人研究结果一致[19—20],且本研究结果证实了上述微环境特征的改变的确显著影响了难分解和易分解凋落物在不同林龄林地中的分解特性。
总体而言,油松和白三叶凋落物的分解速率均随林龄增加呈提高趋势,且两者的分解速率的提高均与土壤铵态氮含量、真菌群落结构(以NMDS轴1或轴2数值表示)和土壤总酶活性呈显著正相关。其原因首先可能是微生物的生长及其对凋落物组分的降解需要适宜比例的养分供应。通常在初始状态或在分解过程中,凋落物的碳氮比较之微生物的需求更高,因此在凋落物分解过程中,定殖其上的微生物往往需要从环境中吸收和固定大量氮素[25]。在本研究中,油松和白三叶凋落物在特定分解时间表现出的明显的富集现象也证实了分解初期凋落物处在相对缺氮的状态(图1)。然而随着林龄的增加,刺槐将通过共生固氮微生物和凋落物向林地土壤持续输入氮素[26],从而使土壤的速效态氮素逐渐积累(表2),这与前人研究结果一致[27]。这种环境中氮素可利用性的提高极可能通过在微生物氮素匮乏时快速提供足够的养分、促进其生长和活性,从而加速凋落物的分解。
同时,林地土壤真菌的群落结构随林龄的改变同样显著影响了难分解和易分解凋落物的分解,这与前人关于真菌主导凋落物分解过程的结论一致[8, 28]。其原因在于土壤真菌能够通过菌丝生长侵入凋落物层[29],并连接不同底物进行转移养分[30—31],因而较之土壤细菌具有更强的迁移能力、并能在与叶围土著微生物的竞争中更具优势;另外,真菌能够分泌复杂的胞外酶系,特别是仅有少数真菌类群能够对于凋落物细胞壁中的木质素成分进行彻底破坏[28, 32]。这使得土壤真菌的变化对凋落物的分解的影响更为明显。本研究中,在各分解时间点均发现了土壤优势真菌属丰度随林龄增加发生大幅增减和演替现象。其原因可能是不同真菌类群对理化环境的适应性存在差异[33],林下微环境因素的改变趋势不利于其生长和繁殖,因而导致其丰度下降或优势地位被其他类群取代。例如被孢霉属更适应养分相对贫乏的生境[34],因此在速效氮素随林龄增加最为明显的夏季其丰度在较高林龄林地中显著降低(图2)。此外,本文的前期工作表明,林龄从10 a增加至43 a的过程中,本研究供试林分产生的凋落物总体呈现木质素含量逐渐降低、氮磷含量显著提高的趋势[35]。导致林地土壤中更倾向于利用木质素等复杂组分或贫养底物的镰刀霉属[24]和赤霉属[36]等丰度在特定分解时间随林龄增加趋向降低,而更倾向于利用纤维素和易利用碳源的古根菌属、支顶孢属和被孢霉属(仅分解第592天时)[37—39]丰度呈一定提高趋势。与此同时,随林龄增加,林地土壤真菌的多样性呈现总体降低趋势,且群落结构发生明显改变(图3)。根据前人对于退耕区植被演替过程中土壤微生物网络分析的研究结果,这种改变也可能导致真菌群落随林龄增加演化为一种更为简单、但功能特性更有利于凋落物分解的形式[24]。
当然,较之微生物而言,土壤酶直接催化了凋落物化学组分的降解和转化过程,因而对凋落物分解产生更明显的影响。本研究中,漆酶、木质素过氧化物酶和锰过氧化物酶是催化木质素降解的关键酶,纤维二糖水解酶和β-葡糖苷酶能够逐步将纤维素水解,多酚氧化酶可促进酚类物质及木质素分解中间产物的降解[8,40]。而N-乙酰氨基葡糖苷酶、蛋白酶和磷酸酶活性的提高可为微生物的增殖、菌丝生长以及凋落物分解酶分泌提供更充分的物质和能量来源[40]。因此前述酶、特别是木质纤维素酶活性随林龄增加的总体提高趋势必然显著加速凋落物的分解,这也支持了前文关于林龄增加过程中土壤微生物群落分解凋落物的功能增强的假设。当然,尽管RDA分析的结果支持了土壤酶活性与部分优势土壤真菌和细菌属的相对丰度间存在显著的相关性(图6),但在林龄增加-菌属相对丰度改变以及酶活性-凋落物分解速率改变两个过程之间尚不足以建立明确的联系,其原因可能是尽管大量研究认为土壤真菌在凋落物分解中发挥了关键性作用,但土壤酶活性实际上是由整个微生物群落(包括了真菌和细菌)共同控制的,单纯基于有限的真菌优势属的改变分析其通过酶活性对凋落物的影响尚存在问题,有待进一步的研究加以阐明。

图6 土壤理化性质、优势菌属相对丰度以及酶活性的冗余分析Fig.6 Redundancy analysis of the physical and chemical characteristics of soil, relative abundance of dominant microbial genus and enzymatic activity
尽管两类凋落物的分解速率均呈现总体随林龄增加而增加的趋势,但在此过程中油松凋落物的分解速率在林龄达到20 a时出现了小幅但显著的降低(表2)。其原因可能是仅在温度达到某个阈值时,微生物才能够降解原本不能利用的凋落物组分[41]。栽植20 a后刺槐的快速生长显著增加了林地的郁闭度,使土壤表面获得的太阳辐射大幅减少,随后林木自疏使得郁闭度开始显著降低,地表温度随之回升[42](表3)。因此,20 a林地较低的土壤温度可能是导致微生物分解能力降低、特别是对于木质素和酚类等难分解物含量更高(表1)的油松凋落物分解速率在该时段显著降低的关键因素。20 a林地土壤总酶活性、特别是木质纤维素酶活性在凋落物分解最为旺盛的夏季的显著降低也支持了上述推测。而对于白三叶凋落物而言,其自身较高的基质质量可能更有利于定殖微生物的生长,相对抵消了温度降低引起的分解减缓。需要注意的是,白三叶凋落物的分解速率额外受到了土壤细菌群落结构、温度、湿度和pH的影响。其原因是土壤细菌也可能通过雨滴击溅、风力作用以及真菌菌丝的携带等[31,43]途径进入凋落物层,由于白三叶凋落物的碳氮比、碳磷比和木质素含量等显著较低,更适应此类富养环境的细菌即使仅能少量迁移定殖,也能够快速扩大种群并表现出更大的分解潜力(例如酸杆菌门Subgroup6即具有分解多种碳水化合物的能力[44])。而细菌的群落组成和结构较之真菌更易受到环境因素的影响[22, 45],导致温度、湿度和pH等因素对白三叶凋落物分解产生更为直接和显著的调控作用。另外,尽管两种凋落物的分解速率均受到真菌群落结构的调控,但在白三叶凋落物分解中发挥影响的主要是土壤真菌群落结构的NMDS2轴而非在油松凋落物分解中发挥主要影响的1轴,这表明林龄增加引起的土壤微环境变化的确通过改变真菌群落结构影响2种凋落物的分解过程,但其影响可能是通过不同类群或群落功能的变化驱动的。
4 结论
随林龄增加,刺槐人工林林地表层土壤温度先降后增,土壤湿度、有效磷含量和pH持续降低,而速效氮含量持续提高;土壤真菌和细菌群落结构显著改变,且前者在各分解时间点均随林龄增加出现属水平上的明显演替;土壤总酶活性和木质纤维素酶活性均在分解第108天时呈先降后增趋势,在分解第389 天和第592天时则持续提高。上述现象最终导致在刺槐人工林中,油松(难分解)凋落物的分解速率随林龄增加呈小幅降低后提高的趋势,白三叶(易分解)凋落物的分解速率呈持续提高趋势。综上所述,刺槐林龄增加引起的土壤理、化和生物微环境变化总体倾向于加速凋落物的分解过程。
猜你喜欢
现代园艺(2023年3期)2023-02-01 06:43:32
防护林科技(2020年10期)2020-12-22 11:46:42
现代园艺(2020年2期)2020-12-22 05:57:04
小溪流(故事作文)(2018年10期)2018-12-28 10:45:08
现代园艺(2018年2期)2018-03-15 08:00:12
现代园艺(2017年22期)2018-01-19 05:07:02
防护林科技(2016年9期)2016-09-27 01:34:21
防护林科技(2016年5期)2016-09-05 01:19:43
山东林业科技(2016年5期)2016-07-05 00:43:00
亚热带资源与环境学报(2015年1期)2015-01-22 07:04:58
