
间作对作物、土壤及微生物影响的研究进展
2024-03-25 17:58:12欧阳子龙贾湘璐石景忠韦妙琴滕维超
江苏农业科学 2024年2期
欧阳子龙 贾湘璐 石景忠 韦妙琴 滕维超
摘要: 间作是一种生态农业,通过植物间优势互补来提高植物的养分积累以促进生长。筛选出优良的间作模式可实现增产增收。首先,间作通过地上部生态位提高植物对光资源的利用能力,并协同地下部生态位提高植物抗逆性和抗病性,直接促进产量增加。其次,间作能改良土壤结构,改变土壤理化性质,提高养分含量和酶活性。较好的土壤条件又能正向促进植物生长、生理和养分吸收,体现出多样性净效益,使得间作系统整体产量提高。最后,间作亦会丰富土壤中微生物群落多样性,改良土壤,平衡有益和有害微生物,对植物生长发育有利。本文从植物生长生产、土壤理化结构和土壤微生物等方面综述了近10年来国内外间作研究概况,并对间作的生态和经济效益、研究的重点方向、深层次机理的探究、产业面临的瓶颈和高通量测序技术的应用这5个方面进行了展望,以期揭示间作对植物、土壤和微生物的影响,为未来间作产业发展提供经验和参考。
关键词: 间作;植物;土壤微生物;土壤酶
中图分类号:S344.2 文献标志码:A
文章编号:1002-1302(2024)02-0018-13
进入到21世纪,随着人口数量不断增多、城乡土地面积比例失调,耕地面积逐渐减小,紧张的耕地使用率让人们更多地思考如何在有限的土地面积上收获更高的产量。基于此,间作被有效运用于国内外农作耕种中,在提升自然资源利用率[1]、增强植物抗病抗逆性[2]、提高单位土地面积产量[3]等方面具有极其重要的作用。
间作是一个非常古老的概念,2种或以上植物于同一生长期内间隔种植的模式为间作,主要运用于农业生产。公元前100年人们进行瓜豆间作并将其方法记载到《泛胜之书》中,这是可考的最早间作记录[4]。明朝时民间已经开始间作棉花‖红薯、小麦‖大豆等植物。1960年之后间作开始在中原大地上广泛流行,不同的植物种类也开始逐渐增多[5]。间作在非洲、南美洲等地区广泛流行,近年来欧洲、北美洲也逐渐采用间作代替部分农业生产[6]。
用于间作生产的模式有林‖草间作[7]、蔬菜间作[8]、林‖农间作[9]、农作物间作[10]、农‖草间作[11]、林‖药间作[12]等多种,这些方面均有较多研究报道。目前我国已经开始大面积间作农作物,并获得了可观的收益[13]。研究表明,间作能有效促进植物根系对营养物质的吸收,加快植物生长发育[14]。玉米‖马铃薯间作使产量比单作分别提高5.6%、9.5%,豇豆‖花生间作使产量比单作分别提高30.00%、875%[15-16]。同时,间作体系下植物根际土壤微生物生物量和群落结构丰富度增加,土壤微生物多样性得到提升[17]。这对促进土壤间物质循环[18]、提高植物抗逆性[19-20]、修复污染土地[21]、改善土壤结构[22]以及调节土壤理化性质[23]等有较大帮助,在农业生产中优势明顯。本文综述了近十年来国内外间作对植物、土壤和微生物影响的研究进展,并作出讨论和展望。
1 间作对植物生长和产量的影响
间作能通过合理利用环境资源促进植物生长发育,提高作物产量[24]。玉米‖大豆间作时,玉米增产23.56%~33.64%[25],水稻‖大豆间作可使产量提升33%[26]。此外,间作通过增加作物的多样性提高地面的覆盖度,从而提升水分、光照、土地资源的利用程度,改善土地理化性质,增加作物产量[27-28]。在辽西半干旱的土壤中,玉米‖大豆间作提高了玉米的穗长、穗质量等指标,主要原因是植物地上部对土地的覆盖、发达的根系保持了土壤中的水分,提高了植物的水分利用效率[29]。探索合理的间作模式具有重大意义,枣‖棉花间作模式中过密的行距及过高灌水量均会导致棉花减产[30]。间作大豆产量在行间距2.4 m时达到最高,比行间距为2.0、2.2 m分别提高37.2%和19.6%[31]。在资源利用合理情况下,间作可提高产量,反之则可能面临产量下降的风险。
土地当量比是间作优势的一个关键因素,在增产的间作模式中,其土地当量比均>1。适宜的间作模式下,土地当量比总是要高于单作模式[32]。另外,间作模式中植物地下部的竞争往往更为激烈,这导致了更长的根长、更大的根表面积和更多的分根数,进而提高了土壤水分的利用效率[33-36],是水资源利用效率提高的重要原因。
间作中根系分泌物的种类和含量比单作时显著增高,有利于提高根际土壤酶活性,加强植物根际抗病性和营养物质转移,而这些过程对促进植物生长发育和产量增加具有重要意义[37-39]。间作通过利用不同种植物之间的生态位差异,能高效地利用土壤环境中的水平和垂直层次,并实现植物在空间分配上的立体化利用,提高植物与土地之间空间和养分的利用效率[40-41]。例如,甘蔗‖花生间作[42]、黄菊‖油茶间作[43]、玉米‖花生间作[44]和苹果‖大豆间作[45],通过利用垂直方向上的生态位差异,偏向地上部生长与偏向地下部生长的不同植物间作栽培,能充分利用土壤的上下空间环境。在水平方向的利用上,植物之间的冠幅不受或者少受相互影响,尽量减少因为枝叶叠加对光合作用的干扰。也有研究表明,尽管根据生态位不同能提高资源利用度、减少植物间相互影响,但随着植物生长,这种影响始终存在[46-47]。例如,较高生态位的植物通过竞争阳光,对较矮植物形成一定的荫蔽度,从而削弱较矮植物的光合作用[48]。该现象在成行列种植的间作模式中尤为显著,特别是种植于边行较矮的植物对自然资源的利用度较低,导致产量下降,较高植物的产量则可能因此增加[49-50]。因此,掌握合理的间作模式并充分利用间作规律,对调控植物的生长和产量具有重要影响。
2 间作对光合作用的影响
光合作用途径中固定的碳水化合物最终形成植物的生物量,提高光合作用效率对提升作物生长生产具有显而易见的价值[51]。植物对光能利用的效率不仅取决于植物本身,还受到如光强、温度、水分等自然环境因子和生物化学试剂等人为因子的影响[52-53]。例如,高温和干旱导致了大豆叶片光合作用显著降低,从而导致大豆减产[54];重度涝渍时花生光合作用及生物量均下降[55-56];22种常用农药中有9种对甜椒的光合系统有显著的抑制作用[57];微塑料聚苯乙烯通过改变微生物的代谢和微生物之间的相关性来削弱植物的生长和光合作用[58]。间作通过促进植物根际分泌物产生、调控微生物群落、改善植物-土壤间微环境等方式提升植物的抗逆性和光合作用,净化农药,降解有机物,从而抵御自然环境及人为因子的干扰[59-61]。
这种抵御作用亦能维持种间竞争下作物的生理状态,进而有利于产量增加。大豆‖亚麻垄沟间作试验表明,大豆在受到种间竞争抑制的情况下,光合速率和叶绿素含量仍高于单作[62]。在大豆‖玉米间作后期,光截获面积增加,降低了叶绿素的降解速率,并提高了叶片寿命[63]。Feng等进一步研究发现,叶片寿命提高直接导致土地当量比升高,其中2行大豆2行玉米模式下土地当量比高达1.54[64]。间作也会改变植物响应光合变化的能力,桑树‖谷子间作缩短了桑树“光合午休”时间,并提高了叶片气孔导度和水分利用效率,明显提高了桑树叶片的光合生产能力,缓解了午间的高光照胁迫[65]。带状间作改善了玉米的光响应曲线,提高了玉米对强光的适应性,而降低了大豆的光响应曲线。该研究还发现,带状间作提高了玉米最大荧光(Fm)、光化学淬灭系数(qP)的值[66]。合理的间作模式能提升植物对光的利用能力,维持较好的生理状态,提高植物产量。
3 间作对土壤的影响
3.1 间作对土壤含水率、pH值和有机质的影响
土壤含水率反映土壤中水分含量的多少,间作通过影响植被密度、植物群落多样性、土壤孔隙度等调控土壤含水率。在埃塞俄比亚南方地区一项抗旱试验中,研究者发现玉米‖紫云英间作具有较高的土壤含水率和产量[67],有利于保持旱地土壤水分,对植物起到促生效果。也有研究表明,土壤含水率通过影响土壤呼吸和养分有效性而间接影响整个间作系统[68-69]。土壤pH值主要受土壤中酸碱性物质的影响,能够通过影响土壤养分的有效性改变土壤肥力,进而对植物生长发育过程起到间接作用[70]。酸性土壤团聚体稳定性较低,会導致土壤有机质淋失增加,降低土壤肥力[71]。土壤pH值对不同元素有效性的影响不同,钾、铁、锌、硼、铜离子在酸性条件下有效性较高,而磷离子在中性条件下有效性最高[72-73]。有研究表明,单作蚕豆根际土壤pH值显著下降,但间作缓解了下降趋势[74]。秦昌鲜等研究发现,甘蔗‖花生间作根际土壤pH值下降,有效磷含量上升,土壤pH值和有效磷之间存在着显著的负相关,可能是间作土壤酸化导致的。上述研究表明,间作对土壤pH值的调控也受到植物因素的影响[75]。
土壤有机质是以各种形式和状态存在于土壤中的含碳有机化合物,其中腐殖质是有机质的主体[76]。土壤有机质含量主要受物质输入和分解过程的共同影响,与土壤肥力、植物根系分泌物、植物营养吸收、土壤微生物群落密切相关[77-79]。Zhang等的研究表明,梨‖芳香植物、甘蔗‖花生间作均可提高土壤有机质含量,进而提高土壤肥力[80-81]。Nyawade等研究发现,马铃薯‖金甲豆、马铃薯‖紫花扁豆2种间作模式都可以提高土壤中可溶性有机质含量,进而避免单作马铃薯连种对土壤有机质的恶化效果[82]。Zhang等则发现间作对土壤有机质含量的影响可能与土壤微生物群落的组成发生改变有关[80]。更多的研究表明,间作对土壤有机质含量具有双重影响。一方面,间作通过提高土壤微环境生态效应促使动植物残体分解,提高土壤有机质含量;另一方面,植物通过竞争土壤养分,加快土壤有机质矿化分解和养分循环,从而降低有机质含量[82-83]。
3.2 间作对土壤大量元素的影响
大量元素氮磷钾素是生态系统中至关重要的营养元素,一切生命过程以及地球养分循环都离不开大量元素[84]。氮素是构成生命最基本的元素,也是蛋白质、核酸等生物体必需物质的重要组分,直接参与整个生命活动过程[85]。间作豆科植物可以通过根瘤菌的固氮作用改善土壤氮素含量,并为其他植物提供氮源[86]。马铃薯与豆科植物间作,可以补给土壤中马铃薯消耗的氮素,达到氮素平衡的目的[87]。有研究进一步表明,豆科‖非豆科间作模式下植物吸氮量显著高于单作[88]。此外,通过氮素从豆科植物根区向非豆科植物根区的转移过程还可以进一步刺激豆科植物根瘤菌的固氮作用[89]。
磷素是氨基酸的重要组分,其含量高低直接影响植物代谢过程,对改良作物品质具有较好的效果[90]。间作能降低磷肥的添加量,通过综合分析现有豆科‖禾本科间作资料发现,在相同产量条件下,间作作物比单作作物需磷量低约21%[91]。间作能有效提高作物对磷素的吸收量,谷类‖豆类间作可大幅度提高土地利用效率和磷素吸收,从而提高作物产量。Latati等研究发现,碱性土壤中豇豆‖玉米间作提高了磷素有效性和玉米产量[92]。植物磷素吸收量与产量具有相关性,在马达加斯加中部高地进行的一项研究中显示,通过植株磷素吸收量可以较好地预测产量[93]。大量研究表明,磷素含量与养分吸收量及植物发育状态有较强的正相关性,可能的原因是磷素通过影响作物根系发育和羧酸盐分泌等调控作物对土壤养分的吸取,并且这个过程具有正向反馈[94-95]。间作模式具有典型的正向反馈作用,植物对磷素的竞争能明显提高磷素吸收量,并提高根际土壤磷酸酶活性,进而促进土壤中难溶性磷的活化,缓解竞争[96]。此外,有研究表明植物对氮素和磷素的吸收具有一定的同步性,氮素含量增加能促进磷素的吸收[97]。由此可见,间作豆科植物能发挥较好的磷素活化利用优势。
钾素与植物体内多种酶的活性密切相关,能够参与蛋白质积累、光合作用和细胞信号传导[98],并增强植物的抗逆性[99]。间作模式下钾素具有和磷素同样的正反馈作用,从而提升植物钾贮量和土壤钾含量,这种反馈效应在枸杞‖禾本科、茶‖板栗、玉米‖大豆、花生‖甘蔗等间作模式中均被观察到[100-103]。在氮素添加后,植物中钾含量也相对增加,氮与钾的比值可以反映土壤对植物养分供给与限制状况[104]。豆科‖非豆科间作时,往往观察到钾素含量上升。在中国热带农业科学院橡胶园进行的一项间作试验表明,橡胶树与9种豆科植物间作均不同程度地增加了土壤全钾和速效钾含量,并且增加了速效氮含量,提高了土壤供氮、供钾能力[105]。一个重要的原因是,土壤中氮素含量的升高促进了交换性钾饱和度的提高,从而提升了植物对钾素的吸收[104]。以上研究表明,间作在促进植物对氮、磷、钾素的利用方面表现突出。
3.3 间作对土壤中、微量元素的影响
中、微量元素广泛参与植物体内生长生理、代谢等过程,对维持植物体内部稳态起到重要作用[106]。植物中的钙、镁可以参与生理生化反应,增强植物的抗逆性[107-108]。锌、铁、锰参与调控光合作用过程,从而促进植物生长[109-111]。硼是高等植物生长发育所必需的营养元素,缺硼会严重影响叶片形态,但是过量硼会抑制植物生长[112]。Vinichuk等研究发现,间作提高了作物中锌、锰的含量,从而改善了植物生长状况[113]。李娟等研究发现,胶园土壤和根际土壤中微量元素存在广泛变异,其中铁、锰含量达到丰或很丰水平[114]。Cao等使用超富集植物景天和油菜间作,发现间作显著提高了镉的植物提取率和油菜的生物量[115]。Rasmussen等使用示踪法探究干旱和间作对菊苣养分吸收的影响,发现菊苣在3.5 m深度吸收氮,而在2.3 m深度吸收微量元素;同时,间作模式下出现了补偿吸收,说明植物种间竞争改变了养分吸收模式[116]。也有研究发现,间作通过提高微生物数量和多态性促进对土壤中微量元素的活化吸收,从而提高植物中相應元素的含量[117-118],接种某些特定菌群亦能强化间作模式对中微量元素的吸收[119]。以上研究表明,间作通过植物种群间互作来改善土壤微生物数量结构和功能,从而影响植物对中微量元素吸收利用的模式。
4 间作对土壤酶的影响
土壤酶在维持土壤生态平衡方面起到重要作用,是土壤生态系统功能的指示剂[120]。土壤酶也是土壤中的生物催化剂,推动土壤中物质转化、养分循环、有机质矿化和有机物降解,对改善土壤理化性质有着重要的作用[121]。土壤酶主要来自微生物、植物根际和部分动物,微生物是土壤酶产生的主要来源,其次是植物根际活化土壤所分泌的酶;某些动物,如蚯蚓等也会产生少量胞外酶[122]。种植模式会对土壤酶活性产生影响,间作模式下植物具有较高产量和养分吸收量,这种优势进一步影响土壤酶活性[123]。例如玉米分别与红薯、向日葵、花生间作,明显提高了脲酶、过氧化氢酶和磷酸酶活性[124];玉米‖大豆间作模式下,根际土壤中脲酶和磷酸酶活性明显提升,细菌和放线菌数量均显著高于单作[125]。大蒜‖小麦间作时,小麦的根际土壤中磷酸酶和脲酶活性在整个生长阶段都明显高于单作,在部分生长阶段过氧化氢酶的活性也显著提高[126]。在以往的研究中,不同种植方式和土壤肥力对于土壤酶活性影响的研究较多。
目前,在土壤中发现50多种与生物过程有关的酶类。间作系统中,水解酶和氧化还原酶扮演着重要角色。水解酶能水解多糖、蛋白质、难溶性有机磷等大分子物质,使其形成小分子物质,更利于植物的吸收;同时,水解酶能够改善土壤结构,促进碳、氮、磷元素的转化过程,从而保证地球生态系统内养分循环[127-128]。氧化还原酶参与生物化学代谢和凋落物分解等过程,与能量的获得与释放有关,与水解酶起着同样重要的作用[129]。在大多数研究中,水解酶和氧化还原酶能较好地解释间作机理,其中脲酶、蛋白酶、磷酸酶、蔗糖酶、纤维素酶、脱氢酶和过氧化氢酶最为常见。
脲酶是一种重要的水解酶,能够促进尿素水解为NH+4和CO2,为植物提供可以直接吸收的氮,促进植物生长[130]。花生‖茶叶间作显著提高了0~40 cm不同剖面土层脲酶活性,且脲酶活性与有机碳、全氮显著相关[131]。在茶‖板栗间作试验中同样发现,间作使土壤氮素含量和脲酶活性得到了显著提高[132]。
蛋白酶属于水解酶,可以将土壤中蛋白质、肽类分解为氨基酸,以便植物直接吸收利用[133]。蛋白酶参与土壤氮素循环过程,促进有机质分解并释放氨基酸,可作为土壤氮素含量的指示剂[134]。间作对土壤蛋白酶活性的影响随植物种类发生改变,王玉新等研究发现,薰衣草与萝卜、白菜分别间作均会降低土壤蛋白酶活性[135]。颜彩缤等的研究表明,槟榔‖平托花生间作提高了土壤蛋白酶活性[136]。
磷酸酶与磷素生物转化有关,是在生物界普遍存在的一种水解酶[137]。磷酸酶通过活化有机磷,将其转化为无机磷供植物利用。一般情况下,土壤中磷酸酶含量越高,土壤中磷素含量和利用效率也越高[138]。小麦‖豌豆间作模式下,土壤磷酸酶活性和小麦吸磷量比单作、施磷均较大,且间作在低磷时比高磷条件下更有利于小麦磷素积累[139]。也有研究表明,低磷时间作豆科植物可通过诱导根瘤中植酸酶和酸性磷酸酶活性,刺激根瘤固氮效果,从而具有更高的固氮效果[140]。以上研究说明间作能在低磷时有效促进磷素的活化,而在高磷时降低磷酸酶活性以调控土壤养分结构。
蔗糖酶是一种水解酶,它可将蔗糖水解为葡萄糖供微生物直接吸收,与土壤微生物活动密切相关[141]。相关研究表明,黄花菜分别与西瓜、白菜、甘蓝间作时土壤中蔗糖酶活性均高于单作,同时各间作模式下微生物群落结构也发生了明显改变[142]。
纤维素酶属于氧水解酶,可以分解纤维素为小分子糖分,为微生物生命活动提供能源物质[143]。生姜与四季葱、大蒜、大豆间作均可提高土壤纤维素酶活性[144],且间作与施肥的交互影响能显著提高土壤中纤维素酶活性[145]。但纤维素酶属于根系细胞壁降解酶,其活性过高可能会破坏根系细胞结构,进而导致病菌侵入、根系腐烂。袁婷婷等通过研究接种病原菌的间作蚕豆根系发现,蚕豆根际土壤纤维素酶活性降低,病原菌侵入率也随之大大降低[146]。说明间作可以调控并改善间作土壤环境,为植物生长发育提供一定的保障。
土壤脱氢酶属于氧化还原酶,是微生物降解有机物机制中的重要因素之一,可以减少有机物对土壤的污染,降低对植物的毒害作用[147]。间作可以通过提高脱氢酶活性或者微生物丰度等方式促进土壤有机物降解,从而改善土壤环境。大豆分别间作桑树和玉米均提高了土壤脱氢酶活性[148-149]。研究表明苹果园间作苜蓿的模式中,脱氢酶是显著影响土壤微生物生物量的因子之一[150]。代真林等研究发现,玉米‖大豆间作土壤中降解有机物的有益微生物丰度增加[151]。
过氧化氢酶属于氧化还原酶,能够分解生物过程中产生的H2O2,减缓H2O2对植物根系造成的氧化损伤,在土壤中分散较广[152]。和单作相比,葡萄間作黄波斯菊、凤仙花均可显著提高土壤过氧化氢酶活性[153]。黄瓜分别与洋葱、大蒜间作时,也发现了土壤过氧化氢酶活性的提高[154]。
5 间作对土壤微生物的影响
土壤微生物的分解效应能加速植物对营养物质的吸收,同时解体土壤团块,疏通土壤孔隙,促进土壤内部气体交换,进而有利于植物根系的呼吸作用。间作体系中,不同植物种间竞争与促进效应常常会丰富土壤中微生物群落多样性,微生物数量增加的同时也会反向调节植物生长,在合适的间作模式下二者之间能够达到互惠互利、共同促进的效果[155]。因此,土壤微生物数量变化是最能反映土壤状况的一项指标,也与间作模式对土壤活性的影响有关。茶树‖大豆间作模式下细菌、放线菌、真菌数量均上升,同时微生物生物量、多样性指数也得到提高[156-158]。研究表明,金橘‖大豆间作模式下植物根际土壤微生物生物量与根系长度显著增加[159];小麦‖蚕豆间作下土壤细菌、真菌、放线菌数量加权平均后均下降,而间作对植物生长促进作用和微生物多样性均提升,说明间作可促进植物与微生物间互相作用达到有机平衡[160]。
间作方式能影响植物生长状况、根系分泌物、土壤结构等关键因子,从而影响微生物群落结构和多样性。通过不同间作试验可探究最适宜微生物的土壤环境,实现最大化经济与收益的平衡。李梦辉等研究发现,香蕉/辣椒间作对生物量、土壤养分含量和土壤酶活性影响显著,同时发现根系的分隔方式对不同种类土壤微生物数量产生了显著影响[161]。张海春等、吕慧芳等也得到了类似的研究结果[162-163]。张亮亮等研究发现,枣树‖棉花间作在土壤0~60 cm土层垂直方向上微生物量碳、氮迅速下降;微生物量碳、氮与土壤指标有显著的相关性[164]。黑豆‖燕麦间作下微生物生物量、碳含量均较单作得到提高,但是氮含量较黑豆单作时有所降低[165]。马琨等利用不同品种马铃薯间作栽培试验后得出土壤微生物氮含量升高,碳含量下降,使得土壤碳氮比呈现下降的趋势。同时发现,不同品种马铃薯间作后细菌种数比单作显著增加,真菌和放线菌数量降低[166]。而牛宇等则研究发现,樱 桃‖绿豆间作土壤放线菌、细菌数量无显著变化,大樱桃‖豇豆间作土壤细菌数量呈极显著减少[167]。
间作能够改变微生物对碳源的利用模式,进而引发微生物群落结构发生改变。马琨等研究发现,玉米‖马铃薯间作模式下,以多聚类化合物、芳香类化合物、羧酸类、氨基酸类为碳源的微生物种群活性增加[166]。而蚕豆‖马铃薯间作系统下,促进了以碳水化合物为碳源底物的微生物种群的形成,同时也增加了土壤微生物总量。杜春风使用玉米‖马铃薯间作3年后得出土壤每孔颜色平均变化率(AWCD值)整体高于单作,且提高了土壤微生物对羧酸类、多聚化合物类碳源的利用能力[168]。至于对芳香类化合物、氨基酸类碳源的利用无明显变化,是因为间作达到一定时间后土壤理化性质、土壤肥力以及土壤微生物群落结构较之前发生了很大的变化。张萌萌等在桑树‖苜蓿间作试验中发现间作桑树根际土壤微生物AWCD值、均匀度指数、优势度指数、多样性指数均高于单作桑树,而苜蓿根际土壤则呈现相反的趋势[169]。同时,桑树根际土壤微生物对碳源的利用活性提高,而苜蓿根际土壤微生物则改变了对碳源的活化利用类型。逄好胜等通过进一步试验,发现羧酸类碳源是引起桑树‖苜蓿间作土壤微生物群落功能差异的主要碳源,该试验为进一步分析桑树‖苜蓿土壤微生物群落功能多样性奠定了较好的理论基础[170]。
较多研究表明,间作推动了根际微生物群落结构的转变,有益菌群比例变得更大。白鹏华等研究发现,枣树‖绿豆间作后枣树根际土壤细菌数量上升的同时真菌数量下降,土壤微生物群落构成逐渐由真菌型转变为细菌型[171]。该间作模式下部分真菌被认为是一种病原菌群,它们在土壤中数量的下降表明土壤的生态结构得到了改善,即枣树‖绿豆间作对土壤生态系统具有一定的有机调控作用。但是,具体的影响仍然无法准确测得,推测这可能与微生物之间、微生物与土壤环境之间复杂的作用关系所导致。对于蔬菜间作,夏枫研究发现,苦瓜与大白菜、芥菜、葱、大蒜分别间作下有益真菌拟青霉属(Paecilomyces)相对丰度增加,且苦瓜‖大蒜间作下木霉属(Trichoderma)、毛壳菌属(Chaetomium)等有益真菌相对丰度显著增加[172]。以上研究说明,部分间作模式在促进有益真菌而抑制有害真菌生长方面有较好的效果,这对植物生长发育起到了良好的改善和调控作用。
同种植物分别与其他植物间作为研究最适间作模式提供了一条思路,张海春等使用番茄和生菜、芹菜分别间作,发现土壤的酶活性、生物量、植物产量显著提高,克服了番茄的连作障碍[162]。董宇飞等进一步研究发现,使用3年辣椒‖玉米、辣椒‖大蒜间作与辣椒连作相比,细菌、放线菌数量显著增加,而真菌数量显著减少[173]。该研究同时发现土壤中细菌优势种群变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria),真菌优势种群子囊菌门(Ascomycota)、担子菌门(Basidiomycota)等有益菌相对丰度提高,有益于土壤中有益微生物数量的增加。谭礼宁还证实了花生‖谷子间作土壤微生物种群的结构发生了改变,螺原体属(Spiroplasma)等有害菌数量减少,链霉菌科(Streptomycetaceae)和芽孢杆菌属(Bacillus)等有益菌数量增多[174]。牛宇等也发现玉米‖马铃薯间作改变了根际真菌的种、属数,优势种由明球囊霉菌(Glomus clarum)、地球囊霉菌(Glomus geosporum)转变为福摩萨球囊霉菌(G.formosanum)和球泡球囊霉菌(G.globiferum),结果表明土壤微生物种类结构发生了改变[167]。邓文等也得到了类似的研究结果[175]。以上研究进一步印证了间作对改善土壤微生物种群类型和群落结构具有显著性的效果。部分土壤微生物对间作体系的反馈作用能提高整体的环境抵抗能力,乔旭发现丛枝菌根真菌(arbuscular mycorrhizal fungi,AMF)能抑制小麦‖蚕豆间作系统中的杂草生长,保证作物的正常生长发育[176]。土壤病原微生物会抑制单作小麦的生长,但对小麦‖蚕豆间作系统的植物生长无影响,说明小麦‖蚕豆间作能克服土壤病原微生物对植物生长的负效应,这与王光州的研究结果[177]相似。筛选并利用优良的间作模式进行农业生产,是一项可持续的生态产业。
6 植物-土壤-微生物互作提高间作产量
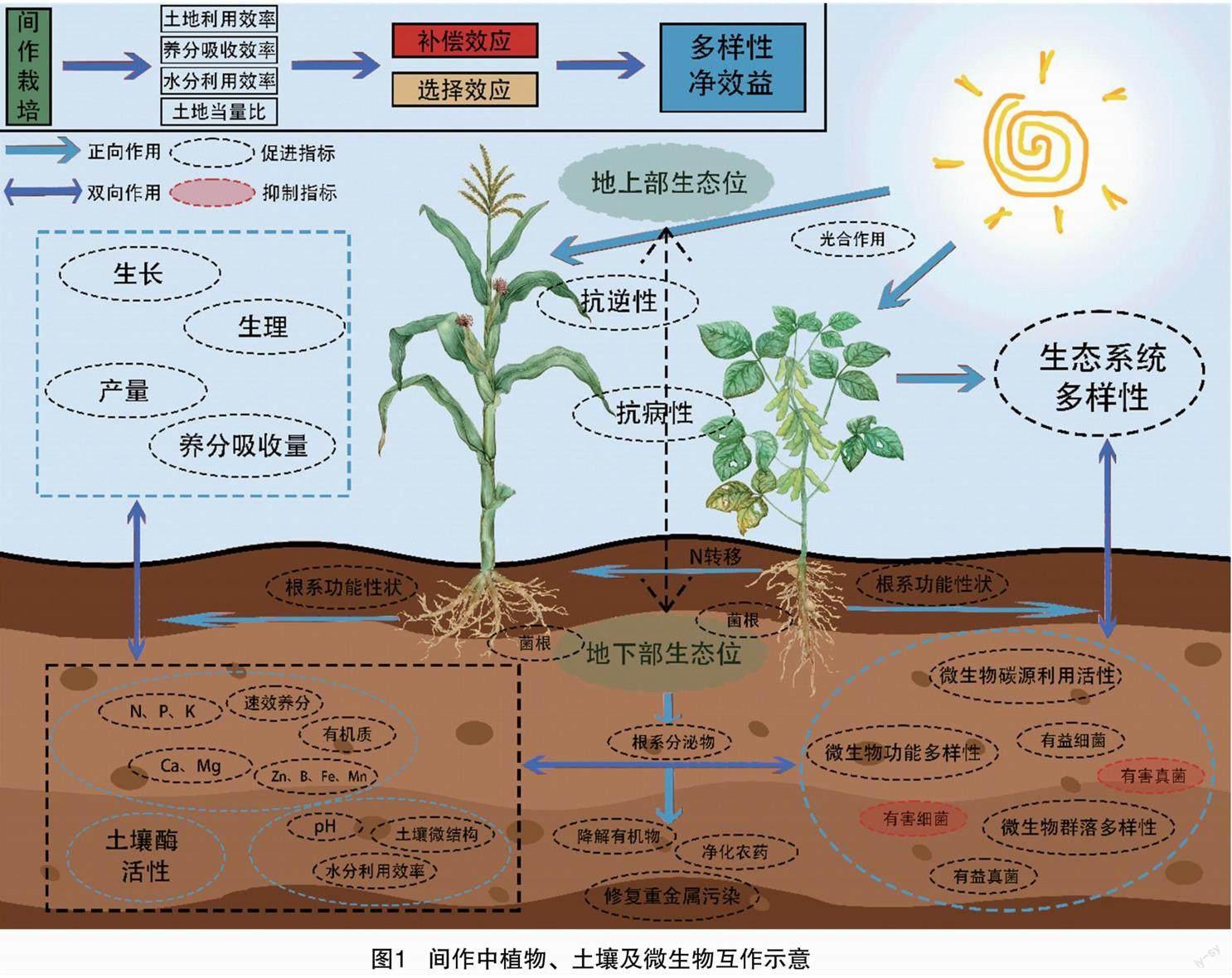
间作具有土地利用效率、养分吸收效率、水分利用效率和土地当量比优势,从而形成高產。在间作系统中,间作优势主要由2个方面决定,一方面是补偿效应,即植物间通过生态位互补从而提高资源利用,促进生产力增加;另一方面是选择效应,即由优势植物主导的产量增加。补偿效应和选择效应共同组成多样性净效益,即间作模式下生产力的增加量。多样性净效益的增加与间作系统中植物、土壤和微生物互作过程密不可分。
间作通过地上部生态位提高植物对光资源的利用能力,并协同地下部生态位提高植物抗逆性和抗病性。菌根、根系分泌物均有助于有机物降解、净化农药及重金属污染修复。在经典的间作豆科植物模式中,豆科植物通过根瘤菌固氮作用,进一步将氮素转移至非豆科植物根系周围,以提高氮素利用效率。间作中,植物根系通过促进有益微生物(如细菌和真菌)、提高微生物碳源利用程度、增加微生物群落多样性并提高功能多样性,调控植物生长发育;同时抑制有害微生物,改良土壤、创造生态效益。间作模式下植物根系功能性状的改变亦可改良土壤,提高土壤理化性质平衡性,分解有机物,改善土壤微结构,提高养分含量和酶活性。较好的土壤条件正向促进植物生长、生理、养分吸收和产量的提升,体现出较好的多样性净效益,使得间作系统整体产量提高(图1)。
7 讨论与展望
7.1 间作的生态和经济效益
近10年来,国内外大量间作研究均取得了较好的成效,从植物、土壤和微生物等方面证实了良好的间作模式具有可持续的生态效益。间作符合未来人类生态农业的发展方向,符合人类社会生态健康发展的切身利益。目前,国内外已经对玉米、花生、大豆、棉花、枣、甘蔗、谷子、桑树、苜蓿等常见农粮作物进行广泛的间作栽培试验,主要研究结论在于产量增加、土地利用率提高、节约肥料等,具有客观的生态和经济效益。但由于全球地理环境差异巨大,对不同作物适应生长带来明显影响,不同区域主要经济作物不同,未来应当考虑以当地经济作物为主,并结合成熟的农艺养护管理措施,确保筛选出最适合该区域的间作栽培模式。
7.2 间作研究的重点方向
通过合理利用不同植物的生态位差异,探索植物互惠互益的多样化间作模式,仍然是未来间作产业研究的重点。此外,由于植物间生态位不同,对于间作作物采收时也可依据不同位置设置不同的采收机械,以提高效率。在未来间作大面积推广的背景下,农业机械的发展大潮也会随之涌出,对于传统机械的改进与创新亦是未来发展的一个重点。
由于长期连作,较多土地理化性质恶化严重,通过间作的方法减缓土地恶化程度,甚至改良土壤、提高理化性质是一种新的生态种植手段。既缓解了土地恶化,又增加了区域物种多样性,维护生态系统健康与平衡。目前也有大量研究间作富集植物对重金属的吸收,在治理矿区、河流、工厂等污染区域有显著效果,随着社会的发展,工业化遗留问题也可通过生态间作的方式进行改善。从长期来看,间作可成为工业景观的一部分,能将不同的景观生境加以联接。
7.3 间作深层次机制的探究
良好的植物种间配置模式是探究间作机制的基础,对光照、水分、温湿度、气体排放、土壤等因素的测定是了解间作模式的必要手段。环境因子变化是导致间作产量改变的主要原因,对于同一间作模式而言,在不同环境因子影响下植物生长情况并不完全一致,产量各异。此外,植物对环境的适应能力也因品种差异而不尽相同,需要大量基础研究以探讨最适间作模式。
土壤因子亦不可忽视,土壤中大量元素是植物生长发育所需最主要元素,大部分研究也主要集中于间作模式下大量元素的变化情况。现有研究资料表明,间作对中微量元素含量具有重要的影响,但目前国内外对间作如何影响土壤中微量元素含量变化的机制研究尚少见,且关注点多在利用富集植物间作修复或者改善矿质污染的土地,提高土地利用率。说明该研究方向没有得到足够重视,仍有存在科学探索的可能性。例如,间作是如何通过促进植物对中微量元素的高效吸收,进而改善相关生理过程和代谢水平的?间作模式下中微量元素的吸收利用效率与作物品质有何关联?使用示踪法标记土壤中微量元素在植物吸收转运及利用过程或是一个较为成功的研究方法,能更清晰地展示间作模式下植物对元素的竞争吸收与活化利用状况。
目前间作机理也较多集中于植物生长和对养分的吸收利用状况,也有研究分析了土壤酶发生的变化。究其深层次的因素,如该间作模式下土壤-土壤酶-土壤微生物互作情况的研究尚少见, 绝大部分研究仅停留在土壤或土壤-土壤酶层面。因此,对于间作系统的研究应当注重各环节的整体性分析,未来的研究也应当尽量展现整个间作系统,从环境因子到植物,再到土壤-微生物,越是全面的研究,越有利于间作机理的发掘,促进间作产业的发展。
7.4 间作产业面临的瓶颈
目前大部分地区农业生产仍以单一作物连作为主,间作模式仍需大力推广,以覆盖更多的土地。一个重要的原因是,间作总体持续时间较长、产生经济效益所需时间久, 相比之下连作当年生作物能带来迅速的利润,部分农民不愿意投入更多的时间成本进行间作管理。此外,间作模式自身的特点造成了田间管理和作物采收上的不便,在规模化农作物采收时更是缺乏相应工具,以至于面临较高的人力成本。
在研究成果落地方面,目前大量间作研究仅停留在理论水平,在实际农业生产上运用较少,一方面是由于知识产权形成和成果转化的进度仍需要加快;另一方面是间作模式的推广力度仍需要加强。通过以政府统筹、规划、补贴为导向的政策逐步带动传统农业向生态间作农业转型,或以试点农业模式带动区域农业,可以起到范式作用,引导突破瓶颈。
7.5 高通量测序技术在间作研究中的应用
随着科学技术不断成熟,高通量测序技术推进了有关土壤微生物结构、功能和物种多样研究的发展。国内外已有较多研究利用高通量测序技术对间作土壤样本进行分析,以探究土壤微生物群落的构成和变化情况,该技术的进一步完善有利于土壤微生物的生态研究。利用高通量测序技术对间作土壤进行分析,探究土壤微生物群落与植物生长和产量变化的影响机制,能够加深我们对间作系统的准确认识,是一条快捷、方便的新途径。
目前使用高通量测序技术探究土壤微生物群落结构和组成多样性的研究较多,但精准定位关键微生物种类、深入发掘目标微生物群落特定功能的研究较少。在间作系统中,关键微生物群落是什么,发挥了什么样的功能?间作改变微生物群落组成会对生态系统造成什么样的长期影响?通过微生物群落改良能否进一步提高间作系统生态效益和生产力以促进解决粮食问题?目前对于间作模式中微生物的研究仍需要与实际生产更紧密地结合起来,分析出何种条件下哪些核心微生物群落占主导作用,并构建植物-土壤-微生物互作体系,是能通过高通量测序解决的一项重要科学问题。
参考文献:
[1] 董宛麟,张立祯,于 洋,等. 农林间作生态系统的资源利用研究进展[J]. 中国农学通报,2011,27(28):1-8.
[2]贺 佳,安曈昕,韩学坤,等. 间作群体生态生理研究进展[J]. 作物杂志,2011(4):7-11.
[3]Schmidt O,Curry J P.Effects of earthworms on biomass production,nitrogen allocation and nitrogen transfer in wheat-clover intercropping model systems[J]. Plant and Soil,1999,214:187-198.
[4]崔爱花,刘 帅,白志刚,等. 间作对旱地作物生长发育及生理生态影响的研究进展[J]. 中国农学通报,2021,37(18):1-5.
[5]方旭飞. 种植模式和覆盖方式对土壤水分和玉米生长指标的影响及效益分析[D]. 沈阳:沈阳农业大学,2018:7-10.
[6]李 美. 玉米花生间作群体互补竞争及防风蚀效应研究[D]. 沈阳:沈阳农业大学,2012:6-18.
[7]吴永波,吴殿鸣,薛建辉,等. 杨树-冬小麦间作系统细根分布特征及对施氮的响应[J]. 生态与农村环境学报,2015,31(3):320-325.
[8]Laffray X,Toulab K,Balland-Bolou-Bi C,et al. Evaluation of trace metal accumulation in six vegetable crops intercropped with phytostabilizing plant species,in a French urban wasteland[J]. Environmental Science and Pollution Research,2021,28(40):56795-56807.
[9]Xue Z L,Wang Y L,Yang H J,et al. Silage fermentation and in vitro degradation characteristics of orchardgrass and alfalfa intercrop mixtures as influenced by forage ratios and nitrogen fertilizing levels[J]. Sustainability,2020,12(3):871.
[10] Salgado G C,Ambrosano E J,Rossi F,et al. Biological N fixation and N transfer in an intercropping system between legumes and organic cherry tomatoes in succession to green corn[J]. Agriculture, 2021,11(8):690.
[11]Hu S,Liu L J,Zuo S F,et al. Soil salinity control and cauliflower quality promotion by intercropping with five turfgrass species[J]. Journal of Cleaner Production,2020,266:121991.
[12]童炳麗,刘济明,熊 雪,等. 米槁-绿壳砂林药间作系统可行性[J]. 生态学报,2019,39(15):5690-5700.
[13]王丽杰. 玉米与大豆间作高产栽培技术[J]. 种子科技,2019,37(15):56,59
[14]Zhao X H,Dong Q Q,Han Y,et al. Maize/peanut intercropping improves nutrient uptake of side-row maize and system microbial community diversity[J]. BMC Microbiology,2022,22(1):14.
[15]金建新,何进勤,冯付军,等. 马铃薯/玉米间作对作物生理生态特性的影响[J]. 贵州农业科学,2019,47(5):14-19.
[16]邹晓锦,刘子琪,牛世伟. 花生与豇豆间作体系节肥增效试验初报[J]. 南方农业,2019,13(32):3-4.
[17]Liu C Z,Cai Q Z,Liao P R,et al. Effects of Fallopia multiflora-Andrographis paniculata intercropping model on yield,quality,soil nutrition and rhizosphere microorganisms of F. multiflora[J]. Plant and Soil,2021,467(1):465-481.
[18]Nadir S,Othieno C,Kebeney S. Nutrient dynamics in Eucalyptus plantations of different ages before and during intercropping[J]. International Journal of Plant & Soil Science,2018,22(1):1-13.
[19]Bayala R,Diedhiou I,Bogie N A,et al. Intercropping with Guiera senegalensis in a semi-arid area to mitigate early-season abiotic stress in A.hypogea and P.glaucum[J]. Journal of Agronomy and Crop Science,2022,208(2):158-167.
[20]Liu H M,Gao Y,Gao C Q,et al. Study of the physiological mechanism of delaying cucumber senescence by wheat intercropping pattern[J]. Journal of Plant Physiology,2019,234/235:154-166.
[21]Yan Y X,Yang J,Wan X M,et al. Temporal and spatial differentiation characteristics of soil arsenic during the remediation process of Pteris vittata L. and Citrus reticulata Blanco intercropping[J]. Science of the Total Environment,2022,812:152475.
[22]Chen C F,Liu W J,Wu J N,et al. Can intercropping with the cash crop help improve the soil physico-chemical properties of rubber plantations?[J]. Geoderma,2019,335:149-160.
[23]Zhang X L,Teng Z Y,Zhang H H,et al. Nitrogen application and intercropping change microbial community diversity and physicochemical characteristics in mulberry and alfalfa rhizosphere soil[J]. Journal of Forestry Research,2021,32(5):2121-2133.
[24]Gong X W,Dang K,Liu L,et al. Intercropping combined with nitrogen input promotes proso millet (Panicum miliaceum L.) growth and resource use efficiency to increase grain yield on the Loess Plateau of China[J]. Agricultural Water Management,2021,243:106434.
[25]吕 雯. 玉米‖大豆复合群体耗水特征与生长对灌水量的响应[D]. 杨凌:西北农林科技大学,2021:102-103.
[26]Shah T M,Tasawwar S,Bhat M A,et al. Intercropping in rice farming under the system of rice intensification—an agroecological strategy for weed control,better yield,increased returns,and social- ecological sustainability[J]. Agronomy,2021,11(5):1010.
[27]Koskey G,Leoni F,Carlesi S,et al. Exploiting plant functional diversity in durum wheat–lentil relay intercropping to stabilize crop yields under contrasting climatic conditions[J]. Agronomy,2022,12(1):210.
[28]Singh R,Chaudhary R,Somasundaram J,et al. Impact of crop covers on soil properties,runoff,soil-nutrients losses and crop productivity in vertisols of central India[J]. Indian Journal of Soil Conservation,2014,42(3):268-275.
[29]劉 洋,董 智,董 俊,等. 辽西半干旱区不同玉米间作花生模式对作物产量和水分利用效率的影响[J]. 辽宁农业科学,2022(1):8-12.
[30]王 飞,陈 旭,万素梅,等. 枣棉间作行距和灌水量对棉花干物质及产量的影响[J]. 江苏农业科学,2021,49(9):70-74.
[31]王 甜,庞 婷,杜 青,等. 田间配置对间作大豆光合特性、干物质积累及产量的影响[J]. 华北农学报,2020,35(2):107-116.
[32]Khanal U,Stott K J,Armstrong R,et al. Intercropping—evaluating the advantages to broadacre systems[J]. Agriculture,2021,11(5):453.
[33]Gurin A,Rezvyakova S,Revin N.Nutritional regime of the soil and growth activity of the apple tree root system in orchards with legume- cereal grass intercropping[J]. E3S Web of Conferences,2021,247:01029.
[34]Ma L S,Li Y J,Wu P T,et al. Effects of varied water regimes on root development and its relations with soil water under wheat/maize intercropping system[J]. Plant and Soil,2019,439(1/2):113-130.
[35]Gong X W,Dang K,Lv S M,et al. Interspecific root interactions and water-use efficiency of intercropped proso millet and mung bean[J]. European Journal of Agronomy,2020,115:126034.
[36]Mushagalusa G N,Ledent J F,Draye X.Shoot and root competition in potato/maize intercropping:effects on growth and yield[J]. Environmental and Experimental Botany,2008,64(2):180-188.
[37]Gao X A,Wu M,Xu R N,et al. Root interactions in a maize/soybean intercropping system control soybean soil-borne disease,red crown rot[J]. PLoS One,2014,9(5):e95031.
[38]Jiang Y Y,Zheng Y,Tang L,et al. Rhizosphere biological processes of legume//cereal intercropping systems:a review[J]. Journal of Agricultural Resources and Environment,2016,33(5):407-415.
[39]Fu X P,Wu X,Zhou X G,et al. Companion cropping with potato onion enhances the disease resistance of tomato against Verticillium dahliae[J]. Frontiers in Plant Science,2015,6:726.
[40]Feng C,Sun Z X,Zhang L Z,et al. Maize/peanut intercropping increases land productivity:a meta-analysis[J]. Field Crops Research,2021,270:108208.
[41]Pivato B,Semblat A,Guégan T,et al. Rhizosphere bacterial networks,but not diversity,are impacted by pea-wheat intercropping [J]. Frontiers in Microbiology,2021,12:674556.
[42]唐秀梅,蒙秀珍,蔣 菁,等. 甘蔗间作花生对不同耕层土壤微生态的影响[J]. 中国油料作物学报,2020,42(5):713-722.
[43]古 龙,夏翩翩,李建安.黄菊对油茶林2种杂草萌发和幼苗生长的化感作用[J]. 经济林研究,2020,38(2):34-45.
[44]Zhang W P,Gao S N,Li Z X,et al. Shifts from complementarity to selection effects maintain high productivity in maize/legume intercropping systems[J]. Journal of Applied Ecology,2021,58(11):2603-2613.
[45]李超楠,王若水,周 宣,等. 滴灌水肥调控对苹果-大豆间作系统光合特性和水分利用的影响[J]. 水土保持学报,2020,34(3):299-310.
[46]Zhang W P,Liu G C,Sun J H,et al. Temporal dynamics of nutrient uptake by neighbouring plant species:evidence from intercropping[J]. Functional Ecology,2017,31(2):469-479.
[47]Yang Y,David M,Tjeerd J M,et al. Robust increases of land equivalent ratio with temporal niche differentiation:a meta-quantile regression[J]. Agronomy Journal,2016,108(6):2269-2279.
[48] Wang R N,Sun Z X,Bai W,et al. Canopy heterogeneity with border- row proportion affects light interception and use efficiency in maize/peanut strip intercropping[J]. Field Crops Research,2021,271:108239.
[49]Wang Y Q,Zhao Z G,Li J P,et al. Does maize hybrid intercropping increase yield due to border effects?[J]. Field Crops Research,2017,214:283-290.
[50]Singh S P.Studies on spatial arrangement in sorghum-legume intercropping systems[J]. The Journal of Agricultural Science,1981,97(3):655-661.
[51]Levine R P.The mechanism of photosynthesis[J]. Scientific American,1969,221(6):58-72.
[52]Kumarathunge D P,Medlyn B E,Drake J E,et al. Acclimation and adaptation components of the temperature dependence of plant photosynthesis at the global scale[J]. The New Phytologist,2019,222(2):768-784.
[53]Sharma A,Kumar V,Singh R,et al. Effect of seed pre-soaking with 24-epibrassinolide on growth and photosynthetic parameters of Brassica juncea L. in imidacloprid soil[J]. Ecotoxicology and Environmental Safety,2016,133:195-201.
[54]Ergo V V,Veas R E,Vega C R C,et al. Leaf photosynthesis and senescence in heated and droughted field-grown soybean with contrasting seed protein concentration[J]. Plant Physiology and Biochemistry,2021,166:437-447.
[55]Zeng R E,Chen T T,Wang X Y,et al. Physiological and expressional regulation on photosynthesis,starch and sucrose metabolism response to waterlogging stress in peanut[J]. Frontiers in Plant Science,2021,12:601771.
[56]蔡 倩. 玉米和花生對水分胁迫的响应机制及灌水技术研究[D]. 沈阳:沈阳农业大学,2017:1-2.
[57]Giménez-Moolhuyzen M,van der Blom J,Lorenzo-Mínguez P,et al. Photosynthesis inhibiting effects of pesticides on sweet pepper leaves[J]. Insects,2020,11(2):69.
[58]Ren X W,Tang J C,Wang L,et al. Microplastics in soil-plant system:effects of nano/microplastics on plant photosynthesis,rhizosphere microbes and soil properties in soil with different residues[J]. Plant and Soil,2021,462(1):561-576.
[59]Teresa G S M,Carlos G,Julen U,et al. Mycorrhizal-assisted phytoremediation and intercropping strategies improved the health of contaminated soil in a peri-urban area[J]. Frontiers in Plant Science,2021,12:693044.
[60]Lv J X,Dong Y,Dong K,et al. Intercropping with wheat suppressed Fusarium wilt in faba bean and modulated the composition of root exudates[J]. Plant and Soil,2020,448(1):153-164.
[61]单明娟,秦 华,陈俊辉,等. 两种间作体系对丛枝菌根真菌侵染及多氯联苯去除的影响[J]. 应用与环境生物学报,2018,24(3):470-477.
[62]Jo S G,Kang Y I,Om K S,et al. Growth,photosynthesis and yield of soybean in ridge-furrow intercropping system of soybean and flax[J]. Field Crops Research,2022,275:108329.
[63]Zhou T,Wang L,Sun X,et al. Improved post-silking light interception increases yield and P-use efficiency of maize in maize/soybean relay strip intercropping[J]. Field Crops Research,2021,262:108054.
[64]Feng L Y,Ali Raza M,Shi J Y,et al. Delayed maize leaf senescence increases the land equivalent ratio of maize soybean relay intercropping system[J]. European Journal of Agronomy,2020,118:126092.
[65]朱文旭,张会慧,许 楠,等. 间作对桑树和谷子生长和光合日变化的影响[J]. 应用生态学报,2012,23(7):1817-1824.
[66]Chen X F,Sun N,Gu Y,et al. Photosynthetic and chlorophyll fluorescence responses in maize and soybean strip intercropping system[J]. International Journal of Agriculture and Biology,2020,24(4):799-811.
[67]Bekele B,Ademe D,Gemi Y,et al. Evaluation of intercropping legume covers with maize on soil moisture improvement in misrak azerinet berbere woreda,SNNPR,Ethiopia[J]. Water Conservation Science and Engineering,2021,6(3):145-151.
[68]Marschner P,Zheng B.Direction and magnitude of the change in water content between two periods influence soil respiration,microbial biomass and nutrient availability which can be modified by intermittent air-drying[J]. Soil Biology and Biochemistry,2022,166:108559.
[69]Dou X Y,Wang R S,Zhou X,et al. Soil water,nutrient distribution and use efficiencies under different water and fertilizer coupling in an apple-maize alley cropping system in the Loess Plateau,China[J]. Soil and Tillage Research,2022,218:105308.
[70]张丽芳,胡海林. 土壤酸碱性对植物生长影响的研究进展[J]. 贵州农业科学,2020,48(8):40-43.
[71]唐 琨,朱伟文,周文新,等. 土壤pH对植物生长发育影响的研究进展[J]. 作物研究,2013,27(2):207-212.
[72]Daraz U,Li Y,Sun Q Y,et al. Inoculation of Bacillus spp. modulate the soil bacterial communities and available nutrients in the rhizosphere of vetiver plant irrigated with acid mine drainage[J]. Chemosphere,2021,263:128345.
[73]王 晖,邢小军,许自成. 攀西烟区紫色土pH值与土壤养分的相关分析[J]. 中国土壤与肥料,2007(6):19-22,49.
[74]Hooper S L,Burstein H J.Minimization of extracellular space as a driving force in prokaryote association and the origin of eukaryotes[J]. Biology Direct,2014,9(1):1-42.
[75]秦昌鮮,彭 崇,郭 强,等. 甘蔗花生间作体系中红壤地根际pH值与有效磷含量的关系[J]. 广西糖业,2019(3):18-24.
[76]赵香兰. 有机质在果园土壤肥力中的作用[J]. 河北果树,1990(4):47-50.
[77]余涵霞,王家宜,万方浩,等. 植物凋落物影响土壤有机质分解的研究进展[J]. 生物安全学报,2018,27(2):88-94.
[78]邵 帅,何红波,张 威,等. 土壤有机质形成与来源研究进展[J]. 吉林师范大学学报(自然科学版),2017,38(1):126-130.
[79]寇建村,杨文权,李尚玮,等. 我国果园土壤有机质研究进展[J]. 北方园艺,2016(4):185-191.
[80]Zhang Y,Han M Z,Song M N,et al. Intercropping with aromatic plants increased the soil organic matter content and changed the microbial community in a pear orchard[J]. Frontiers in Microbiology,2021,12:616932.
[81]Tang X M,Zhang Y X,Jiang J,et al. Sugarcane/peanut intercropping system improves physicochemical properties by changing N and P cycling and organic matter turnover in root zone soil[J]. PeerJ,2021,9:e10880.
[82]Nyawade S O,Karanja N N,Gachene C K K,et al. Short-term dynamics of soil organic matter fractions and microbial activity in smallholder potato-legume intercropping systems[J]. Applied Soil Ecology,2019,142:123-135.
[83]Cong W F,Hoffland E,Li L,et al. Intercropping affects the rate of decomposition of soil organic matter and root litter[J]. Plant and Soil,2015,391(1/2):399-411.
[84]Agnan Y,Courault R,Alexis M A,et al. Distribution of trace and major elements in subarctic ecosystem soils:sources and influence of vegetation[J]. Science of the Total Environment,2019,682:650-662.
[85]Inselsbacher E,Wanek W.An unexpected source of nitrogen for root uptake:positively charged amino acids dominate soil diffusive nitrogen fluxes[J]. New Phytologist,2021,231(6):2104-2106.
[86]Costa Tania L,Sampaio Everardo V S B,Araújo Elcida L,et al. Contributions of Leguminosae to young and old stands of neotropical forests under different environmental conditions[J]. Annals of Forest Science,2021,78(2):48.
[87]Nyawade S O,Karanja N N,Gachene C K K,et al. Optimizing soil nitrogen balance in a potato cropping system through legume intercropping[J]. Nutrient Cycling in Agroecosystems,2020,117(1):43-59.
[88]Li L K,Zou Y,Wang Y H,et al. Effects of corn intercropping with soybean/peanut/millet on the biomass and yield of corn under fertilizer reduction[J]. Agriculture,2022,12(2):151.
[89]王 華,牛德奎,胡冬南,等. 不同肥料对油茶林土壤氮素含量、微生物群落及其功能的影响[J]. 植物营养与肥料学报,2014,20(6):1468-1476.
[90]Pierre W H,Norman A G. Soil and fertilizer phosphorus in crop nutrition[M]. New York:Academic Press,1953:1-16.
[91]Tang X Y,Zhang C C,Yu Y,et al. Intercropping legumes and cereals increases phosphorus use efficiency;a meta-analysis[J]. Plant and Soil,2020,460:89-104.
[92]Latati M,Blavet D,Alkama N,et al. The intercropping cowpea-maize improves soil phosphorus availability and maize yields in an alkaline soil[J]. Plant and Soil,2014,385(1/2):181-191.
[93]Tomohiro N,Yasuhiro T,Tovohery R,et al. Soil phosphorus retention can predict responses of phosphorus uptake and yield of rice plants to P fertilizer application in flooded weathered soils in the central Highlands of Madagascar[J]. Geoderma,2021,402:115326.
[94]Zhao X,Lyu Y,Jin K M,et al. Leaf phosphorus concentration regulates the development of cluster roots and exudation of carboxylates in Macadamia integrifolia[J]. Frontiers in Plant Science,2021,11:610591.
[95]Wen Z H,Pang J Y,Tueux G,et al. Contrasting patterns in biomass allocation,root morphology and mycorrhizal symbiosis for phosphorus acquisition among 20 chickpea genotypes with different amounts of rhizosheath carboxylates[J]. Functional Ecology,2020,34(7):1311-1324.
[96]孙宝茹.玉米/紫花苜蓿间作磷素高效吸收利用的根系-土壤互作机理[D]. 长春:东北师范大学,2017:2-12.
[97]俞巧钢,叶 静,杨梢娜,等. 不同施氮量对单季稻养分吸收及氨挥发损失的影响[J]. 中国水稻科学,2012,26(4):487-494.
[98]王立梅,刘奕清,阮玉娟. 植物钾素研究进展[J]. 中国园艺文摘,2015,31(5):71,148.
[99]Oosterhuis D M,Loka D A,Kawakami E M,et al. The physiology of potassium in crop production[J]. Advances in Agronomy,2014,126:203-233.
[100] Zhu L Z,He J,Tian Y,et al. Intercropping Wolfberry with Gramineae plants improves productivity and soil quality[J]. Scientia Horticulturae,2022,292:110632.
[101]Wu T,Qin Y,Li M.Intercropping of tea (Camellia sinensis L.) and Chinese chestnut:variation in the structure of rhizosphere bacterial communities[J]. Journal of Soil Science and Plant Nutrition,2021,21(3):2178-2190.
[102]Fan Y F,Wang Z L,Liao D P,et al. Uptake and utilization of nitrogen,phosphorus and potassium as related to yield advantage in maize-soybean intercropping under different row configurations[J]. Scientific Reports,2020,10(1):9504.
[103]Shen X F,Zhao Z H,Chen Y.Effects of intercropping with peanut and silicon application on sugarcane growth,yield and quality[J]. Sugar Tech,2019,21(3):437-443.
[104]姜 勇,徐柱文,王汝振,等. 長期施肥和增水对半干旱草地土壤性质和植物性状的影响[J]. 应用生态学报,2019,30(7):2470-2480.
[105]李 娟,林位夫,周立军,等. 成龄胶园间作不同豆科作物对土壤养分的影响[J]. 热带农业科学,2014,34(7):5-11.
[106]董晓伟. 银杏复合经营土壤微量元素特征及有效性影响因素研究[D]. 南京:南京林业大学,2013:3-8.
[107]张 蓓,刘林婷,刘若南,等. 钙与IAA对易裂果蜜广橘果皮活性氧代谢和相关抗氧化基因表达的影响[J]. 果树学报,2021,38(12):2034-2044.
[108]Chen Z C,Peng W T,Li J,et al. Functional dissection and transport mechanism of magnesium in plants[J]. Seminars in Cell & Developmental Biology,2018,74:142-152.
[109]Chernev P,Fischer S,Hoffmann J,et al. Light-driven formation of manganese oxide by todays photosystem Ⅱ supports evolutionarily ancient manganese-oxidizing photosynthesis[J]. Nature Communications,2020,11:6110.
[110]夏海勇,薛艳芳,孟维伟,等. 间套作体系作物-土壤铁和锌营养研究进展[J]. 应用生态学报,2015,26(4):1263-1270.
[111]李泓池,张 洁,蔡传涛. 微量元素铜锰配施对密花豆生长、光合和生物量的影响[J]. 热带农业科学,2022,42 (6):17-22.
[112]李峤虹. 缺硼胁迫对枳根系生长发育及相关基因表达的影响研究[D]. 武汉:华中农业大学,2016:2-12.
[113]Vinichuk M,Bergman R,Sundell-Bergman S,et al. Response of spring wheat and potato to foliar application of Zn,Mn and EDTA fertilizers on 137Cs uptake[J]. Journal of Environmental Radioactivity,2021,227:106466.
[114]李 娟,周立军. 间作五指毛桃土壤和根主要中微量元素含量及其相关性[J]. 土壤,2020,52(3):645-649.
[115]Cao X R,Wang X Z,Lu M,et al. The Cd phytoextraction potential of hyperaccumulator Sedum alfredii-oilseed rape intercropping system under different soil types and comprehensive benefits evaluation under field conditions[J]. Environmental Pollution,2021,285:117504.
[116]Rasmussen C R,Thorup-Kristensen K,Dresbll D B. The effect of drought and intercropping on chicory nutrient uptake from below 2 m studied in a multiple tracer setup[J]. Plant and Soil,2020,446(1):543-561.
[117]李冬梅. 小麦/苜蓿间作的土壤微生物多样性和种间促进作用研究[D]. 哈尔滨:东北林业大学,2015:11-12.
[118]陈新新. 果园间作芳香植物对土壤微生物多态性及碳氮循环的影响[D]. 北京:北京农学院,2014:1-5.
[119]Tang L,Hamid Y,Zehra A,et al. Fava bean intercropping with Sedum alfredii inoculated with endophytes enhances phytoremediation of cadmium and lead co-contaminated field[J]. Environmental Pollution,2020,265(Part A):114861.
[120]Trap J,Riah W,Akpa-Vinceslas M,et al. Improved effectiveness and efficiency in measuring soil enzymes as universal soil quality indicators using microplate fluorimetry[J]. Soil Biology and Biochemistry,2012,45:98-101.
[121]Muscolo A,Settineri G,Attinà E.Early warning indicators of changes in soil ecosystem functioning[J]. Ecological Indicators,2015,48:542-549.
[122]Caldwell B A. Enzyme activities as a component of soil biodiversity:a review[J]. Pedobiologia,2005,49(6):637-644.
[123]Kumar A,Blagodaskaya E,Dippold M A,et al. Positive intercropping effects on biomass production are species-specific and involve rhizosphere enzyme activities:evidence from a field study[J]. Soil Ecology Letters,2021,4(4):444-453.
[124]王吉秀,祖艷群,李 元,等. 玉米和不同蔬菜间套模式对重金属Pb、Cu、Cd累积的影响研究[J]. 农业环境科学学报,2011,30(11):2168-2173.
[125]刘均霞,陆引罡,远红伟,等. 玉米、大豆间作对根际土壤微生物数量和酶活性的影响[J]. 贵州农业科学,2007,35(2):60-61,64.
[126]孟自力,叶美金,闫延梅,等. 间作大蒜对小麦根际土壤微生物数量及土壤酶活性的影响[J]. 农业资源与环境学报,2018,35(5):430-438.
[127]Yan Z Q,Li Y,Wu H D,et al. Different responses of soil hydrolases and oxidases to extreme drought in an alpine peatland on the Qinghai-Tibet Plateau,China[J]. European Journal of Soil Biology,2020,99:103195.
[128]Wang C L,Shi B K,Sun W,et al. Different forms and rates of nitrogen addition show variable effects on the soil hydrolytic enzyme activities in a meadow steppe[J]. Soil Research,2020,58(3):258.
[129]Yang J S,Yang F L,Yang Y,et al. A proposal of “core enzyme” bioindicator in long-term Pb-Zn ore pollution areas based on topsoil property analysis[J]. Environmental Pollution,2016,213:760-769.
[130]Amini Kiasari M,Pakbaz M S,Ghezelbash G R.Increasing of soil urease activity by stimulation of indigenous bacteria and investigation of their role on shear strength[J]. Geomicrobiology Journal,2018,35(10):821-828.
[131]Farooq T H,Kumar U,Mo J,et al. Intercropping of peanut–tea enhances soil enzymatic activity and soil nutrient status at different soil profiles in subtropical southern China[J]. Plants,2021,10(5):881.
[132]Ma Y H,Fu S L,Zhang X P,et al. Intercropping improves soil nutrient availability,soil enzyme activity and tea quantity and quality[J]. Applied Soil Ecology,2017,119:171-178.
[133]Watanabe K,Hayano K. Source of soil protease in paddy fields[J]. Canadian Journal of Microbiology,1993,39(11):1035-1040.
[134]張 威,张 明,张旭东,等. 土壤蛋白酶和芳香氨基酶的研究进展[J]. 土壤通报,2008,39(6):1468-1474.
[135]王玉新,王天竺,高 平,等. 薰衣草间作两种十字花科蔬菜对土壤酶活性的影响[J]. 中国林副特产,2020(6):25-27.
[136] 颜彩缤,胡福初,王彩霞,等. 槟榔-平托花生间作对土壤养分和土壤酶活性的影响[J]. 热带农业科学, 2020,40(11):14-22.
[137]Gu C H,Wilson S G,Margenot A J. Lithological and bioclimatic impacts on soil phosphatase activities in California temperate forests[J]. Soil Biology and Biochemistry,2020,141:107633.
[138]Leite M V M,Bobulská L,Espíndola S P,et al. Modeling of soil phosphatase activity in land use ecosystems and topsoil layers in the Brazilian Cerrado[J]. Ecological Modelling,2018,385:182-188.
[139]Presti E L,Badagliacca G,Romeo M,et al. Does legume root exudation facilitate itself P uptake in intercropped wheat?[J]. Journal of Soil Science and Plant Nutrition,2021,21(4):3269-3283.
[140]Qin X M,Pan H N,Xiao J X,et al. Increased nodular P level induced by intercropping stimulated nodulation in soybean under phosphorus deficiency[J]. Scientific Reports,2022,12:1991.
[141]de Souza Soares A,Augusto P E D,de Castro Leite B R Jr,et al. Ultrasound assisted enzymatic hydrolysis of sucrose catalyzed by invertase:investigation on substrate,enzyme and kinetics parameters[J]. LWT,2019,107:164-170.
[142]Gao J X,Xie H. Daylily intercropping:effects on soil nutrients,enzyme activities,and microbial community structure[J]. Frontiers in Plant Science,2023,14:1107690.
[143]陈 伟,孙从建,李卫红. 低氮胁迫下苦荞根际土壤纤维素酶活性的响应机制:荧光光谱法测定[J]. 光谱学与光谱分析,2018,38(10):3159-3162.
[144]韩春梅,李春龙,叶少平,等. 不同栽培模式对生姜大田土壤酶活性及土壤养分的动态影响[J]. 北方园艺,2016(4):163-167.
[145]黄田田. 饲用油菜高丹草间作结合施肥对饲草产质量及土壤有机碳组分的影响[D]. 呼和浩特:内蒙古农业大学,2020:31-32.
[146]袁婷婷,赵 骞,董 艳. 阿魏酸胁迫下间作对蚕豆枯萎病发生和根系组织结构的影响[J]. 土壤学报,2021,58(4):1060-1071.
[147] 常学秀,文传浩,沈其荣,等. 锌厂Pb污染农田小麦根际与非根际土壤酶活性特征研究[J]. 生态学杂志,2001,20(4):5-8.
[148]郑晓媛. 桑树/大豆间作的种间关系及其土壤磷酸酶变化[D]. 哈尔滨:东北林业大学,2011:41.
[149]张智晖. 玉米/大豆间作模式对土壤酶活性及土壤养分的影响[J]. 安徽农业科学,2011,39(16):9706-9707.
[150]于 路,黄宇婷,张 奎,等. 间作紫花苜宿促进苹果园土壤氮循环的机制[J]. 北京农学院学报,2023,38 (3):34-39.
[151]代真林,汪婭婷,姚秀英,等. 玉米大豆间作模式对玉米根际土壤微生物群落特征、玉米产量及病害的影响[J]. 云南农业大学学报(自然科学),2020,35(5):756-764.
[152]Cheng G L,Liu H,Wang Q H.Effects of EDTA,EDDS and oxalic acid on catalase and peroxydase activities of soil[J]. Advanced Materials Research,2011,322:39-42.
[153]Wu C F,Liu J,Xia D,et al. Effects of grape seedlings intercropping with post-grafting generation of two floricultural accumulator plants on soil enzymes activity under cadmium stress[J]. IOP Conference Series:Earth and Environmental Science,2019,233:042017.
[154]Zhou X G,Yu G B,Wu F Z.Effects of intercropping cucumber with onion or garlic on soil enzyme activities,microbial communities and cucumber yield[J]. European Journal of Soil Biology,2011,47(5):279-287.
[155]崔爱花,孙亮庆,刘 帅,等. 棉花产量和土壤微生物数量及酶活性对棉田间作系统的响应[J]. 江苏农业科学,2022,50(2):53-58.
[156]覃潇敏,黄少欣,韦锦坚,等. 茶树/大豆间作对茶树土壤和茶叶营养品质的影响[J]. 华北农学报,2019,34(增刊1):129-135.
[157]韦持章,农玉琴,陈远权,等. 茶树/大豆间作对根际土壤微生物群落及酶活性的影响[J]. 西北农业学报,2018,27(4):537-544.
[158]文亚雄. 林农间作油茶林土壤微生物多样性研究[D]. 长沙:中南林业科技大学,2015:36-37.
[159]朱国政. AMF对金橘大豆间作体系中金橘生长及根际土微生物多样性的影响[D]. 桂林:广西师范大学,2015:33.
[160]范 虹,赵 财,胡发龙,等. 土壤含水量、种植模式和丁香酚对土壤微生物种群结构的影响[J]. 中国沙漠,2019,39(2):13-18.
[161]李梦辉,张瑞花,朱 军,等. 不同间隔方式作用下香蕉、辣椒的间作效应[J]. 分子植物育种,2017,15 (3):1156-1163.
[162]张海春,张 浩,胡晓辉. 不同间作模式对温室连作番茄产量、土壤微生物和酶的影响[J]. 西北农业学报,2016,25(8):1218-1223.
[163]吕慧芳,别之龙. 间作小麦对西瓜生长及根际土壤酶和微生物的影响[J]. 中国瓜菜,2019,32(6):32-37.
[164]张亮亮,罗 明,徐金虹,等. 南疆枣树棉花间作对土壤微生物生物量碳、氮的影响[J]. 新疆农业大学学报,2015,38(3):216-223.
[165]王庆宇,李立军,阮 慧,等. 旱地燕麦间作对土壤酶活性、微生物含量及产量的影响[J]. 干旱地区农业研究,2019,37(2):179-184.
[166] 马 琨,杨桂丽,马 玲,等. 间作栽培对连作马铃薯根际土壤微生物群落的影响[J]. 生态学报,2016,36(10):2987-2995.
[167]牛 宇,薄晓峰,秦作霞,等. 间作豆类作物对大樱桃生长和土壤特性、微生物数量的影响[J]. 北方园艺,2019(4):38-44.
[168]杜春凤. 马铃薯/玉米间作栽培对土壤和作物的影响[D]. 银川:宁夏大学,2017:23.
[169]张萌萌,敖 红,李 鑫,等. 桑树/苜蓿间作对根际土壤酶活性和微生物群落多样性的影响[J]. 草地学报,2015,23(2):302-309.
[170]逄好胜,胡举伟,李 鑫,等. 小麦-苜蓿间作对土壤微生物群落功能多样性的影响[J]. 贵州农业科学,2015,43(8):160-164.
[171]白鹏华,刘奇志,张林林,等. 南疆土壤线虫及微生物对枣树与绿豆间作种植方式的响应[J]. 西北农业学报,2015,24(2):104-110.
[172]夏 枫. 施肥与间作对设施蔬菜大棚土壤养分及微生物变化影响的研究[D]. 海口:海南大学,2017:44-45.
[173]董宇飞,吕相漳,张自坤,等. 不同栽培模式对辣椒根际连作土壤微生物区系和酶活性的影响[J]. 浙江农业学报,2019,31(9):1485-1492.
[174]谭礼宁. 谷子/花生盆栽间作蛋白质组学及根际微生物区系研究[D]. 福州:福建农林大学,2015:78-79.
[175]邓 文,胡兴明,于 翠,等. 桑树间作大豆对桑园土壤微生物多样性的影响[J]. 蚕业科学,2015,41(6):997-1003.
[176]乔 旭. 丛枝菌根真菌在植物种间互作中的调节机 制[D]. 北京:中国农业大学,2016:82.
[177]王光州. 土壤微生物调节植物种间互作和多样性—生产力关系的机制[D]. 北京:中国农业大学,2018:88-93.
收 稿日期:2023-04-06
基金项目:崇左市科技计划(编号:崇科FA2018005)。
作者简介:欧阳子龙(1997—),男,湖北天门人,硕士,主要从事园林植物资源及应用研究。E-mail:1627132117@qq.com。
通信作者:滕维超,博士,副教授,主要从事园林植物资源及应用研究。E-mail:119754446@qq.com。
猜你喜欢
中学生理科应试(2024年6期)2024-01-01 00:00:00
河北果树(2020年4期)2020-11-26 06:05:18
江苏农业科学(2016年12期)2017-04-05 17:58:37
江苏农业科学(2016年8期)2017-02-15 19:00:23
江苏农业科学(2016年8期)2017-02-15 19:00:20
新疆农垦科技(2016年2期)2016-08-21 13:50:18
江苏农业科学(2016年6期)2016-07-25 23:51:36
江苏农业科学(2016年6期)2016-07-25 00:07:00
江苏农业科学(2016年2期)2016-04-11 07:17:01
湖北农业科学(2015年15期)2015-09-09 21:56:48
