
模拟干旱下杨树树干液流特征及其对环境因子的响应
2024-03-18 07:57张星宇杨金艳阮宏华张瑞婷
水土保持研究 2024年2期
张星宇,杨金艳,阮宏华,张瑞婷
(南京林业大学生物与环境学院,南京 210037)
IPCC评估报告指出,大气中的CO2和CH4浓度的增加居高不下,导致全球变暖,降水格局发生变化,这会增加局部地区干旱事件发生频率,尤其在中纬度地区[1]。中纬度地区干旱事件频发显著影响了陆地生态系统的碳、水循环,甚至可能使得陆地生态系统从碳源转变为碳汇[2-3]。
陆地向大气输送水分的唯一途径是森林蒸发散[4],森林生态系统是陆地生态系统的主体,因此森林蒸发散是整个生态系统耗水量的重要组分。植物树干液流可以准确测量单木蒸腾耗水量,是估测林分耗水的关键指标之一[5]。另外其与一系列环境因子紧密联系,气象因子可以直接影响植物的瞬时液流速率,而土壤水分直接决定了植物的可利用水量[6]。已有学者针对干旱下植物树干液流对环境因子的响应作了一系列研究,如刘延惠等在茂兰喀斯特原生林细叶青冈树干液流对环境因子的试验中,发现影响树干液流的主要环境因子是光照、空气相对湿度和饱和水汽压差,与土壤含水量无显著相关[7]。王艳兵等[8]研究则认为土壤水分对干旱区或干旱立地条件下的树干液流格外重要。通过对干旱区植物茎干液流变化规律的研究,可以有效地反映出植物对干旱环境的适应机制[9]。目前,关于植物液流速率差异特征已有较多研究,但大多集中于液流速率的个体差异及其在边材上的径向差异,对植株不同干旱下的液流差异研究较少。
我国人工林面积8 003万hm2,其中,杨树人工林面积达到853万hm2,居世界第一,江苏省作为杨树人工林重要栽培区域,其干旱的频度和强度近年也有增加[10-11],尤其是在苏北区域。干旱频发造成杨树人工林水分利用率和光合生产效率均下降,而后其生产力降低[12]。本文以江苏省盐城东台国有林场10 a生杨树(Populusdeltoides)人工林为对象,运用干旱模拟试验,设置空白对照组(CK)、穿透雨移除30%(D1)和穿透雨移除50%(D2)试验组,结合同步监测的环境因子(2018年9月—2020年11月),揭示杨树人工林树干液流对干旱的响应规律与机制。旨在科学评估降水格局改变对森林生态系统碳收支影响提供新的理论认知,也为进一步精确预测降水格局改变对人工林生态系统的影响提供理论基础。
1 研究区概况与研究方法
1.1 研究区概况
试验地位于江苏省盐城市东台市境内的东台国有林场(120°49′E,32°52′N)。东台林场是江苏省沿海重点防护林,临近黄海森林公园,地处黄海之滨,气候类型为过渡地带海洋性季风气候。年平均气温为14.6℃,年平均相对湿度为88.3%,无霜期为225 d,年均降雨量为1 050 mm,年平均日照时间为2 200 h,10℃以上年积温为4 565℃。土壤为脱盐草甸土,土壤质地为砂质壤土,土壤p H 值为偏碱性。区域内主要林分类型为杨树(Populus)、银杏(Ginkgobiloba)、杉木(Cunninghamialanceolata)等。选取10 a生杨树人工林(黑杨派无性系I-35,PopulusdeltoidesCL′35′)为研究对象,平均树高为21 m,平均胸径为23 cm,林分郁闭度为0.60,间距为4 m×6 m。
1.2 研究方法
1.2.1 样地设置 采取随机区组设计(RCBD)进行干旱处理的野外控制试验。干旱处理包括3个水平:对照(CK)、穿透雨移除30%(D1)和穿透雨移除50%(D2)。杨树人工林依据样地内生境条件和前期本底调查分为3个区组(区组间距在300~500 m),每种处理设定3个重复样地,共选定9块观测样地(不同样地随机交叉布置),每个样地面积为25 m×25 m,其中15 m×15 m 为核心测定区域,余下为缓冲区域。每个样地间距为20 m。
1.2.2 杨树树干液流测定与计算 在2018年9月—2020年11月,在杨树1.20 m处安装TDP插针式植物茎流计(Probe12:美国,Dynamax公司),经过48 h以上的缓冲后,记录树干液流数值。树干液流全年连续测定,每0.5 h记录一次数据。液流通过在选定的样树1.20 m处安装TDP 插针式植物茎流计(Probe12,Dynamax,USA)测定相关温度数据,公式计算[13]:
式中:Js为液流密度〔g/(m2·s)〕;dTm为24 h内上下探针的最大温差值(℃);dT为某时刻瞬时温差值(即实时测定的温差值)(℃)。
1.2.3 环境因子测定与计算 根据样地生境差异情况,所有样地划分为3个区组,在2018年9月—2020年11月每个区组利用一套EM50微型气象监测系统(Decagon,USA)对环境因素进行全年连续监测。环境因素指标有空气温度(Ta,℃)、大气相对湿度(Rh,%)、降雨量(P,mm)、光合有效辐射〔Qo,μmol/(m2·s)〕、土壤15 cm 深度的土壤温度(Ts,℃)、土壤体积含水量(VSM,%)、土壤水势(SWP,k Pa)。利用EM50数据采集器,每0.5 h记录一次数据。饱和水汽压差(VPD,k Pa)公式计算[14]:
式中:Ta为大气温度(℃);Rh为相对湿度(%)。
1.3 数据统计分析
液流数据、环境因子进行自动化记录,利用Excel进行数据整理及制图,利用Origin软件绘制液流及环境因子时间动态图;利用R 语言构建混合效应模型,研究期内,为进行多个解释变量的相对重要性分析,本文使用R 语言中的Mu MIn包进行模型的选择,分析哪些变量是解释干旱条件下杨树树干液流变化的重要生态因子。Js作为响应变量,VPD,Ta,P,Qo,Ts,SWP作为解释变量,拟合一个多元非线性回归。首先,使用全部的解释变量,构建一个其与响应变量(Js)的全模型;使用Mu MIn包进行模型选择,获得最优子集,寻求简约模型(利用对变量间关系的解读)的同时最大程度地保留模型解释能力。接着使用rdacca.hp包分析解释变量的相对重要性。
2 结果与分析
2.1 液流与环境因子的动态变化
在2019—2020 两年试验期内,部分环境因子(Ta,Qo和Ts)变化曲线具有相似的动态格局(图1)。温度范围年际差异小,为-7.5~33.53℃。Ts与Ta基本同步,且Ts变化较为稳定,为3~27.4℃,变化幅度比Ta小44%;Qo在两年时间内变化曲线具有相似性,全年变化大致呈双峰型,在1月—2月Qo最低,为5 664 μmol/(m2·s),之后逐渐增加,在6月抵达第一个峰值,为1 036μmol/(m2·s),之后逐渐减缓,8月降至780 μmol/(m2·s),之后稍回升至第2个峰值后再次下降。研究区有明显的雨季旱季区别,雨季时间为7—9月3个月份,两年研究期内雨季P均占全年P一半以上,2019年和2020年分别占54%和65%。VPD 最大值出现在5月24日(2.77 k Pa),前后连续5 d VPD>1.9 kPa,两年均值为0.50 k Pa(0.2~1.06 kPa)。

图1 液流研究期内环境因子的变化Fig.1 Changes of environmental factors during the study period of sap flow(A)Ta and P;(B)Qo;(C)VPD;(D)Ts
在不同干旱处理下对Js和VSM 的影响(图2)。2019年VSM 在D2组较CK 组和D1组均显著降低(p<0.05),但D1组VSM 并未显著下降,在2020年,经过一年的干旱处理后,D1组和D2组的VSM均较CK 组显著下降。Js在两年监测期内D1组和D2组较CK 组均显著下降。

图2 不同干旱处理下对液流密度和土壤体积含水量的影响Fig.2 Effects of Different Drought Treatments on sap flow density and soil volumetric water content
杨树生长季的时间为4月中下旬至10月中旬,早于雨季(图3)。Js年变化呈单峰型曲线,与VPD,Qo的年际变化曲线相似。在3种干旱处理下,2019年和2020年Js均值排名始终为CK〔(32.06 g/(m2·s),25 g/(m2·s)〕>D1〔22.72 g/(m2·s),18.15 g/(m2·s)〕>D2〔17.97 g/(m2·s),15.78 g/(m2·s)〕;Js年际差异变化大,主要集中在生长季,Js最大日均值〔104.26 g/(m2·s)〕出现在2019年的CK样地;CK,D1和D2三组在2019年生长季均值〔61.06,47.25,36.25 g/(m2·s)〕均高于2020 年生长季均值〔42.49,31.98,27.46 g/(m2·s)〕。但在两年试验期内,全年均值差别不大〔±4 g/(m2·s)〕。VSM 在生长季前会到达年峰值,生长季后大幅下降;生长季内的雨季来临时,VSM 缓慢回升,但在年尺度上,仍是一年总最低的水平(28%);雨季即将结束时,VSM 会有较明显的回升;生长季结束后,VSM 先降后升。在CK,D1和D2三组中,随着截流程度增加,VSM 降幅随之增大,差异最大时空白组的含水量是移除D1 组的1.24 倍,是D2组的1.5倍;两年平均值也存在明显差异。

图3 模拟干旱试验中土壤体积含水量及液流的年变化Fig.3 Annual changes of soil volumetric water content and sap flow in simulated drought test
在两年生长季(4月下旬—10月中旬)期间的典型连续晴天内,Js的峰值时间均为10:00—12:00(图4),基本17:00开始Js急速下降,夜间Js均值为0.78 g/(m2·s)。非生长季的液流峰明显较窄,且出现时间晚于生长季。但在夜间仍然存在微弱液流〔0.24 g/(m2·s)〕,占液流峰值的10%,同年生长季的液流峰值几乎是非生长季的30倍。
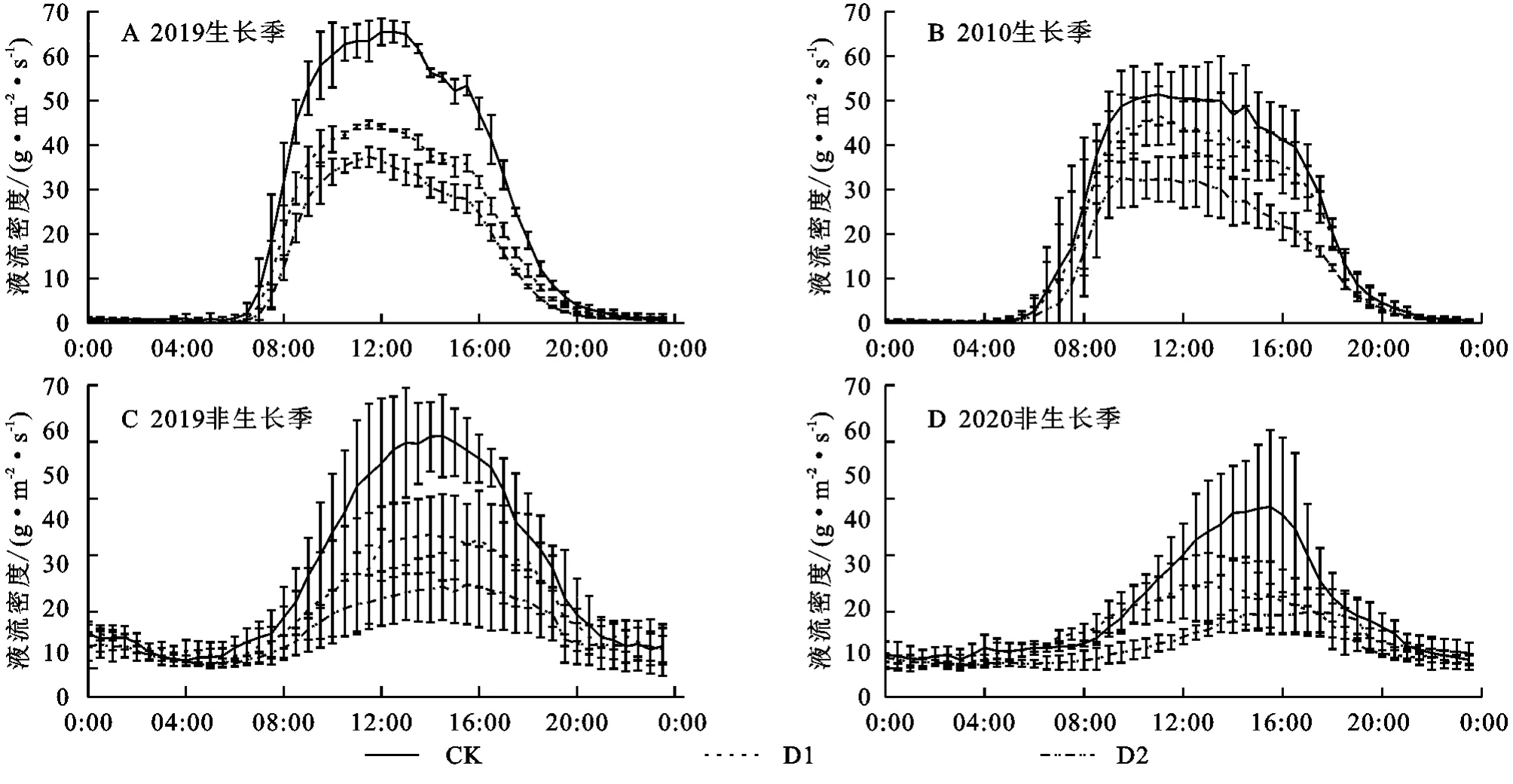
图4 典型晴天下液流生长季与非生长季的日变化Fig.4 Diurnal changes of sap flow in growing season and non-growing season under a typical sunny day
2.2 模拟干旱下杨树树干液流与环境因子的关系
将各环境因子与杨树人工林树干液流在全模型的基础上进行模型选择得以确定影响树干液流的最佳解释变量(图5)。3个样地环境因子揭示树干液流变化均在0.6以上,因此能较好地预测不同干旱条件下液流对环境因子的响应。因为各环境因子能够彼此影响并不是独立的,如降雨改变时,土壤含水量会随之改变,本文更准确地探讨了各环境因子的方差解释率。从图5中可以看出,在CK 样地,Qo,SWP,Ts,VPD 与树干液流极显著相关,能解释液流变化的65.8%;其中Ts(61.53%)对液流变化的贡献最大,其次是VPD(19.30%)。在D1样地中,VPD,Ts,SWP能解释液流变化的68.7%,仍是Ts(68.44%)对液流变化的影响贡献最大,SWP与液流变化呈负相关,VPD,Ts与液流变化呈正相关。在D2样地中,VPD,Ts,Ta,SWP,P能解释此样地液流变化的64.1%,D1样地杨树人工林比其他两组,对环境因子,尤其是Ta更敏感(R2=0.36)。混合效应模型结果表明:VPD和Ts与液流变化极显著相关(p<0.01),Ta,P和SWP与液流变化显著相关(p<0.05),此模型能解释液流变化的61.9%。
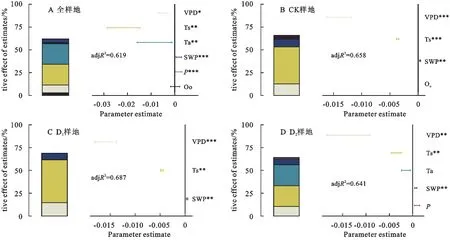
图5 模拟干旱下液流对环境因子的下响应Fig.5 Simulated response of sap flow to environmental factors under drought conditions
3 讨论
3.1 液流与环境因子的动态变化
研究区内树干液流速率在夏季达到峰值,接着随着时间推移逐渐下降,到达林木休眠期时液流密度趋近于0。在生长季内,为维持生长发育,液流活动强烈,而在非生长季内,液流活动受到树木生长状况的影响而减弱[15]。树木液流受环境影响具有较大复杂性。这与Qo的年际变化相似,但Qo的R2(0.47)较小,我们认为是未将生长季与非生长季区分的原因。
根据混合效应模型(图5),在环境因子中,Ts(36.85%)和Ta(36.40%)对液流变化的贡献最大,但Ta每日昼夜温差波动较大,Ts则较为稳定。其次为VPD(13.72%)。这与诸多学者研究一致。Ts升高能促进树干液流的启动,并控制土壤水分要素进而改变林地蒸散。苏军德等使用TDP技术测定祁连山圆柏(Sabinachinensis)的液流变化,发现Ta是主要限制因子[16];Hayat等通过多元回归模型分析Ta,VPD 和气孔导度(Gs)共同解释了半干旱区沙柳(Salixpsammophila)Js中中63%的变化量[2]。另外进一步说明了VPD 与树干液流密切相关,VPD 能改变植物表面与大气的水汽压差,进而影响空气阻力和气孔导度等导致植物蒸腾速率的差异,当饱和水汽压差处于较小的范围时,随气孔导度增大,气孔对饱和水汽压差的敏感度逐渐增加[17]。温度、Qo和VPD是影响液流变化的主要气象因素;土壤含水量是相关性较强的土壤因素,结果与前人研究一致。
除影响树干液流的众多环境因子外,树木自身的生物学结构特征,如木质部导水率径向生长,也会影响到树干液流[18]。这需要展开进一步研究进行说明。本研究杨树人工林树龄、胸径均基本一致,有必要在模拟干旱下对不同树龄、多个树种进行进一步试验研究。此外,本文还发现土壤温度对液流变化起到较为直观的影响,且土壤温度变异程度较小,便于测算,可以考虑增加这部分研究,以了解液流对地下环境因子的响应机制。
3.2 模拟干旱对液流的影响
在本模拟干旱试验中,随着截流程度增大,干旱样地的土壤含水量和树干液流均低于对照样地(图3),降雨通过影响土壤含水量,对树干液流变化的影响增大(图5),Besson在葡萄牙南部的地中海林地上通过分别采用干旱、环境和湿润处理试验得出截流降水使林分冠层年蒸腾作用降低了10%,而灌溉使其增加了11%,这与本试验研究结论相似[19]。
许多学者对不同干旱梯度植物树干液流的影响开展了诸多研究,大量研究表明植物的树干液流随着截流程度的增大而降低,进而影响林地蒸散。如孛永明在黄土高原利用包裹式液流计研究柠条和沙棘树干液流与环境因子的关系,结果表明土壤含水量对液流变化的影响较大,降雨补给了土壤水分,对树干液流密度存在脉冲效应[20]。Brinkmann等研究温带树种对夏季土壤干燥的液流响应研究中认为土壤湿度下降可以通过降低树干液流从而引发树木的强烈生理反应[21]。韩新生等研究认为山杏树干液流与土壤水分呈极显著负相关[22]。本研究结论为在生长季前期杨树人工林树干液流与土壤含水量呈正相关,后期转为负相关(图4),即树干液流受到土壤温度和土壤水势的显著影响。我们认为树种生物学特性及根系适应土壤水分机制的差异,以及研究时间尺度的差异。
在我们的试验中杨树人工林的林龄和胸径基本一致,截流降水树干液流与土壤水分呈极显著正相关,有诸多学者与本试验研究结论相同[23-25]。进一步说明了雨季引起的表层土壤含水量变化造成的,移除30%的穿透雨提高了各气象因子对液流变化的解释程度,我们认为在一定的干旱条件下会提高树干液流对环境变化的敏感性,对此有利于完整地把握树木的水分利用策略。
4 结论
综上,树干液流年变化呈单峰型曲线,干旱处理下,随着截流程度增大,树干液流随着降低,且在D1样地对环境因子的敏感性提高。通过混合效应模型得出,土壤含水量对液流变化的解释率最高,气象因子中为饱和蒸汽压差,饱和蒸汽压差通过改变气孔导度进而改变树干液流,土壤含水量直接为植物根系供水从而影响植物树干液流,在一定的干旱条件下会提高树干液流对环境变化的敏感性,对此有利于完整地把握树木的水分利用策略。但在试验中杨树人工林的林龄和胸径差异不显著,鉴于野外试验条件限制以及森林生态系统水分循环影响因子的复杂性,本文仅对液流进行了浅层的研究和探索,仍需要继续探讨树木自身性状对环境的响应等,有助于提高对森林尺度水分收支的理解。
猜你喜欢
时代英语·高一(2019年1期)2019-03-13
现代园艺(2017年19期)2018-01-19
现代园艺(2017年21期)2018-01-03
喜剧世界(2017年10期)2017-06-01
电源技术(2017年1期)2017-03-20
山东林业科技(2016年5期)2016-07-05
广西林业科学(2016年3期)2016-03-16
电源技术(2015年7期)2015-08-22
中国质量与标准导报(2014年5期)2014-02-28
储能科学与技术(2014年4期)2014-02-27
