
晋西黄土区不同密度刺槐人工林叶片-枯落物-土壤化学计量特征
2024-04-14 12:51:42巩大鹏毕华兴王劲峰赵丹阳黄靖涵宋艺琳
林业科学研究 2024年2期
巩大鹏,毕华兴,2,3,4,5,6*,王劲峰,赵丹阳,黄靖涵,宋艺琳
(1. 北京林业大学,北京 100083;2. 林木资源高效生产全国重点实验室,北京 100083;3. 山西吉县森林生态系统国家野外科学观测研究站,北京 100083;4. 水土保持国家林业和草原局重点实验室,北京 100083;5. 北京市水土保持工程技术研究中心,北京 100083;6. 林业生态工程教育部工程研究中心,北京 100083;7. 北京市丰台区水文和水土保持工作站,北京 100165)
林分密度影响了林内光照、水分、养分等重要自然资源,成为制约林木生产力及养分循环的重要因素之一[1-5]。合理的林分密度可以增强林木抵御病虫害的能力,保证林木有一个良好的生长发育环境。
生态化学计量是基于化学计量学,将应用生物学、化学和物理学等多学科相互结合,探索生态系统中物质能量的平衡及其相互关系的方法[6];也是一种研究植物养分,判断限制元素的方法[7]。叶片通过土壤提供的C、N、P 等养分来维持生命活动,最终以枯落物的形式分解将养分返还给土壤,这三者间的相互作用与环境相互影响,使得它们成为了生态系统养分循环的重要过程[8]。目前国内外对植物、枯落物、土壤等的C、N、P 化学计量特征围绕着不同林种、不同海拔、化学计量比与枯落物分解速率相关性等方面展开了大量的研究[9-13]。部分国内学者针对不同树种开展了对密度与植物叶片、枯落物、土壤生态化学计量特征关系的研究调查[14-15]。但由于树种、立地条件、林分结构等因素的不同,密度对叶片、枯落物和土壤间C、N、P 化学计量特征的影响仍有很大的不确定性,尚需进行深入的探讨。
刺槐(Robinia pseudoacaciaL.)具有易繁殖、生长速度快、耐干旱以及适应性强等特点,常用于黄土区的植被恢复,是黄土高原的主要造林树种之一[16]。目前对于刺槐人工林在林分密度方面的研究,主要集中于土壤理化性质[17]、土壤与枯落物的持水能力等方面[18-20]。关于刺槐人工林化学计量特征的研究多集中于林龄方面[21-23],而林分密度对刺槐人工林叶片—枯落物—土壤养分及其化学计量比影响的研究尚未开展。本研究以500、1 000、1 500、2 000、2 500 株·hm-2密度的刺槐人工林为对象,探究叶片-枯落物-土壤养分含量及其化学计量特征随密度的变化趋势,以及三者间的耦合关系,研究结果可为晋西黄土区刺槐人工林的经营提供科学参考。
1 研究地区与研究方法
1.1 研究区概况
研究区位于山西省临汾市吉县蔡家川流域(36°12′29″~36°17′27″ N,110°39′48″~110°47′49″E),该流域属于典型的黄土残塬沟壑区。其总面积约为40.10 km2。海拔920~1 430 m。属于暖温带大陆性季风气候,年平均降水量578.9 mm,降水主要集中于6—9 月,约占全年降水量的70%,年均潜在蒸发量为1 724 mm,远超过年均降水量。褐土和黄土母质为主要土壤类型。主要树种由山杨(Populus davidianaDode.) 、 辽东栎(Quercus liaotungensisMayr.) 等天然次生林,以及人工营造的刺槐(Robinia pseudoacacia)、油松(Pinus tabulaeformisCarr.)、侧柏(Platycladus orientalisLinn.)等水土保持林组成[24]。
1.2 样地布设
试验于2022 年8 月,在研究区选择5 种不同密度(500 株·hm-2、1 000 株·hm-2、1 500株·hm-2、2 000 株·hm-2、2 500 株·hm-2)的刺槐林样地,每种密度设置3 个重复样地(20 m × 20 m),在每个样地内,测定样地海拔、坡度、坡向等立地因子,对样地内的乔木进行每木检尺,测量树高、林分密度、树冠大小等数据,同时记录林龄、林分密度和郁闭度等信息,每个密度梯度下3 块重复样地的海拔、坡度、坡向基本一致,林龄相同,样地基本信息见表1.
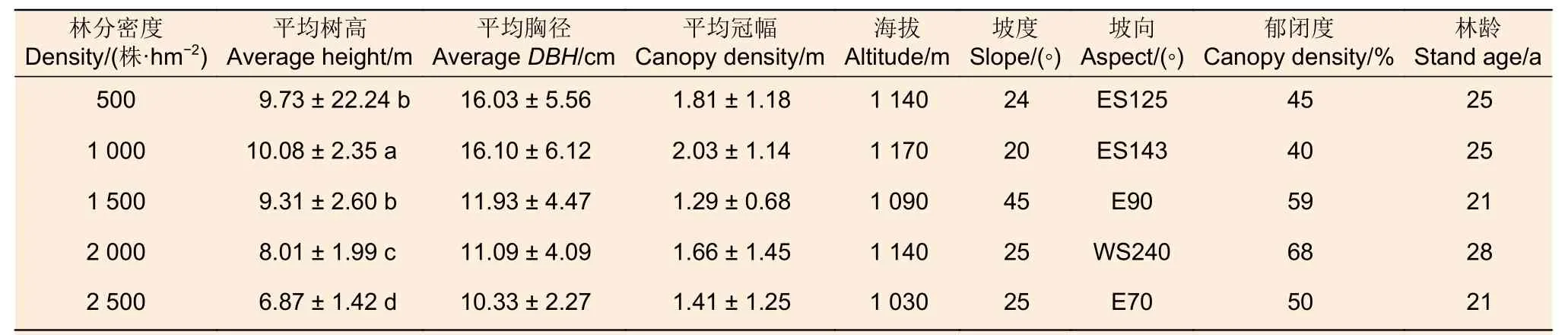
表1 不同密度刺槐人工林样地基本特征Table 1 Basic characteristics of Robinia pseudoacacia plantation plots of different densities
1.3 样品采集与测定
样品采集于8 月12 日开始进行,在每个样地内,选择3 株标准木(具有平均树高和胸径),用高枝剪对树冠东西南北4 个方向,按照上中下3 个部位采集叶片样品,同时收集树干周围的枯落物。将叶片和枯落物分别混匀装入密封袋带回实验室,在80 ℃下烘干至质量恒定,冷却后用粉碎机粉碎,过筛后装袋保存用于C、N、P 含量测定。在清除树冠投影下方的枯落物后,采取五点取样法采集0~30 cm 土层土样,将土样风干、研磨、过0.25 mm 筛后,于实验室测定C、N、P 含量。
植物和枯落物用HCl 溶液去除无机碳后使用德国elementar 生产的有机碳分析仪(vario TOC SELECT)测定有机碳含量,土壤有机碳含量采用采取外加热—重铬酸钾氧化法测定碳含量,土壤、叶片、枯落物样品氮含量用凯氏定氮法测定、磷含量用HClO4—H2SO4消煮—钼锑抗比色分光光度计法测定。
1.4 数据处理
采用EXCEL 2003 进行数据处理,使用origin 作图,运用SPSS 20 软件对数据进行统计分析。用单因素方差分析(one-way ANOVA)和多重比较的方法(LSD,p<0.05)检验不同密度下刺槐人工林叶片、枯落物、土壤C、N、P 及其化学计量比的差异,采用Pearson 相关系数分析它们之间的相关性。
2 结果与分析
2.1 不同密度刺槐人工林叶片的C、N、P 含量及其化学计量比
不同密度间叶片含C、N、P 量随着密度增加表现为,先增加在减小趋势,C∶N 没有显著变化,叶片C∶P、N∶P 随密度增加先降低后增加(图1)。500 株·hm-2和1 000 株·hm-2刺槐林叶片C 含量显著高于其他3 种密度叶片(P<0.05);2 000 株·hm-2和2 500 株·hm-2叶片P 含量显著低于其他密度(p<0.05); 2 000 株·hm-2和2 500 株·hm-2林分叶片C:P 显著高于其他密度(p<0.05)。
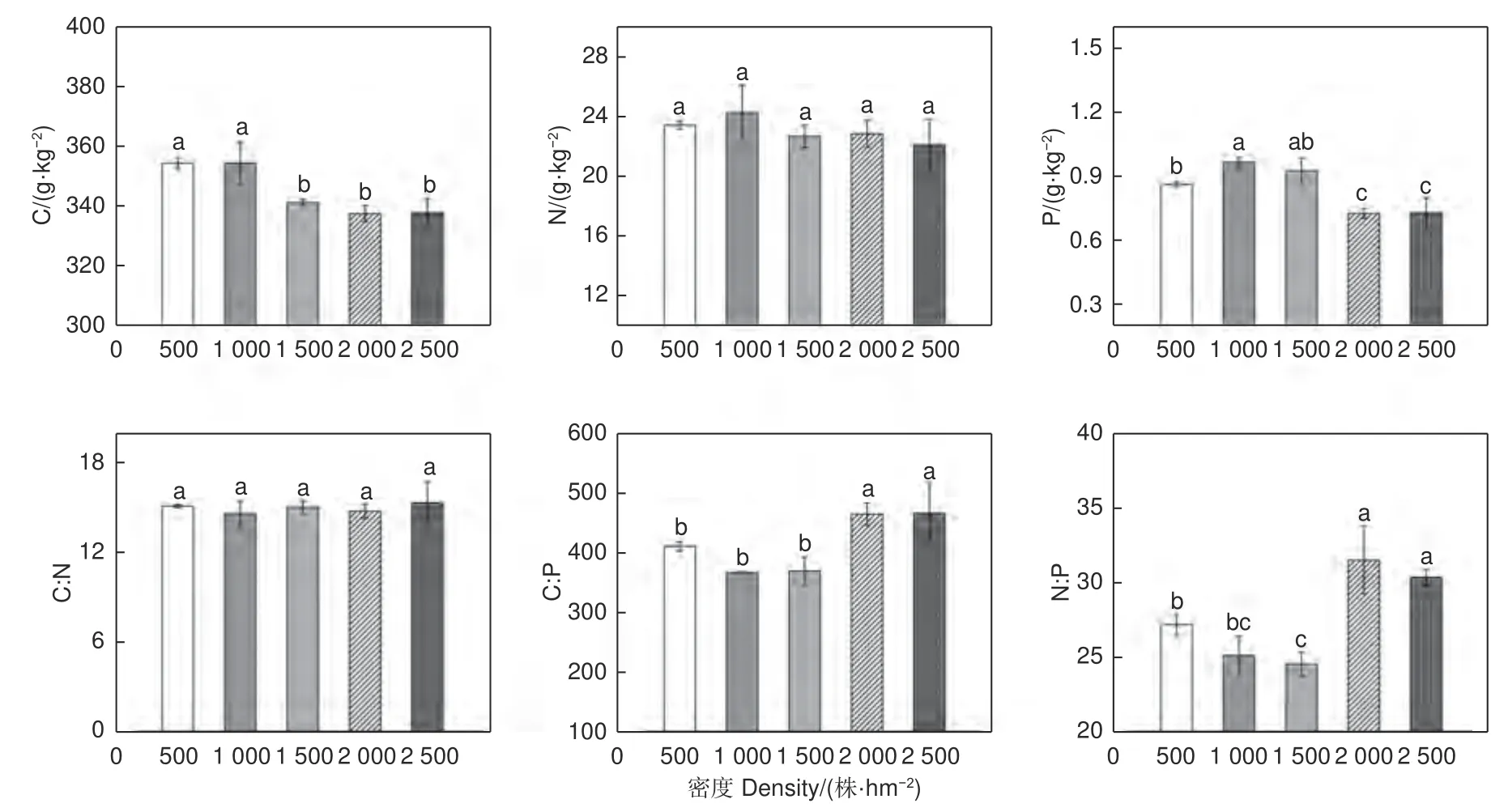
图1 不同密度刺槐人工林叶片C、N、P 含量和C∶N、C∶P、N∶P 值Fig. 1 C, N, P content and C∶N, C∶P and N∶P values of leaves of Robinia plantations of different densities
2.2 不同密度刺槐人工林枯落物的C、N、P 含量及其化学计量比
随着刺槐林密度的增大,枯落物的C∶N、C∶P、N∶P 先减少后增大;枯落物的C、N 变化规律呈“M”字形,枯落物P 呈先增大后减少的规律(图2)。不同密度刺槐林枯落物C 含量存在显著差异(p<0.05),2 000 株·hm-2和2 500株·hm-2的C 含量显著高于其他密度(p<0.05)。1 000 株·hm-2含N 量显著高于其他密度(p<0.05)。1 500 株·hm-2含P 量显著高于其他密度(p<0.05)。1 000 株·hm-2枯落物的C:N 显著低于其他密度(p<0.05)。C∶P 在五个密度间均存在显著差异(p<0.05)。1 500 株·hm-2和2 000 株·hm-2枯落物的N∶P 分别与其他3 种密度枯落物的N∶P 存在显著差异(p<0.05)。
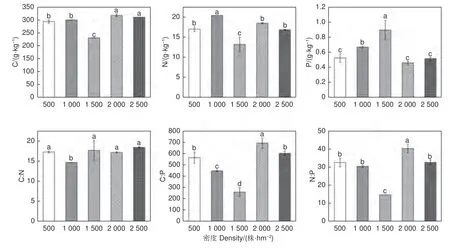
图2 不同密度刺槐人工林枯落物C、N、P 含量和C∶N、C∶P、N∶P 值Fig. 2 C, N, P contents and C∶N, C∶P, N∶P values of litter of Robinia plantations of different densities
2.3 不同密度刺槐人工林土壤的C、N、P 含量及其化学计量比
随着刺槐林密度增大,土壤C 含量及C∶P 降低;P、N 含量及N∶P 先升高后降低,C∶N 先降低后增加(图3)。500 株·hm-2和1 000株·hm-2土壤含C 量分别与其他3 种密度呈显著差异(p<0.05)。1 500 株·hm-2土壤含N 量显著高于其他4 个密度(p<0.05);1 500 株·hm-2土壤含P 与其他4 个密度间存在显著差异(p<0.05),显著高于其他密度土壤含P 量(p<0.05)。C∶N、C∶P、N∶P 区间分别为3.57 ~14.32、6.51~22.95、0.48~2.08,且均在5 个密度间存在显著差异(p<0.05),其中500 株·hm-2土壤C∶P、N∶P 显著高于其他密度C∶P、N∶P(p<0.05);1 500 株·hm-2土壤C∶N 显著低于其他密度的土壤C∶N(p<0.05)。
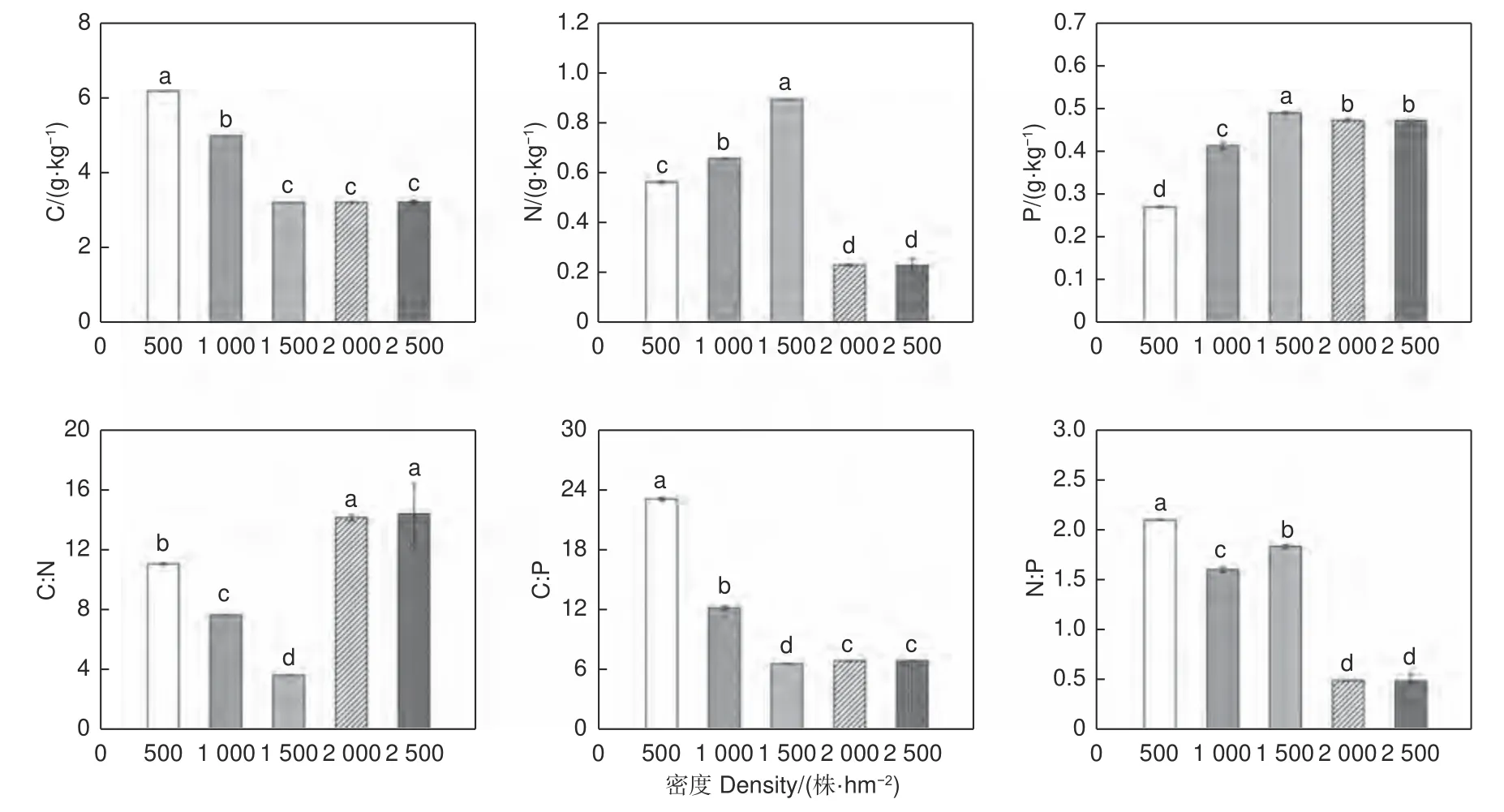
图3 不同密度刺槐人工林土壤C、N、P 含量和C∶N、C∶P、N∶P 值Fig. 3 C, N, P content and C∶N, C∶P and N∶P values of soil of Robinia plantations of different densities
2.4 刺槐人工林叶片-枯落物-土壤C、N、P 含量及其化学计量比之间的相关性
由表2 可以看出,叶片N 与叶片C 呈正相关关系;叶片P 与叶片C、N 呈显著正相关关系(p<0.05)。枯落物C 与叶片P 呈显著负相关关系(p<0.05);枯落物N 与枯落物C 呈极显著正相关关系(p<0.01);枯落物P 与叶片P 呈极显著正相关关系(p<0.01),与枯落物C 呈极显著负相关关系(p<0.01)。土壤C 与叶片C 呈极显著正相关关系(p<0.01);土壤N 与叶片P、枯落物P 呈极显著正相关关系,与枯落物C 呈极显著负相关关系,土壤P 与叶片C、土壤C 呈极显著负相关关系(p<0.01)。
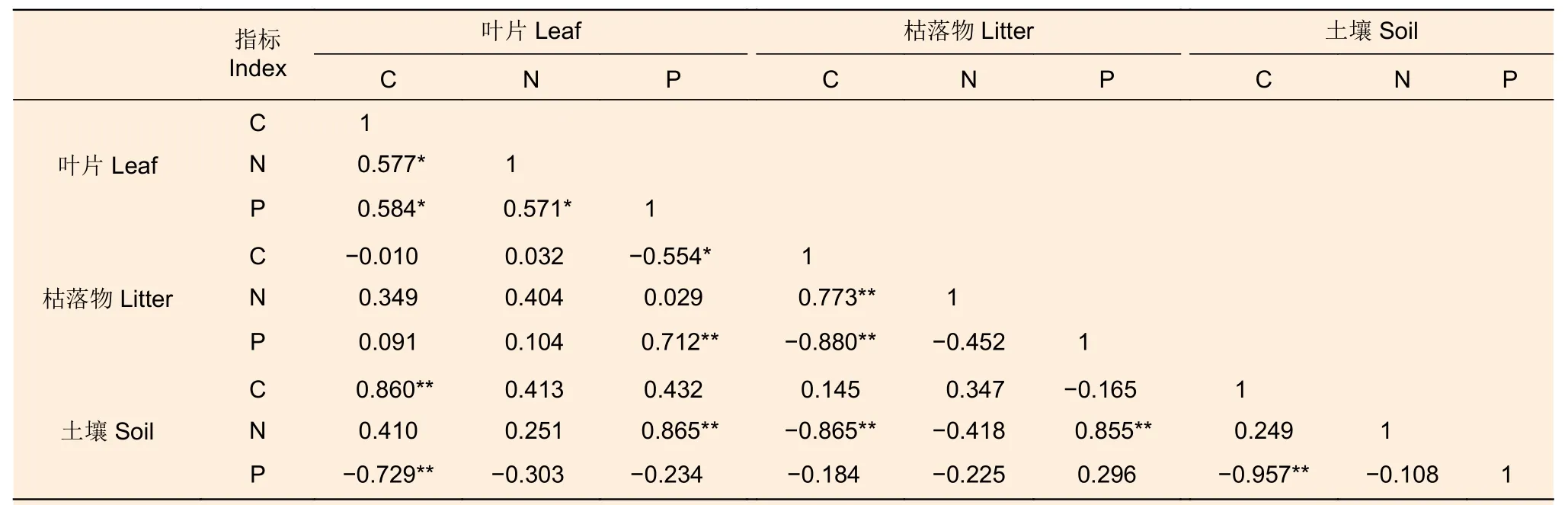
表2 刺槐人工林叶片-枯落物-土壤C、N、P 含量的相关性 Table 2 Correlation of leaves-litter-soil C, N and P contents in Robinia plantations
由表3 可以看出,叶片N∶P 与C∶P 呈极显著正相关关系(p<0.01)。枯落物C∶N 与叶片C∶P 呈显著正相关关系(p<0.05);枯落物C∶P 与叶片C:P、N:P 呈极显著正相关关系(p<0.01);枯落物N∶P 与叶片N∶P、枯落物C∶P 呈极显著正相关关系(p<0.01),与叶片C∶P 呈显著正相关关系(p<0.05)。土壤C∶N 与叶片C∶P 、N∶P 和枯落物C∶P、N∶P 呈极显著正相关关系(p<0.01);土壤N∶P 与叶片C∶P、N∶P 和土壤C∶N 呈极显著负相关关系(p<0.01),与枯落物C∶P、N:P 呈显著负相关关系(p<0.05),与土壤C:P 呈极显著正相关关系(p<0.01)。
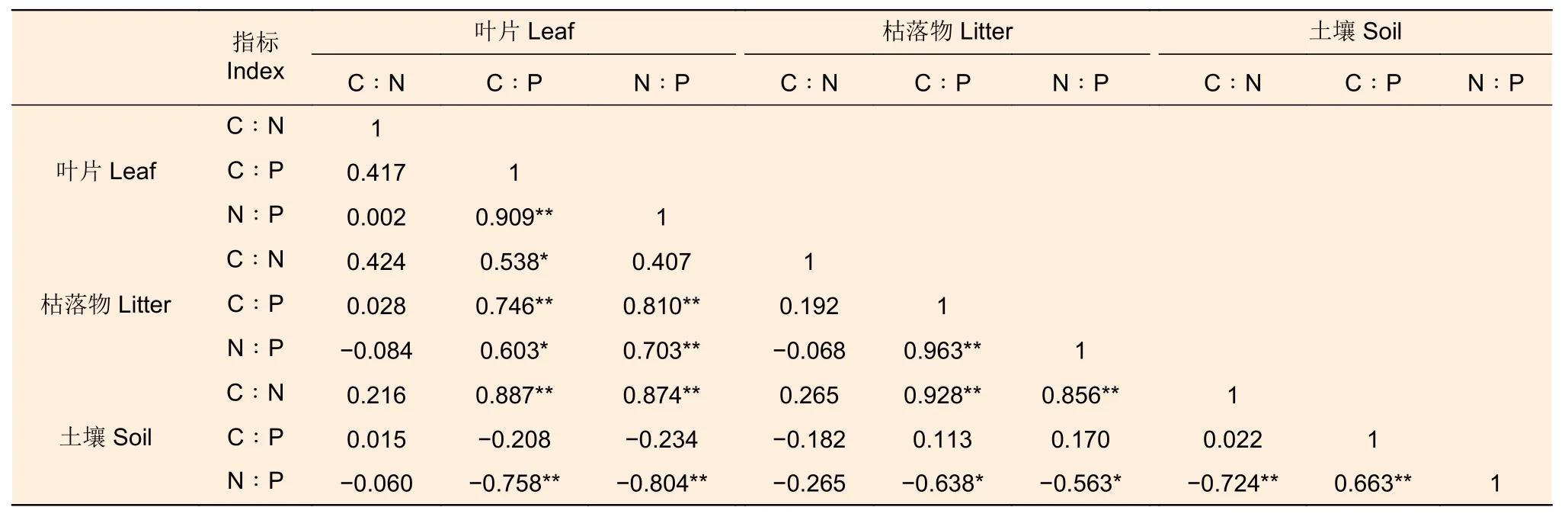
表3 刺槐人工林叶片-枯落物-土壤C、N、P 化学计量比的相关性Table 3 Correlation of leaf-litter-soil C, N and P stoichiometry ratios of Robinia acacia plantations
3 讨论
3.1 不同密度刺槐人工林叶片的C、N、P 含量及其化学计量比
研究区5 种密度的刺槐人工林叶片C、N、P 含量的均值分别为345.11 g·kg-1、23.08 g·kg-1、0.84 g·kg-1,其中叶片C、P 含量明显低于全球492 种陆地植物叶片C、P 平均含量464 g·kg-1、1.99 g·kg-1[25];N 含量高于全球植物叶片N 含量20.09 g·kg-1,也高于我国753 种陆地植物叶片N 平均含量18.6 g·kg-1[26]。说明晋西黄土区的刺槐叶片N 含量较为丰富,这主要是由于刺槐是豆科树种,其根的具有根瘤菌,拥有固氮的能力,能吸附空气中的氮氧化物[21]。刺槐林叶片C 含量随林分密度的增大逐渐降低,这可能是由于林分密度较低时,种间竞争不激烈,叶片可以维持较好的光合同化产物量,但随着林分密度增大,种间竞争加剧,林内透光率下降,导致植物叶片光合作用能力减弱[3]。
本研究发现密度对叶片的C、N、P 含量有一定影响,C∶N 保持相对稳定,C∶P、N∶P 会随密度变化而发生改变,说明密度对叶片生长和N、P 养分限制有一定影响,这与王凯等[27]研究结果相同。C、N、P 对植物的生长发育都起到了相当重要的作用,其生态化学计量比体现了植物对养分的吸收效率、固C 效率以及N、P 养分的限制格局[28]。快速生长的植物具有相对较低的C∶N 和C∶P[29],且生长速率与N∶P 呈负相关关系[30],说明适宜密度更有利于叶片生长。在1 000 株·hm-2和1 500 株·hm-2的刺槐林中,叶片C、N、P 含量较高,C∶P、N∶P 较低,说明养分更多分配到叶片上,促进叶片寿命增长,以便获得更多光合产物,提高固C 效果,为适宜经营密度。叶片N、P 含量呈先增加在减小趋势,这可能是因为林分密度过小,林分郁闭度低,过强的光照使得土壤含水量下降,这导致土壤养分的有效性下降,不利于植物吸收,所以低密度植物叶片养分含量较低;当林分密度增大,种间竞争加剧,植物会通过提高N、P 利用效率以此缓解养分胁迫的影响,从而使叶片养分升高,但随着密度增大,植物对养分的需求量增大,导致土壤养分下降,使得单株植物养分的积累量降低,从而使叶片N、P 含量随着密度增大开始下降[4,27]。植物体中N 和P 作为主要的限制性营养元素,对植物结构、功能以及生长有着一定的影响[31]。因此,可以通过N∶P 大小判断土壤养分的限制情况,当叶片N:P<14 时,植物生长主要受N 限制;当N:P>16 时,植物生长主要受P 限制;当14<N:P<16 时,植物生长受N 和P 共同限制[32]。以此作为判断依据,晋西黄土区不同密度的刺槐人工林N∶P(24.52~31.50)均大于14,说明该地刺槐林生长主要受到P 限制。
3.2 不同密度刺槐人工林枯落物的C、N、P 含量及其化学计量比
枯落物是植物与土壤间的纽带,在生态系统中的养分循环发挥着重要作用。其C、N、P 含量及其化学计量比可以反映土壤养分的供给与植物对养分的利用状况[33]。研究区5 种密度的刺槐人工林枯落物的C、N、P 含量均值分别为290.31 g·kg-1、17.16 g·kg-1、0.61 g·kg-1,其中N 含量高于全球401 种木本植物N 含量10.90 g·kg-1[34],充分说明刺槐对N 的利用效率较高。5 种不同密度的枯落物的C:N、C:P、N:P 大致呈先降低后升高的趋势,密度对它们有着明显扰动作用,这与王凯等研究结果相同[27],这可能是因为密度影响植物对衰老叶片C、N、P 含量的利用效率,从而改变枯落物的C、N、P 含量,使得其化学计量比随密度变化而变化[27]。枯落物的C:N、N:P 可以反应枯落物的分解速率,其分解速率与C:N、N:P 成负相关关系[11]。有研究表明,当C:N<40 时,会出现枯落物净N 矿化现象,且矿化作用会随着枯落物C∶N 减小而变得越发明显[35];C:P 低于600 时,枯落物P 开始释放[36];当枯落物N∶P>25 时,枯落物的分解速率受到P 的限制,因为P 含量较低时,枯落物具有较高的N 和木质素,由此造成分解速率降低,N:P 越大受到P 限制越强烈[37]这反映出刺槐林的枯落物分解速率以及养分返还受密度影响。1 500 株·hm-2枯落物的N 含量最低,可能是由于该密度下枯落物的分解速率较快,部分N 已经返还给土壤,而1 500 株·hm-2刺槐人工林土壤N 含量显著高于其他密度,可以说明这点[4]。1 500 株·hm-2枯落物的P 含量较高,可能是因为该密度下土壤P 含量丰富,使叶片对衰老、凋落叶片P 的重吸收率降低,让其保有了更多P 用于返还给土壤中[27]。在研究区中,除1 500 株·hm-2刺槐人工林枯落物N∶P 低于25,其分解速度受N 限制,其余密度均高于25,分解受P 限制。
3.3 不同密度刺槐林土壤的C、N、P 含量及其化学计量比
土壤中C、N、P 含量在一定程度限制林木的生长发育,其化学计量比反映土壤对林木养分的供应情况[33]。本研究区中,刺槐人工林土壤C 随着密度的增大而减少,N、P 含量随密度的变化呈先增大后减少的趋势,C∶N、C∶P 随密度增加呈先减少后增大的趋势,这与王岩松等[17]研究结果相同。这可能是因为枯落物量随着密度的增大而增加,从而增大了对土壤养分的补充,但林分密度的持续增长,加剧了种间竞争,林木为了生长发育消耗了大量的N 和P,并且过密的林分结构降低了林内透光度,减少了土壤水分的含量,影响了水热气肥条件,从而对土壤中酶的活性产生影响,使得枯落物分解速率降低,影响养分回归土壤以及土壤固化C、N、P 的能力[38]。有研究表明在P 缺乏地区,土壤C∶P 越低,植物生长越快[39]。研究区中,1 500 株·hm-2刺槐林的C∶P 最低,证实1 500 株·hm-2为适宜生长的密度。而随着林分密度继续增大,林木间的生存空间变小,对养分竞争变得越来越激烈,从而消耗了大量的P,使得P 对人工刺槐林的限制更强[40]。
3.4 刺槐林叶片-枯落物-土壤C、N、P 含量及其化学计量比的相关性分析
枯落物C 与叶片P 呈显著负相关关系(p<0.05),枯落物P 与叶片P 呈极显著正相关关系(p<0.01),枯落物C∶N 与叶片C∶P 呈显著正相关关系(p<0.05);枯落物C∶P 与叶片C:P、N:P 呈极显著正相关关系(p<0.01);枯落物N∶P 与叶片N∶P,与叶片C∶P 呈显著正相关关系(p<0.05),这表明枯落物的养分部分承载自叶片,二者关系紧密。土壤C 与叶片C 呈极显著正相关关系(p<0.01),土壤N 与叶片P、枯落物P 呈极显著正相关关系,土壤P 与叶片C 呈极显著负相关关系(p<0.01)。土壤C∶N 与叶片C∶P、N∶P 和枯落物C∶P、N∶P 呈极显著正相关关系(p<0.01);土壤N∶P与叶片C∶P、N∶P 和土壤C∶N 呈极显著负相关关系(p<0.01),与枯落物C∶P 呈显著负相关关系(p<0.05)。这说明土壤是叶片养分的主要来源,枯落物分解是土壤养分重要的补充途径。这与任悦等[41]对樟子松人工林N、P 的化学计量特征以及赵成娇[42]对樟子松人工林的化学计量特征研究结果相似;与王亚娟等[43]对油松人工林的生态化学计量特征研究结果不一致,说明植物与土壤间的化学计量关系存在的差异,受不同林分结构,气候环境差异等因素影响[44]。不同密度的刺槐人工林C、N、P 含量均表现为叶片>枯落物>土壤,这与白雪娟等[29]研究结果一致。植物光合作用主要是在叶片中进行,新陈代谢旺盛,其C、N、P 含量相对较高[33]。枯落物养分低于叶片,是因为N、P 在叶片衰老时,逐步转移到新鲜年轻的叶片,这种行为表现出植物对养分利用效率,降低对土壤养分的依赖[45]。
4 结论
(1)不同林分密度下,刺槐林叶片N∶P 均大于14,表明晋西黄土区刺槐林生长主要受到P 限制,2 000 株·hm-2和2 500 株·hm-2的林分受P 限制最严重。
(2)不同林分密度之间,叶片-枯落物-土壤C、N、P 及其化学计量比中除了叶片N、C∶N 以外,均存在不同的显著差异性,表明密度在一定程度上影响着刺槐林的养分分配和化学计量特征。
(3)1 000~1 500 株·hm-2的刺槐林叶片C、N、P 含量最高;枯落物和土壤N、P 含量最高;受养分限制影响最小,枯落物养分分解释放速率较快,为适宜经营密度范围。
猜你喜欢
落叶果树(2021年6期)2021-02-12 01:28:54
现代园艺(2020年2期)2020-12-22 05:57:04
文苑(2020年12期)2020-04-13 00:54:10
海峡姐妹(2019年1期)2019-03-23 02:42:46
小溪流(故事作文)(2018年10期)2018-12-28 10:45:08
现代园艺(2018年2期)2018-03-15 08:00:12
现代园艺(2017年19期)2018-01-19 02:50:08
现代园艺(2017年21期)2018-01-03 06:42:24
山东林业科技(2016年5期)2016-07-05 00:43:04
山东林业科技(2016年5期)2016-07-05 00:43:00
