
高原鼠兔干扰对高寒草甸植物物种和功能性状beta多样性的影响
2024-05-06 03:20孙文涛庞晓攀徐雪婷郭正刚
生态学报 2024年7期
李 捷, 孙文涛, 庞晓攀, 徐雪婷, 杨 欢, 郭正刚
草种创新与草地农业生态系统全国重点实验室, 兰州大学草地农业科技学院 兰州 730020
物种多样性包括3个不同的水平:局域多样性(alpha多样性)、区域多样性(gamma多样性)以及生境间多样性(beta多样性)[1—2]。alpha多样性和gamma多样性都忽略了物种组成在不同群落间的变化,而beta多样性作为物种多样性的重要测度指标,可以通过反映群落物种组成沿某一环境梯度的变化,体现不同群落多样性的差异程度,从而揭示植物群落演替趋向和沿环境梯度的生态过程[1—3]。目前,大多数草地植物beta多样性研究采用Whittaker指数或是Whittaker指数的单调转换[4—6],仅能体现植物组成在不同群落生境间的相似性,无法全面观测植物beta多样性究竟是通过什么途径变化的。
Baselga提出物种群落的总相似性可以转换为两个部分,即总beta多样性(SOR)可以分解成物种周转引起的beta多样性(SIM)和嵌套导致的beta多样性(NES),为更大空间尺度的草地植物多样性维持提供了科学方法[7]。其中,物种周转暗示同时保护多个位点的多样性维持策略,嵌套则暗示重点保护物种丰富度高的位点[8]。此外,大多数植物beta多样性研究主要基于物种维度,然而物种之间并不相互独立,功能性状在物种间的差异程度是不同的[9—10]。研究发现对于欧洲和美洲的温带植物,在物种beta多样性较高的情况下,植物功能性状beta多样性仍然较低[11]。因此,基于物种维度,并不能很好地揭示驱动beta多样性格局形成的作用力。然而,目前植物功能性状beta多样性的研究主要聚焦于森林生态系统[12—13],而是否能够采用植物功能性状beta多样性的变化途径,分析草地植物群落的构建过程尚不明确。基于功能性状维度的beta多样性研究,为草地群落生态学研究提供了新的视角[14]。
生物干扰是草地植物beta多样性的潜在影响因素。牛莉芹等关于五台山草甸群落的植物beta多样性分析结果表明,旅游中的人为踩踏、建设因素是引起当地草甸植物beta多样性降低的主要原因[6]。同时,草食动物干扰是决定草地植物群落生境异质性的主要小尺度因子,会在草地生境内形成不同的群落生境,改变植物beta多样性。已有研究表明,内蒙古典型草原植被beta多样性随放牧强度的增加逐渐减小[4];羊草杂类草草甸beta多样性随放牧强度增加,呈明显上升趋势[5]。这些研究表明大型草食动物放牧干扰可以改变草地群落植物beta多样性。除大型草食动物之外,小型啮齿类草食动物也是草地生态系统不可缺少的组分[15—19]。研究表明,随高原鼠兔干扰强度的增加,高寒草甸植物物种beta多样性指数表现为先增加后降低的变化趋势[16],随着欧洲地松鼠(Spermophiluscitellus)干扰强度的增加,美国温带草原beta多样性指数表现为逐渐增加的变化趋势[17]。此外,黑尾土拨鼠(Cynomysludovicianus)可以增加不同空间尺度的土壤节肢动物beta多样性[18]。然而这些研究并没有设立未干扰区作为真正的对照区。探讨草地植物物种beta多样性和植物功能性状beta多样性对小型啮齿类草食动物干扰的响应,确定小型啮齿类草食动物如何影响草地植物物种和功能性状beta多样性的变化途径,尚需科学研究提供证据。
高原鼠兔(Ochotonacurzoniae)是青藏高原分布最广的小型啮齿类草食动物,主要栖息于高寒草甸,由于营家族式生活,高原鼠兔野生种群往往扩展迅速,数量庞大[16, 20],是青藏高原高寒草甸主要的生物干扰因子之一[20—22],也被认为是高寒草甸的“生态系统工程师”[22]。高原鼠兔可以通过排泄、采食等多种干扰方式,增加环境异质性[23—25],影响群落物种组成,进而改变高寒草甸植物多样性[16]。然而,目前关于高原鼠兔干扰对高寒草甸植物多样性影响的研究,主要集中于高原鼠兔干扰对局域植物多样性的影响[25—27],关于高原鼠兔干扰下,高寒草甸植物beta多样性变化途径的研究没有提及。本研究采用空间多点同步建立高原鼠兔干扰区和未干扰区,首先确定高原鼠兔干扰对高寒草甸植物物种beta多样性和植物功能性状beta多样性的影响;其次,揭示高原鼠兔干扰下高寒草甸植物beta多样性的变化途径究竟是周转还是嵌套,理论上尝试确证植物功能性状beta多样性能够解析小型草食动物干扰对草地植物群落构建的影响,同时揭示高原鼠兔干扰对高寒草甸植物beta多样性和植物功能性状beta多样性变化途径的影响,为通过高原鼠兔管理而维持青藏高原高寒草甸植物多样性提供科学依据。
1 研究区概况和方法
1.1 研究地区概况
高原鼠兔分布范围广,不同分布区地貌和环境特征差异大,可以生活在多种土壤类型、地形和气候的环境中[19]。为避免土壤、地形和气候条件对研究结果的影响,本研究选择在青藏高原东缘多个地点建立调查样带 (图1)。调查地点的平均海拔高度介于3200—3750 m,年平均降水量为400—653 mm。调查地点为寒冷、潮湿的高原大陆性气候,但研究点基本气候特征有一定差异(表1)。根据中国土壤分类系统和国家土壤调查局资料,调查地点的土壤类型为高寒草甸土。研究地点草地类型均为以莎草科为主高寒草甸。这些高寒草甸已承包给牧户,每家每户将高山草甸分为暖牧区和冷牧区,在过去的几十年里,在4月中旬到10月初期间,冷牧区被围栏围起来,10月中旬到4月初,围栏开放给牦牛和藏羊放牧。调查位点优势种均为莎草科的高山嵩草(Carexparvula)和矮生嵩草(C.alatauensis)。主要伴生种有莎草科植物青藏薹草(Carexmoorcroftii)、豆科植物高山豆(Tibetiahimalaica)、菊科植物长毛风毛菊(Saussureahieracioides)与蒲公英(Taraxacummongolicum)、蔷薇科植物莓叶委陵菜(Potentillafragarioides)等。尽管调查地点境内高寒草甸分布有多种小型啮齿类草食动物,但本研究通过目视法,选择仅有高原鼠兔存在的样地作为高原鼠兔干扰样地(图1)。
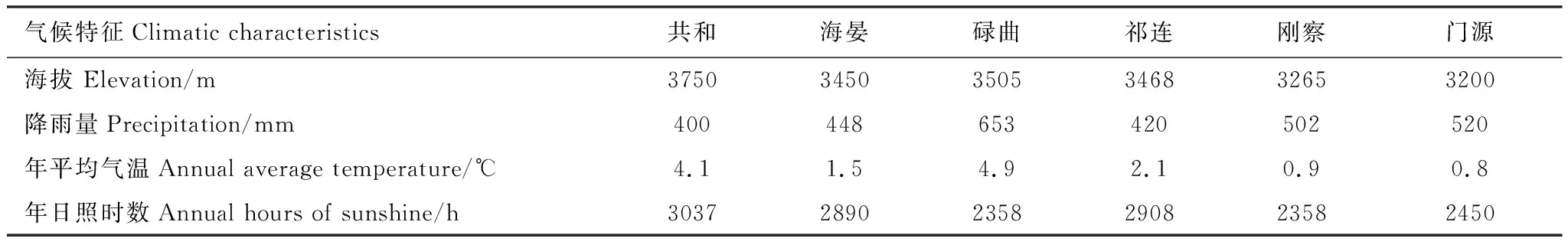
表1 研究区域基本地理气候特征Table 1 The basic climatic characteristics of study sites
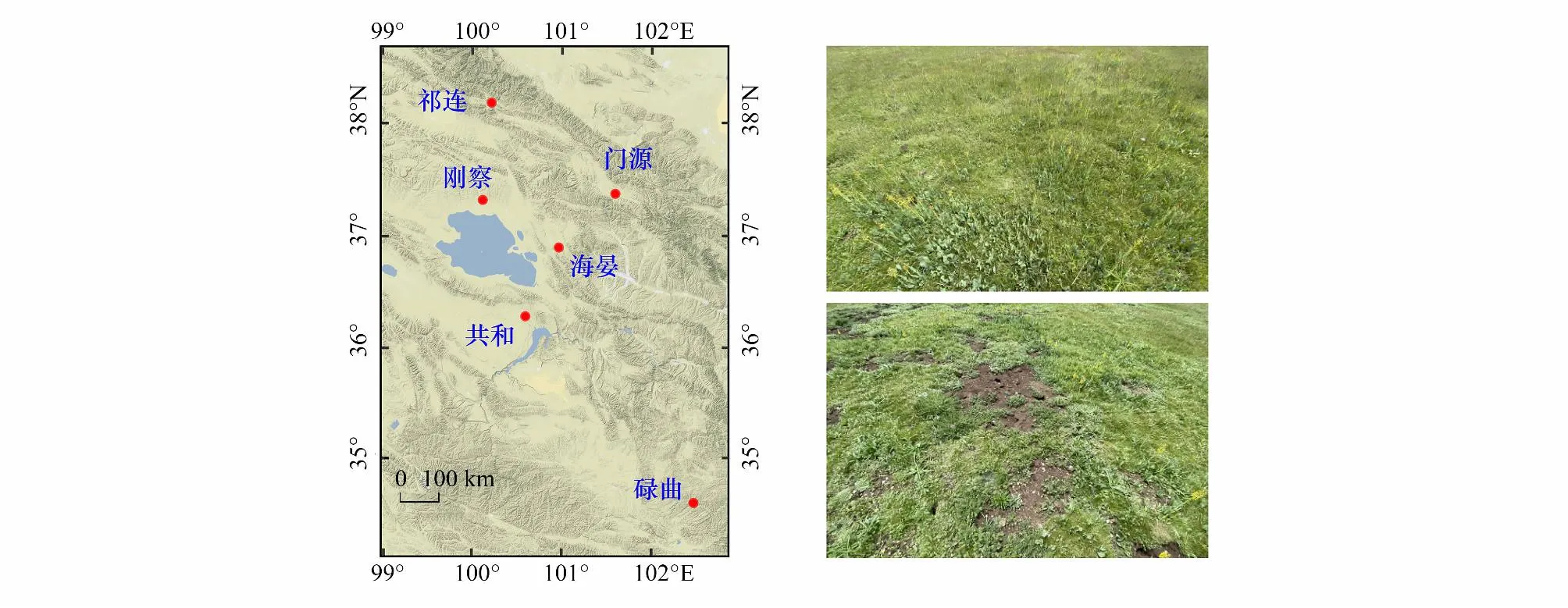
图1 研究地点高寒草甸分布图Fig.1 The sampling sites and the conditions of alpine meadows within the Qinghai-Tibetan Plateau, China, including those both in the absence and presence of plateau pika populations
1.2 试验设计和调查
研究样地设置在全年围封的高寒草甸,属于冷季放牧区(即牧草休眠期牧民打开围栏放牧),暖季(即牧草生长季)不放牧。因此本研究所调查的高寒草甸,从植被返青到植被枯黄,不存在大型放牧动物干扰。高原鼠兔喜栖息于低矮开阔的生境[29],营家族式生活,具有领域性,在高寒草地内呈镶嵌分布[16, 30]。由于高原鼠兔的扩散是一个渐进的过程,有些高寒草甸虽然处于低矮开阔状态,适宜高原鼠兔栖息,但目前没有高原鼠兔生活,因此,目前没有高原鼠兔栖息的低矮开阔草甸,是高原鼠兔的潜在入侵区,可以作为高原鼠兔干扰区的对照区,从而真实测度高原鼠兔干扰对高寒草甸的影响[30]。由于高原鼠兔营家族生活,具有巢域特征,一个家族的高原鼠兔栖息地生境平均面积约为1263 m2,因此,为了最大程度模拟高原鼠兔的巢域特征,本研究设置样地大小为35 m × 35 m[31—32]。
本研究采用随机配对法设置调查样地(调查位点)。首先,选择地势相对平坦、草地类型一致的高寒草甸区,确定是否有高原鼠兔干扰存在[26]。高原鼠兔干扰以高寒草甸内目视有高原鼠兔出没和有效洞口存在为依据[28,26]。首先,通过目视法选定第1个高原鼠兔干扰样地(即为1个调查位点)。选定第1个样地后,继续驱车向任意方向前行,当距离超过5 km时,再次选择第2个干扰样地,以确保两个高原鼠兔家族生活范围不重复,以此类推,所有干扰位点构成一个高原鼠兔干扰样带(图2)[16]。同时,在干扰样地确定后,根据草甸类型和地貌特征,在干扰样地附近选择等同面积的未干扰样地,以没有发现高原鼠兔和高原鼠兔洞口为基准。配对的未干扰样地位于干扰样地的任意方向,与干扰样地的距离大约为500 m到1000 m,距离过大则无法保证该配对干扰和未干扰样地的高寒草甸类型和地形地貌的一致性;距离过小则无法排除高原鼠兔移动对未干扰样地的影响[32]。每个调查地点分别设置10个干扰样地和10个未干扰样地,六个研究地点累计设置120个调查样地(干扰样地和未干扰样地各60个)。
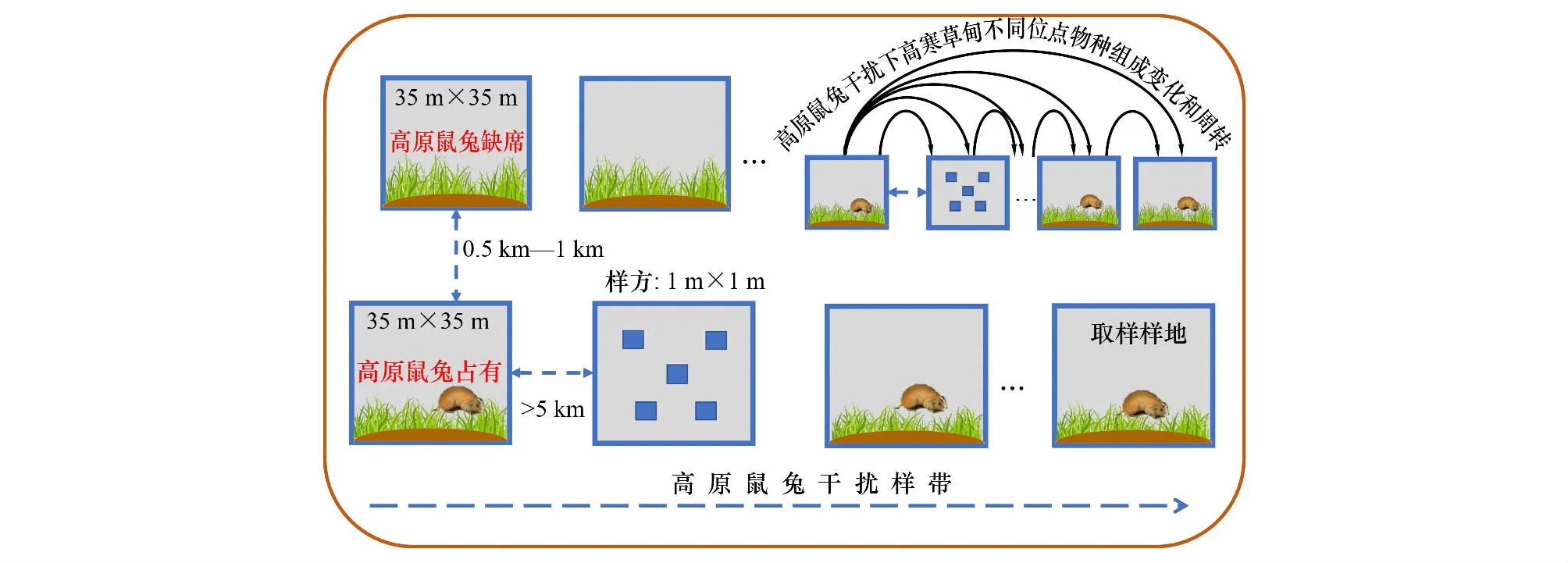
图2 试验样地布设示意图Fig.2 Experimental design schematic diagram
高原鼠兔主要在8月初对高寒草甸的干扰最明显,这是由于高原鼠兔是群居动物,且幼体在出生年份不扩散,因此高原鼠兔种群数量可以在短期内迅速增加,7月底到8月可达到高峰[26]。由于6个调查点年平均气温等气候和地理特征存异,可能导致高原鼠兔的活跃时间在不同调查点分异,因此,本研究确定基本调查时间为7月底到8月,并结合年平均气温和高寒草甸植物生长状况,判断调查点干扰明显期,分别按照碌曲,共和,海晏,刚察,祁连,门源的顺序开展样方调查,以最大程度反映高原鼠兔干扰对植物多样性的影响。在每个样地中以“W”形安排五个调查样方(图2)。样方的大小为1 m×1 m,各个样方之间的距离控制在8 m左右,必要时稍作移动,以避免裸露的土壤斑块,共计600个植被调查样方。本研究首先调查并记录了所有样方中的所有植物种类,并统计每种植物在样方内的丰度。植物高度用卷尺测定物种的高度,当物种株数或丛数高于30,则随机选择30株进行测量,若物种株数或丛数小于30,则全部进行测量。随后采用针刺法测定样方内每个植物的盖度。
测定了12种高寒草甸植物的关键功能性状(或从文献中收集):(1)生长型(半基生、短基生、矮丛、直立多叶和莲座型)、(2)生活型(地上芽、地下芽、地面芽、高位芽)、(3)生活史(一年生和多年生)、(4)叶面类型(光滑、具毛、具刺)、(5)功能类群(禾本科、莎草科、豆科和杂类草)、(6)高度、(7)盖度、(8)丰度、(9)传粉方式(虫媒、风媒和自授粉)、(10)种子扩散(重力传播、风力传播、自体传播,动物传播)、(11)克隆繁殖(地下克隆、地上克隆和无)、(12)固氮类型(固氮和非固氮)。本研究从青藏高原植物区系文献和网站中采集了各植物的生长形态、分布、生活史、传粉、扩散和固氮类型等数据。以上植物物种功能性状同时考虑了生理特征,形态特征,生殖特征,且考虑了选择指标获取的可操作性和易测性。
1.3 数据处理与分析
统计分析与作图均在R语言(版本号:4.2.0)中进行。为验证高原鼠兔干扰对高寒草甸植物群落组成存在影响,本研究对高原鼠兔干扰区与未干扰区调查位点进行聚类分析,确定最佳群落聚类数目,分析方法采用R包“factoextra”,针对调查位点数量较多,优先选择“K-means均值聚类”。高寒草甸植物物种beta多样性的计算采用Baselga提出的方法[33];植物功能性状beta多样性采用Villéger提出并已成功运用的计算方法[9—10]。这两种方法都基于多个调查位点的三个不同的beta多样性系数:(1)Sørensen系数,衡量所有调查位点的总体beta多样性;(2)Simpson系数,衡量由于物种或功能性状在不同位点间的相互替代引起的周转率;(3)嵌套系数,用于衡量由丰富度差异引起的嵌套程度[33—34]。物种和植物功能性状beta多样性的计算,目前已经可以通过R语言实现,计算方式储存在R包“betapart”中[33]。
对于植物物种beta多样性,建立多位点物种组成数据集,采用R语言中的“beta.multi”函数直接实现。对于植物植物功能性状beta多样性,首先整合物种功能性状数据集;其次,基于高尔距离(Gower distance)计算所有草本植物功能性状的距离矩阵,并通过“Lingoes”方法对该功能距离矩阵进行矫正;然后,使用主坐标分析(Principal component analysis, PCoA),选择总解释率85%以上的前几个独立轴,代表所有草本植物的综合功能性状。主坐标分析通过R语言“vegan”包实现。最后,结合多位点物种组成数据集和综合功能性状值,计算基于多维功能特征空间中的凸包体积(即群落中物种所占据的特征空间体积的n维度量),计算过程采用R语言中的“functional.beta.multi”函数实现。
采用线性混合模型(LMM)分析高原鼠兔干扰对高寒草甸物种beta多样性和植物功能性状beta多样性的影响。具体的,将地点设置为随机因子,将配对的高原鼠兔干扰与未干扰样地设置为固定因子,确定高原鼠兔干扰对beta多样性以及周转和嵌套组分的影响是否存在显著差异。线性混合模型分析利用R包“lme4”和“lmeTest”实现。为了进一步明确高原鼠兔干扰下物种beta多样性和植物功能性状beta多样性的主要变化途径,用周转组分与总beta多样性的比值确定周转所占的相对比例,用嵌套组分与总beta多样性的比值,计算嵌套所占的相对比例。
2 结果
2.1 调查位点高寒草甸植物群落组成和功能特征
高原鼠兔干扰区和未干扰区共记录到120个高寒草甸植物群落,累计出现草本植物72种,代表了调查区域的gamma多样性。调查地点物种alpha多样性平均数量为(52±12)种(平均值±标准差)。所有调查位点最丰富的植物物种是矮生嵩草(Carexalatauensis),是调查区高寒草甸的优势种。其他物种均为高寒草甸植物群落常见伴生种,如高山嵩草(Carexparvula)、冷地早熟禾(Poacrymophila)、矮火绒草(Leontopodiumnanum)和美丽风毛菊(Saussureapulchra)等(每个地点的完整植物清单详见补充数据)。
对于高寒草甸植物群落最佳聚类个数,由高原鼠兔未干扰区的17类,转变为高原鼠兔干扰区的9类(图3),表明高原鼠兔干扰显著改变了高寒草甸植物群落组成。对所有72个物种的12个关键性状进行主坐标分析,前三个PCoA轴捕获了89%的草本植物性状变异(测量12种草本植物功能性状的主坐标分析详见补充数据)。
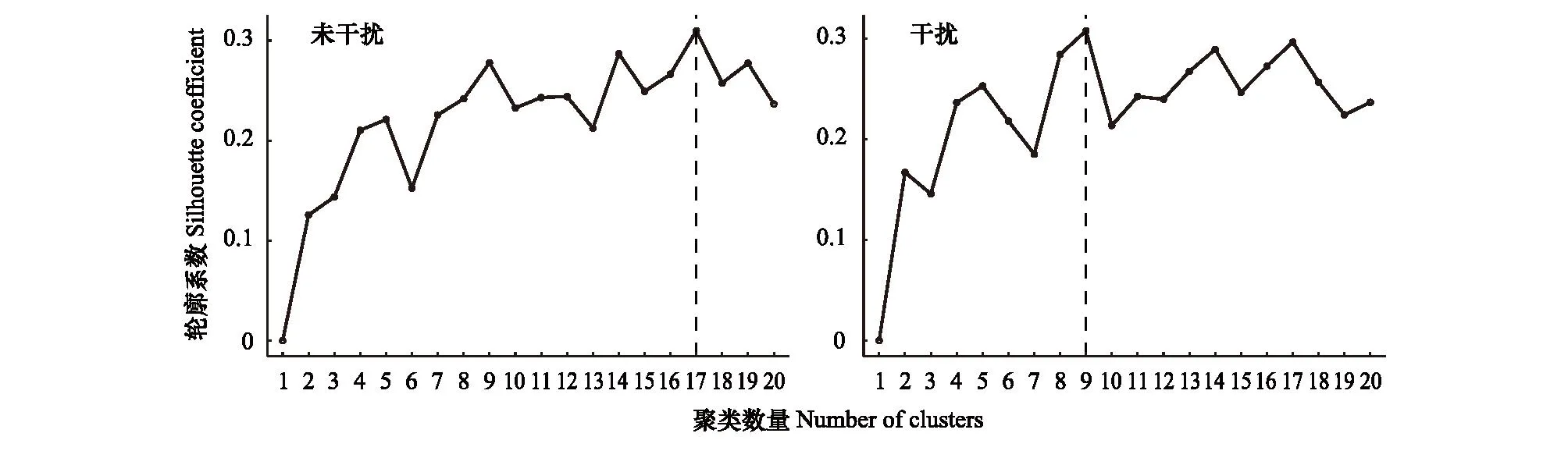
图3 高原鼠兔未干扰区和干扰区高寒草甸植物群落聚类分析Fig.3 The k-means cluster analysis of plant community in alpine meadow高原鼠兔干扰与未干扰区高寒草甸群落组成特征采用均值聚类法分析,虚线对应干扰区或未干扰区60个调查位点的最佳聚类个数
2.2 高原鼠兔干扰对高寒草甸植物beta多样性的影响
高原鼠兔干扰显著降低了高寒草甸植物群落物种的相似性指数(F=15.54;P<0.01),降幅为28.1%。具体表现为高原鼠兔显著降低了物种beta多样性的周转组分值(F=9.86;P<0.01),降幅达36.6%,但对beta多样性的嵌套组分无显著影响(图4)。高原鼠兔干扰显著降低了高寒草甸植物群落功能相似性(F=11.39;P<0.01),降幅为28.7%,同时,显著降低了功能周转组分(F=14.91;P<0.01),降幅为34.3%,但对功能嵌套组分不具有显著影响(图5)。
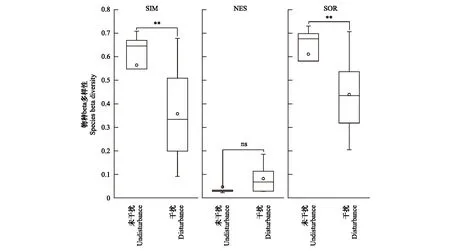
图4 高原鼠兔干扰对高寒草甸植物物种beta多样性的影响Fig.4 Effects of disturbance by plateau pika on plant species beta diversitySIM:植物物种beta多样性的周转组分;NES:植物物种beta多样性的嵌套组分;SOR:植物物种总beta多样性;显著性检验水平为0.05,*表示显著水平< 0.05,**表示显著水平< 0.01,ns表示无显著差异

图5 高原鼠兔干扰对高寒草甸植物功能性状beta多样性的影响Fig.5 Effects of disturbance by plateau pika on plant functional beta diversityF.SIM:植物功能性状beta多样性周转组分;F.NES:植物功能性状beta多样性嵌套组分;F.SOR:植物功能性状总beta多样性
2.3 高原鼠兔干扰下高寒草甸植物beta多样性的变化途径
高原鼠兔干扰下高寒草甸总植物物种beta多样性的周转和嵌套组分表现为SOR、SIM和NES值分别为0.44、0.36和0.08(表2),周转和嵌套组分在总植物物种beta多样性中占比分别是75.4%和24.6%,表明高原鼠兔干扰区高寒草甸植物物种beta多样性变化的主要途径,是物种在空间上的周转。高原鼠兔干扰下高寒草甸总植物功能性状beta多样性的周转和嵌套组分表现为SOR、SIM和NES值分别为0.38、0.13和0.24(表2),周转和嵌套组分在总植物功能性状beta多样性中占比分别是31%和69%,表明高原鼠兔干扰下高寒草甸植物功能性状beta多样性变化的主要途径是物种在空间上的嵌套。
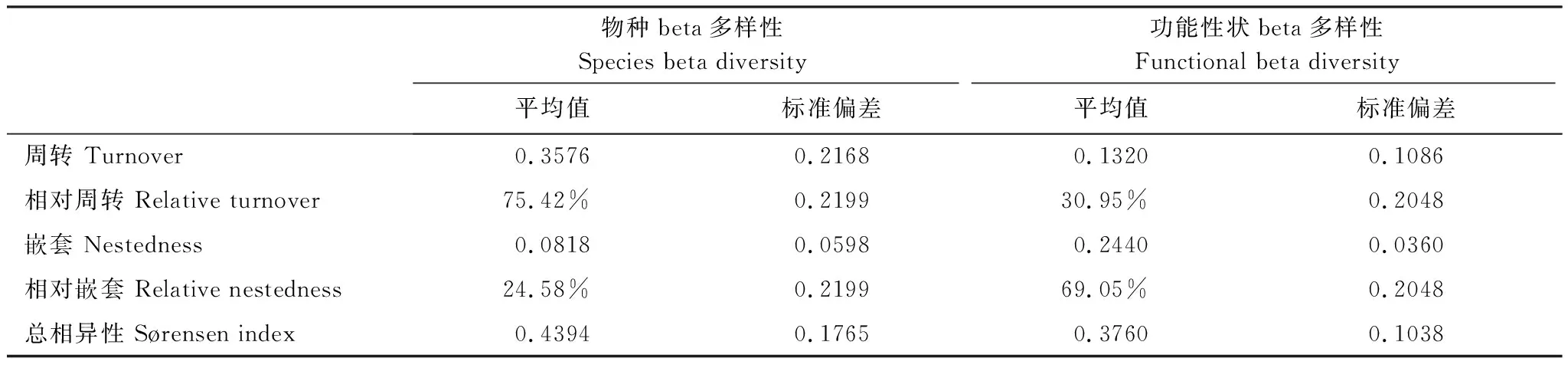
表2 高原鼠兔干扰下高寒草甸植物beta多样性的变化途径Table 2 The pathway of the change of beta diversity induced by plateau pika disturbance in alpine meadows
3 讨论
物种beta多样性体现不同位点之间物种组成的差异程度[1]。本研究中,高寒草甸的植物物种和功能性状beta多样性相对较低(最大值为1),表明调查位点高寒草甸植被组成相似,特有植物种类相对较少,生态独特性较低。该结果是可以预测的,因为本研究所有的野外调查位点,都位于青藏高原东北部的高寒草甸,这些高寒草甸具有相似的栖息地特征、物种相互作用和系统发育历史。例如嵩草属的高山嵩草和矮生嵩草具有广泛的地理分布,而某些植物则在调查区域内被异地(或异地的同源植物)植物所取代。例如,在不同的调查位点中记录了几个密切相关的同源植物:长毛风毛菊(Saussureahieracioides)和美丽风毛菊(Saussureapulchra),刺芒龙胆(Gentianaaristate)和麻花艽(Gentianastraminea)等。
Baselga提出有两种不同的途径会导致物种beta多样性发生变化:一是周转,即一个群落中的物种在另一个群落中被其他物种代替;二是嵌套,即群落中发生物种的增加或丢失,导致物种匮乏的群落是物种较丰富群落的子集[7]。两种变化途径意味着不同的生态过程:地方性的生境异质性、干扰等因素会促进群落产生物种周转,而物种有序灭绝或沿梯度殖民则体现为嵌套格局[34—35]。本研究表明,无论是高原鼠兔栖息地,或者是高原鼠兔未干扰高寒草甸,植物物种beta多样性的大部分差异,均是由物种在空间上的周转引起。如果群落物种组成的差异是随机产生的,并且存在空间自相关等生态过程,则群落组成差异的变化途径表现为物种替换,即可以通过物种周转解释物种beta多样性的变化[35]。研究区域高寒草甸beta多样性主要来源于物种的随机分布和自相关的过程,可能的原因是高寒草甸植物扩散能力相对较弱,因此其分布更容易受植物生长历史以及扩散程度的影响。由于区域内相对隔离的栖息地、气候和物种扩散能力,使一些物种比其他物种受益,造成高寒草甸物种beta多样性变化由周转主导。植物功能多样性主要强调草地植物群落内不同植物物种间功能性状的总体差异,以及植物性状对环境变化的响应[14,31]。尽管功能alpha多样性指数已广泛应用于草地植物群落,并有研究证明高原鼠兔干扰可以改变高寒草甸局域植物群落的功能多样性指数[31],然而植物功能性状beta多样性的研究在草地植物群落中依然匮乏[36—38]。本研究首次将高寒草甸植物功能性状beta多样性的变化途径划分为功能周转和功能嵌套,一定程度上证实了采用植物植物功能性状beta多样性,可以解释高寒草甸植物群落beta多样性变化。高寒草甸植物植物功能性状beta多样性大部分差异,是由功能嵌套引起,一方面表明植物功能性状在物种间的差异程度是不同的[9],另一方面可能是由于高寒草甸植物群落的功能性状是趋同进化而来[31],经生境过滤筛选进入到各个群落的物种功能性状比较相似,造成功能性状在较大空间范围内存在冗余[39]。
高原鼠兔干扰显著影响了高寒草甸植物群落物种组成,具体表现为高原鼠兔干扰下,物种beta多样性减小,群落相似性降低,这是由于高原鼠兔扰动在空间和时间上往往是分散的,可以通过形成洞口、裸斑、重新分配土壤、选择性采食、与土壤动物互作等方式(表3),增加植被空间异质性,改变植物群落间的物种周转。高原鼠兔干扰下,物种beta多样性的周转组分显著降低,表明物种周转对高原鼠兔干扰具有敏感的响应。这是因为高原鼠兔的选择性采食和刈割等行为[40,43],可以引起群落中部分物种的消失[43],同时高原鼠兔粪便以及土壤种子库也会为群落带来新的物种[44],引起群落物种获得率增加,替代率相对减小,最终对群落植物多样性产生影响。本研究发现高原鼠兔干扰对高寒草甸物种嵌套无显著影响,表明物种获得或损失对高原鼠兔响应不敏感。这与荒漠牧区大型草食动物放牧结果一致[45],这可能与高寒草甸自然环境严酷,物种构成稳定、植被均匀等特征有关。已有研究表明在高原鼠兔干扰下,土壤动物丰富性存在明显变化,具体表现为高原鼠兔干扰可以增加鞘翅目成虫、蜘蛛目和鳞翅目幼虫[46],这说明在高原鼠兔下,生态系统食物网的复杂性会有明显增加[16],而土壤食物网的变化亦是小型啮齿草食动物干扰调节下,高寒草甸群落物种组成由随机性过程到确定性过程转变的重要内在原因。高原鼠兔干扰下,高寒草甸植物物种beta多样性以物种周转为主导,说明高原鼠兔干扰区域内的高寒草甸植物多样性维持,要注重保护所有的位点,而非简单的物种保护,因为每个位点由于不同物种的替代,植物物种组合具有较高的独特性。

表3 高原鼠兔干扰草地的方式及影响范围Table 3 The ways and effects disturbance by plateau pika on alpine meadows
研究表明,高原鼠兔干扰下高寒草甸植物植物功能性状beta多样性降低,群落功能相似性降低。这是因为未被高原鼠兔种群占有并扩张的高寒草甸,植物群落通过种内和种间竞争和长时间演替,物种间关联固定,表现为形成以莎草科植物为优势的稳定结构,群落内植物功能特征差异相对稳定[47]。而高原鼠兔干扰下,原来植物群落间物种的关系和种群空间分布特征被打破,给原来竞争弱势的杂类草植物,提供良好的生长环境[48],促进植物各自适应新的土壤生长环境和竞争关系,从而引起植物群落中植物利用和获取资源的途径更加多样化,导致植物个体功能呈现多样化趋势,占据更多生态空间[49],增加了植物群落的功能性状特征,进而减小了群落功能相似性。高原鼠兔干扰会降低功能周转,可能是因为高原鼠兔干扰下,部分植物(例如R策略者),其根系主要生长在土壤表层,导致高原鼠兔栖息区的生境具有旱化特性,这势必诱导一些植物群落组分进一步改变。例如湿生植物湿生扁蕾(Gentianopsispaludosa)、唐松草(Thalictrumaquilegifolium)等具有逐渐减少乃至退出的趋势,而一些中生和旱生植物,例如珠芽蓼(Bistortavivipara)、莓叶委陵菜(Potentillafragarioides)等具有逐渐增多的趋势。植物群落内组分的变化和环境压力增加,导致部分植物强烈的趋同选择[31],会限制植物功能多样性的变化,从而改变植物植物功能性状beta多样性变化成分,导致功能周转所占的比例降低。此外,高原鼠兔干扰有机会在植被上形成肥岛效应[30],增强黄花棘豆(Oxytropisochrocephala)等豆科植物的固氮能力,导致土壤速效氮和全氮含量显著升高(TN >65 mg/kg)[41],冷地早熟禾(Poacrymophila)等禾本科嗜氮植物种间竞争力因此增强,着反而导致矮火绒草(Leontopodiumnanum)等低矮植物生长潜势受到抑制,容易在较高氮浓度下被过滤掉,其余物种通常表现为功能相似,进而降低了物种功能周转的比例。此外,本研究结示了高原鼠兔干扰下,高寒草甸植物功能性状beta多样性以嵌套格局为主导。因此,可以仅通过保护高原鼠兔干扰区域内植物功能性状丰富的位点,维持较高的植物功能性状多样性。
4 结论
植物功能性状beta多样性可以揭示小型啮齿类草食干扰对草地植物多样性的影响从局域多样性向多元群落间的转变过程,有助于从物种和功能保护的视角,分别提出高寒草甸植物多样性的维持策略。高原鼠兔干扰区域内,高寒草甸植物物种beta多样性的变化以周转为主导,植物功能性状beta多样性的变化以嵌套为主导。因此,需要同时保护区域内所有高原鼠兔栖息地,以达到维持植物物种多样性的目的,而可以仅通过保护该区域内植物功能性状丰富的位点,维持较高的植物功能性状多样性。虽然本研究通过嵌套和周转组分量的分析,提出了高原鼠兔干扰下植物多样性的保护策略,但针对面积广袤的青藏高原而言,显然仅有抛砖引玉的作用。分析高原鼠兔干扰引起高寒草甸植物beta多样性的变化途径具有空间依赖性,也暗示青藏高原境内不同地区应该采用不同的生物多样性保护策略。
猜你喜欢
疯狂英语·新读写(2023年4期)2023-05-10
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
系统工程学报(2021年4期)2021-12-21
现代园艺(2021年23期)2021-12-01
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
活力(2019年21期)2019-04-01
计算机工程(2014年6期)2014-02-28
河南科技(2014年23期)2014-02-27
