
内蒙古绒山羊KAP6.2基因多态性与生长性状和绒细度性状的关联分析
2024-03-12 08:44:40王聪亮张政轩白晶晶蓝贤勇陈生会朱海鲸
家畜生态学报 2024年3期
王聪亮,张政轩, 2△,白晶晶,付 琪,蓝贤勇,陈生会,屈 雷*,朱海鲸, 5, 6*
(1.榆林学院陕西省绒山羊工程技术研究中心,陕西 榆林 719000;2. 榆林市榆阳区牛家梁区域畜牧兽医站,陕西 榆林 719000; 3. 西北农林科技大学动物科技学院,陕西 杨凌 712000;4. 陕西神奥农业生物技术开发股份有限公司,陕西 神木 719318; 5. 陕西省“四主体一联合”肉羊工程技术校企联合研究中心,陕西 神木 719318;6. 陕西浩丽绒山羊科技发展有限公司,陕西 子洲 718499)
内蒙古绒山羊是内蒙古自治区经过长期自然选择和人工选育而培育的地方优质品种[1],独特的区域环境和气候使得内蒙古绒山羊肉质紧实、味美质优,羊绒纤维细而柔软,在国内外享有盛名[2-3]。作为绒肉兼用型品种,产肉性能往往与生长性状关系紧密,但目前很难做到肉、绒兼顾选育。家畜的体型外貌往往不同程度影响着生产性能[4],且生长性状越好,体重就越大,经济价值就越高,因而进行内蒙古绒山羊生长性状表型的选育,对于提高经济效益和育种改良意义重大。传统育种方法耗时长、难度大,而分子标记辅助选择(marker assisted selection, MAS)方法能够极大程度缩短育种年限、有效提高育种效率、快速培育高产优质品种和提高经济性状,在家畜育种过程中发挥较大作用[5-7],故利用MAS技术寻找影响山羊体尺和与绒相关等经济性状的重要候选基因和相关分子标记,对于加快该品种的育种进程有着重要理论价值。
角蛋白相关蛋白(Keratin- associated Protein, KAPs)和角蛋白中间丝(Keratin intermediate filament, KET-IF)是构成羊毛纤维的主要结构蛋白,按照其结构功能分为高硫蛋白(high-sulfer KAPs)、超高硫蛋白(Ultra-high-sulfer KAPs)和高甘氨酸酪氨酸蛋白(high-glycine-tyrosine KAPs),其中高甘氨酸酪氨酸蛋白包括KAP6.n、KAP7和KAP8多基因家族[8-11]。KAP在不同物种或不同品种毛发内的结构和含量差异较大,其中绵羊毛中KAP最高含量为12%,在针鼹鼠高达30%~40%[12-13]。研究人员通过遗传连锁分析和体细胞杂交技术将高甘氨酸酪氨酸蛋白KAP6.1、KAP8和高硫蛋白KAP1.1、KAP1.2以及KAP3.2分别定位在绵羊第1号和21号染色体上[14-15],其中KRTAP6和KRTAP8基因与美利奴羊纤维直径和羊毛产量显著相关[16]。张亚妮等[17]报道称,在绒山羊中,KAP1.1和KAP1.3基因分别与产绒量和体重显著相关,KAP13.1基因可作为羊绒纤维直径性状标记辅助选择的遗传标记[18],其中角蛋白基因的差异表达可导致不同地域环境和品种的绒山羊羊绒品质存在较大差异[19-20]。
目前关于KAP6.2基因与内蒙古绒山羊生长性状相关性的研究未见报道,因此,本研究以KAP6.2基因作为目标基因,探索KAP6.2基因的InDel多态性,并分析其与内蒙古绒山羊生长性状和绒细度性状的关联性,以期为内蒙古绒山羊的选种选配奠定基础。
1 材料与方法
1.1 材料
1.1.1 试验动物 试验所用内蒙古绒山羊均来自陕西省榆林市某养殖基地。随机选取409只(1.0~1.5岁)育成羊和189只(1.5~4.0岁)成年羊,均为雌性。试验羊均舍饲,饲喂环境和饲喂日粮相同。
1.1.2 样品的采集与生长性状的测定 剪取试验羊约2 g的耳组织样,放入含有1.0 mL 70%酒精的2.0 mL Eppedorff管中,冰盒中保存带回,置于-20 ℃冰箱保存。同时测量试验羊的10个生长性状数据,包括体重、体高、体长、胸围、髋骨宽、管围、十字部高、胸深、胸宽[21],测量体尺的同时,贴皮取试验羊肩胛骨处约3 g羊绒样品并测定绒细度。
1.2 方法
1.2.1 DNA提取与浓度测定 受试羊耳组织的基因组DNA参照高盐法[22]提取,利用Nanodrop分光光度计和10 g/L 琼脂糖凝胶电泳分别测定DNA浓度和纯度,稀释至20 ng/μL,-40 ℃保存备用。
1.2.2 引物设计与合成 参考NCBI数据库中公布的山羊KAP6.2基因序列(GenBank登录号:AY316158.1),利用Zhao等[23]文章中的引物对候选变异位点进行多态性检测,引物由上海生工生物工程技术服务有限公司合成。
1.2.3 PCR体系与反应条件 总PCR反应体系(25 μL):PCR Master Mix 12.5 μL,上下游引物各0.5 μL,DNA模板1 μL,以ddH2O补至25 μL。PCR扩增条件参照赵俊星等[24]的文章,扩增产物用3%的琼脂糖凝胶电泳与测序鉴定分型。
1.2.4 数据统计与分析 利用在线网站SHEsis(http://analysis.bio-x.cn)分析了KAP6.2基因InDel多态位点在内蒙古绒山羊群体中的Hardy-Weinberg平衡P值与基因型频率等结果,应用一般线性模型Yijk=μ+Gi+Eij分析不同参数对性状的影响,其中μ为总体平均值,Gi代表基因型固定效应,Eij代表随机误差。利用SPSS 23.0软件和单因素方差分析法分析KAP6.2基因InDel突变位点不同基因型与内蒙古绒山羊不同群体生长性状和绒细度性状的差异显著性,其中育成羊群体绒细度性状DD基因型只有1个,故运用独立样本t检验方法单独对II和ID基因型与育成羊绒细度性状进行关联分析。再取各个性状表型值的组内平均值,获取个体表型值与组内平均值的差值,以单因素方差分析法分析不同基因型与内蒙古绒山羊群体生长性状和绒细度性状之间的差异显著性,以上结果以“平均值±标准误”表示,P<0.05表示差异显著,P<0.01表示差异极显著。
2 结果与分析
2.1 KAP6.2基因的PCR扩增与测序
利用合成引物对KAP6.2基因外显子区24-bp InDel突变位点进行PCR扩增,结果显示(图1),KAP6.2基因InDel突变位点存在3种基因型:纯合插入型(II,362 bp)、杂合型(ID,362/338 bp)与纯合缺失型(DD,338 bp),测序结果(图2)与电泳结果一致。结果表明,KAP6.2基因24-bp InDel突变(AY316158.1.g:119-142 del CAGGATACGGCTGTGGATACGGTT)在内蒙古绒山羊群体中存在多态性。
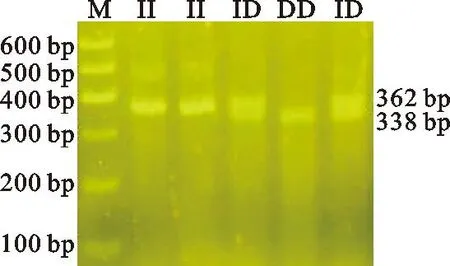
图1 KAP6.2基因InDel位点扩增Fig.1 Amplification of InDel site of KAP6.2 gene M. Marker 1;II:插入/插入;ID. 插入/缺失;DD. 缺失/缺失;下同 M. Marker 1; II. insertion/insertion; ID. insertion/deletion; DD. deletion/deletion; the same below
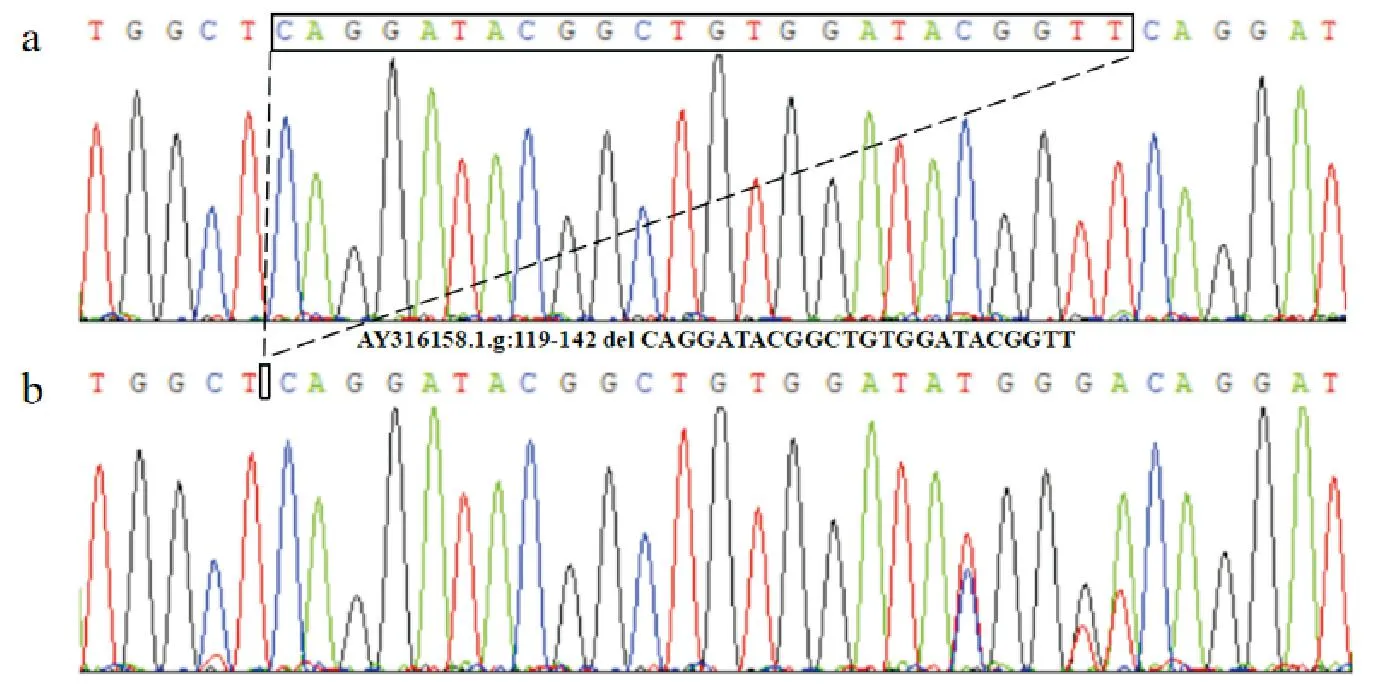
图2 KAP6.2基因InDel位点测序图a. ID基因型测序结果,b. II基因型测序结果Fig. 2 Sequencing of InDel site of KAP6.2 genea. ID genotype sequencing results; b. II genotype sequencing results
2.2 KAP6.2基因InDel位点遗传参数分析
根据基因分型结果对内蒙古绒山羊试验群体进行计算分析(表1),KAP6.2基因InDel突变位点在内蒙古绒山羊育成羊和成年羊群体杂合度分别为0.146和0.081,均属于低度多态(0.1
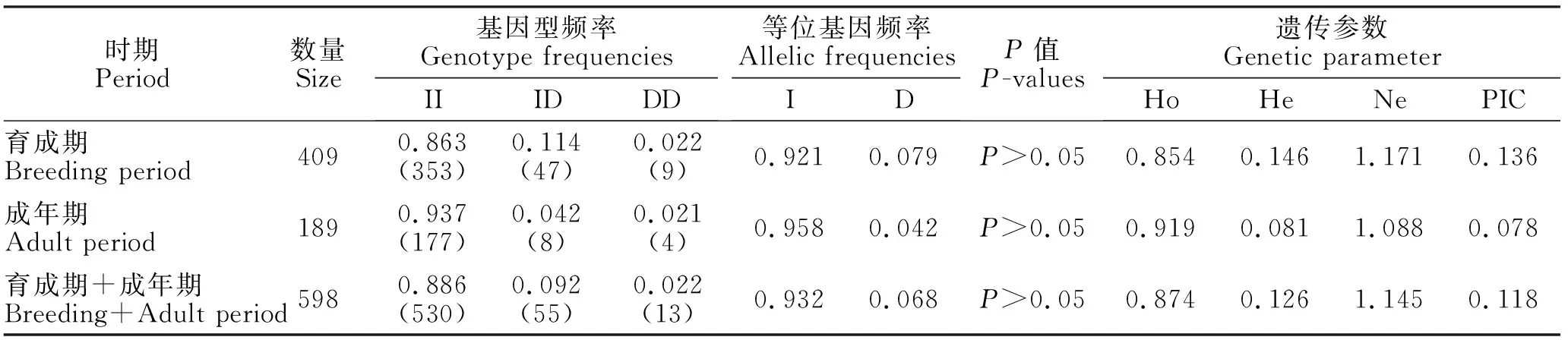
表1 内蒙古绒山羊群体KAP6.2基因多态位点遗传参数值Table 1 Genetic parameters values of KAP6.2 gene polymorphism locus in Inner Mongolia cashmere goat
2.3 内蒙古绒山羊KAP6.2基因多态性与生长性状和绒细度性状的相关分析
由表2可知,在育成羊中(n=409),该InDel突变与胸围、十字部显著相关(P<0.05),与体长、胸深和胸宽呈极显著相关(P<0.01);在成年羊中(n=189),该InDel突变与胸宽显著相关(P<0.05),与胸围、十字部高和胸深极显著相关(P<0.01),优势基因型均为DD;在整个内蒙古绒山羊群体中(n=598),该InDel突变与体长、胸围、十字部高、胸深和胸宽极显著相关(P<0.01),优势基因型为DD型;与绒细度显著相关(P<0.05),优势基因型为ID型。
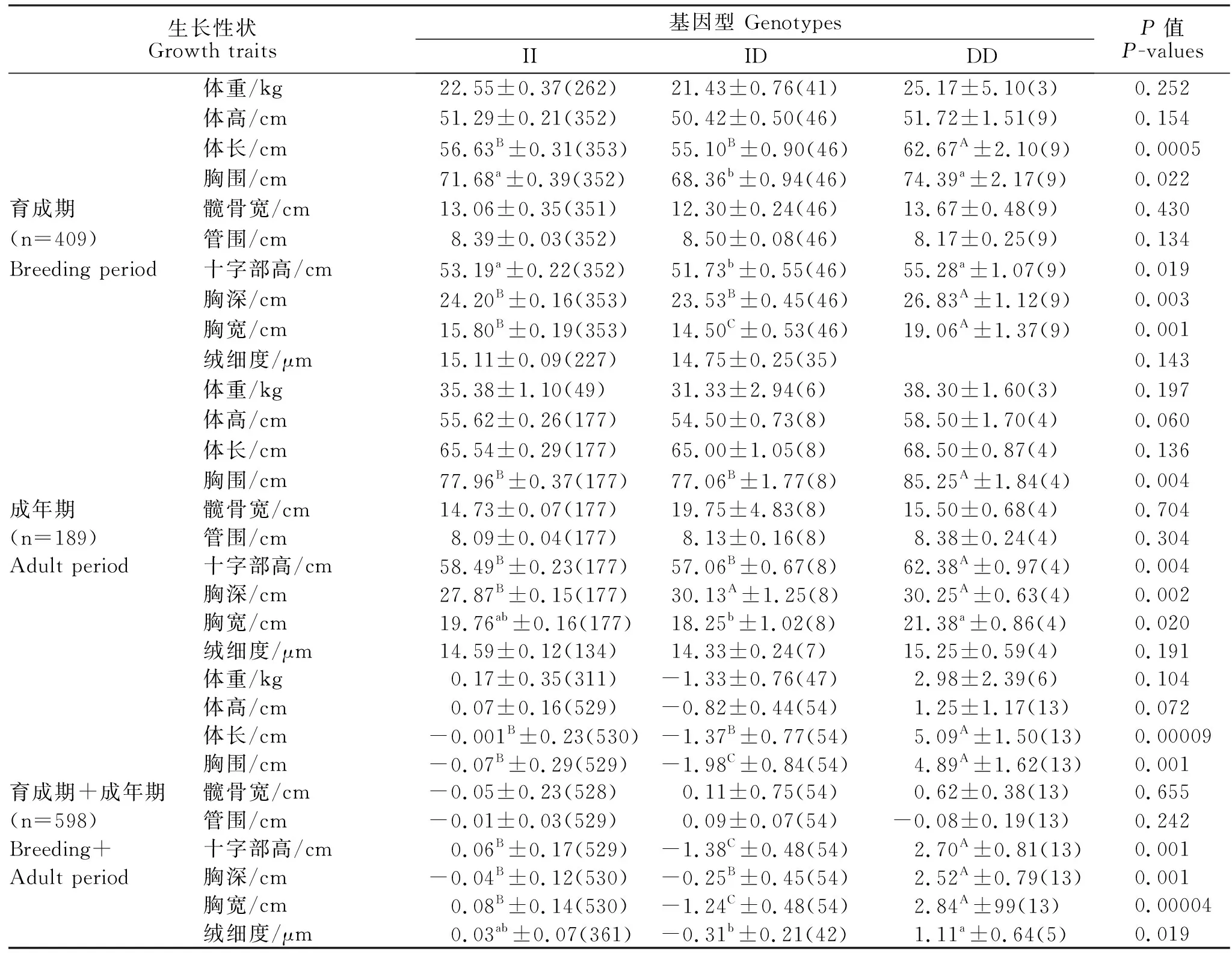
表2 内蒙古绒山羊育成羊KAP6.2基因多态性与生长性状和绒细度性状的关联分析Table 2 Correlation analysis of growth traits and cashmere fineness of Inner Mongolia cashmere goats in breeding period with polymorphisms in KAP6.2 gene
3 讨 论
内蒙古绒山羊养殖业是内蒙古地区主要的经济支柱,其羊绒细度较细,是多国纺织羊绒精品的主要原料[24-25]。但产肉量较低、绒细度变化较大是目前内蒙古绒山羊养殖过程中亟待解决的问题[26]。因此,利用选择现代分子标记辅助选择技术来提高经济性状和实现内蒙古绒山羊高效育种具有重要意义。
KAP6.2基因属于高甘氨酸酪氨酸蛋白多基因家族,目前在山羊上对KAP基因家族的研究主要集中在与毛、绒相关的多态性检测和选择候选基因上[27]。研究显示,KAP6.1基因可作为与优质细毛羊、辽宁绒山羊的产绒量和绒细度性状相关的遗传标记位点[17,28],同时该基因已定位到绵羊的1号染色体上[19]。多项研究表明,KAP6.2基因在多个山羊、绵羊品种中存在多态性,与部分品种绒、毛性状相关。例如,KAP6.2基因在辽宁绒山羊、辽-岢杂种山羊和黎城大青羊3个山羊品种内检测到18个SNP突变位点,并造成了氨基酸的序列变化[24],在绵羊和中卫山羊也检测到SNP位点,在中卫山羊为中度多态[29-30]。在高原型藏山羊上,KAP6.2基因与产绒量、绒长度和绒细度性状显著相关,可作为首选标记位点[31]。此外,KAP6.2基因在毛囊分化中具有空间表达变化的特征[32-33],在陕北白绒山羊存在24 bp的缺失突变,提示该缺失突变可能是陕北白绒山羊绒型多样、产绒量高的原因之一[23]。同时,KAP6.2基因的核苷酸和氨基酸序列在不同地域和不同品种山羊中高度保守,在不同生长发育阶段皮肤的初级、次级毛囊的皮质层和内根鞘中均有不同程度表达[34-35]。
本研究发现,KAP6.2基因外显子区24-bp InDel突变在内蒙古绒山羊群体中稳定存在且存在多态性,多态信息含量、杂合度等往往与群体的遗传变异程度呈正相关[36]。多态信息含量显示,在内蒙古绒山羊育成羊群体、成年羊群体和整个内蒙古绒山羊群体中,多态信息含量分别为0.136、0.078和0.018,均属于低度多态,可能是遗传分化程度较低,与其它种群基因交流较少导致。Hardy-Weinberg平衡检验表明,此突变位点均处于平衡状态,表明该群体未受到人工选育或者受人工选育影响程度较小,也可能是在遗传漂变、迁徙中处于动态平衡,可以对这些位点加强选择力度[37]。同时,此InDel突变位点与育成羊和成年羊胸围、十字部高、胸深和胸宽等多个生长性状具有显著或极显著关联,优势基因型均为DD型,该结果在具有显著或极显著关联的生长性状和优势基因型上具有很好的一致性,说明该位点可以作为内蒙古绒山羊选育的重要位点,在后续育种工作中优先选择DD型山羊个体进行选育。在整个内蒙古绒山羊群体中,KAP6.2基因此InDel突变与绒细度具有显著关联,而次级毛囊中KAP8.2表达量是初级毛囊的2.71倍,可能对绒细度具有重要调控作用[38],KAP6.2与KAP8.2同属高甘氨酸酪氨酸蛋白[8-11],推测KAP6.2基因可能通过类似调控从而影响绒毛纤维的表型性状,具体作用机制需进一步探究。绒细度与羊绒经济价值有着直接关系[39],在实际生产中意义重大,但在育成羊和成年羊群体中恰恰相反,没有表现出显著性,这可能与样本量大小有直接关系,但不排除与不同生长发育阶段和遗传差异的关系[34-35]。
4 结 论
KAP6.2基因外显子区24-bp InDel突变位点能够显著影响内蒙古绒山羊的生长发育,可作为内蒙古绒山羊部分生长性状和绒细度性状选育的候选基因,为内蒙古绒山羊选育工作提供理论依据。
猜你喜欢
今日农业(2022年14期)2022-09-15 01:43:28
昆钢科技(2022年2期)2022-07-08 06:36:18
系统工程学报(2021年4期)2021-12-21 06:21:08
新世纪智能(高一语文)(2020年12期)2020-06-01 08:14:20
建筑科技(2018年6期)2018-08-30 03:41:10
新农业(2016年23期)2016-08-16 03:38:12
新农业(2016年22期)2016-08-16 03:34:42
化纤与纺织技术(2015年1期)2015-12-26 03:33:40
中国医药导报(2015年27期)2015-02-28 22:08:01
当代畜禽养殖业(2014年7期)2014-02-27 07:59:13
